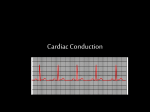
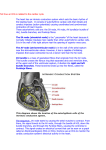
Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
Optical mapping of atrioventricular node reveals a conduction barrier between atrial and nodal cells BUM-RAK CHOI AND GUY SALAMA Department of Cell Biology and Physiology, School of Medicine, University of Pittsburgh, Pittsburgh, Pennsylvania 15261 rabbit atrioventricular node; compact node; atrioventricular nodal delay; decremental conduction; atrioventricular node conduction; photodiode array; voltage-sensitive dyes; 4-[b-[2-(di-n-butylamino)-6-naphthyl]vinyl]pyridinium (AV) node was first described by Tawara (25) in 1906 as the only electrical connection between the atria and the ventricles in mammalian hearts. The node is located between the interatrial septum (IAS) and interventricular septum (IVS) in a region called the triangle of Koch. It is composed of a spindle-shaped compact network of small cells that was found to be essentially the same in various species. The main function of the AV node is to delay depolarization (e.g., activation) between the atria and the ventricles and thereby coordinate their contraction. Histological analysis of the AV node indicated that the node consists of five morphologically distinct cell types: 1) transitional cells commingled with 2) atrial cells; 3) midnodal THE ATRIOVENTRICULAR cells, 4) lower nodal cells, and 5) cells of the penetrating AV bundle embedded within the central fibrous body (17). Transitional cells are distinguished from atrial cells by their smaller size, pale staining reaction, and extensive connective tissue. Midnodal cells are closely packed, have little intervening connective tissue, and form the ‘‘compact’’ node. Lower nodal cells are elongated, smaller than atrial cells, and form a bundle parallel to the AV ring. The AV node of the rabbit heart (,1.5 mm in length) has been extensively studied to characterize action potentials (APs) and activation delays in various regions of the AV node. AV nodal cells were divided into three zones on the basis of their electrophysiology (18): atrionodal (AN), nodal (N), and nodal-His (NH) cells. The N zone is an area of slow conduction and slow AP upstrokes, the AN zone is a transitional region between fast-conducting atrial muscle and the N zone, and the NH zone is a transitional zone between the N zone and the His bundle. This classification was not strict and was further extended on the basis of the AP response following a premature atrial stimulation (3). AN cells were further subdivided into AN and ANCO cells because, at fast pacing rates, APs of ANCO cells exhibited two components, or a notch, on the AP upstroke. Premature stimulation and pacing at faster rates also served to distinguish N from NH cells. To correlate a particular AP response to the morphology of the cell, APs were recorded with microelectrodes filled with potassium ferricyanide (24) or cobalt-containing KCl (1) to selectively stain cells that fired a particular type of AP. Such studies suggested that AN potentials emanate from transitional cells and NH potentials emanate from the lower nodal cells. It should be noted that the diffusion of the stain to neighboring cells made it difficult to demonstrate unequivocally that N-cell APs originate from anatomically defined midnodal cells (1). Activation delays across the AV node were measured with intracellular microelectrodes (3, 4); however, a detailed spread of activation within the midnodal and lower nodal zone could not be determined because markedly different AP characteristics and activation times could be measured at any location (but at unknown depths), with some cells activating early and others late in the same region. As a result, it was not possible to detect a wave of depolarization within the node and thereby measure a conduction velocity in the AV node. Hoffman and Cranefield (9) introduced the concept of decremental conduction to explain propagation delays at the AV node. AV node cells and, in particular, N cells have high intracellular resistance and reduced intercel- 0363-6135/98 $5.00 Copyright r 1998 the American Physiological Society H829 Downloaded from http://ajpheart.physiology.org/ by 10.220.32.247 on May 2, 2017 Choi, Bum-Rak, and Guy Salama. Optical mapping of atrioventricular node reveals a conduction barrier between atrial and nodal cells. Am. J. Physiol. 274 (Heart Circ. Physiol. 43): H829–H845, 1998.—The mechanisms responsible for atrioventricular (AV) delay remain unclear, in part due to the inability to map electrical activity by conventional microelectrode techniques. In this study, voltage-sensitive dyes and imaging techniques were refined to detect action potentials (APs) from the small cells comprising the AV node and to map activation from the ‘‘compact’’ node. Optical APs (124) were recorded from 5 3 5 mm (,0.5-mm depth) AV zones of perfused rabbit hearts stained with a voltagesensitive dye. Signals from the node exhibited a set of three spikes; the first and third (peaks I and III) were coincident with atrial (A) and ventricular (V) electrograms, respectively. The second spike (peak II) represented the firing of midnodal (N) and/or lower nodal (NH) cell APs as indicated by their small amplitude, propagation pattern, location determined from superimposition of activation maps and histological sections of the node region, dependence on depth of focus, and insensitivity to tetrodotoxin (TTX). AV delays consisted of t1 (49.5 6 6.59 ms, 300-ms cycle length), the interval between peaks I and II (perhaps AN to N cells), and t2 (57.57 6 5.15 ms), the interval between peaks II and III (N to V cells). The conductance time across the node was 10.33 6 3.21 ms, indicating an apparent conduction velocity (QN ) of 0.162 6 0.02 m/s (n 5 9) that was insensitive to TTX. In contrast, t1 correlated with changes in AV node delays (measured with surface electrodes) caused by changes in heart rate or perfusion with acetylcholine. The data provide the first maps of activation across the AV node and demonstrate that QN is faster than previously presumed. These findings are inconsistent with theories of decremental conduction and prove the existence of a conduction barrier between the atrium and the AV node that is an important determinant of AV node delay. H830 A CONDUCTION BARRIER REGULATES AV DELAY MATERIALS AND METHODS Experimental protocol. This study describes data from nine experimental groups of rabbit AV node preparations for a total of 44 hearts. In group 1 (n 5 3) hearts, the atrial-His bundle (AH) interval and AV delays were measured before and after the hearts were stained with the voltage-sensitive dye to examine possible pharmacological effects of the dye. In group 2 (n 5 4) hearts, APs were simultaneously recorded with microelectrodes and optical mapping techniques to help identify the origin of peak II (see Isochronal maps and movies of activation). In group 3 (n 5 9) hearts, activation across the IAS and the crista terminalis, conduction velocity across the compact node, and AV delay were measured. In group 4 (n 5 4) hearts, AP signals were analyzed as a function of depth of focus. In group 5 (n 5 4) hearts, optical recordings of APs were carried out, and then the tissue was labeled with fiducial marks to superimpose activation maps on histological sections of the tissue. In group 6 (n 5 3) hearts, AV node preparations were used to test the effects of tetrodotoxin (TTX). In group 7 (n 5 3) hearts, the effects of cutting the His bundle were investigated, and in group 8 (n 5 5) hearts, the effects of exposure to acetylcholine (ACh) were tested. In groups 1–8, the hearts were allowed to beat at their intrinsic rates, which were controlled by the sinoatrial (SA) node or primary pacemaker; a crush of the SA node interrupted the normal heart rate and, after a few minutes, the AV node became the primary pacemaker. In group 9 (n 5 9) hearts, the SA node was intentionally dissected and the heart was paced on the right atrium (1–2 mm below the SA node) to measure changes in AV delay and optical APs as a function of heart rate. Alternatively, the SA node was dissected to measure activation patterns with the AV node as the primary pacemaker. This investigation conformed with the Guide for the Care and Use of Laboratory Animals published by the National Institutes of Health [DHHS Publication No. (NIH) 85–23, Revised 1985, Office of Science and Health Reports, Bethesda, MD 20892]. AV node preparation. New Zealand White rabbits (2.0–3.0 kg) were anesthetized with pentobarbital sodium (Nembutal; 35 mg/kg) by intravenous injection in an ear vein. The chest was opened, and heparin (200 U) was injected in the inferior vena cava. After a few minutes, the heart was removed and perfused through the aorta in a modified Langendorff perfusion. The perfusate contained (in mM) 130 NaCl, 12.5 NaHCO3, 1.2 MgSO4, 4.75 KCl, 1.0 CaCl2, and 20 dextrose. Solutions were continuously gassed with 95% O2-5% CO2. The pH was adjusted to 7.4 with NaHCO3. Input of perfusate at the aorta was controlled with a peristaltic pump (Minipuls 2, Gilson, Middleton, WI) and was connected to a graduated manometer to obtain a physiological mean aortic pressure of 80 mmHg. Aortic pressure was monitored with a manometer and/or a pressure transducer (P10, Statham, Waltham, MA). The flow rate of the pump determined the flow of perfusate delivered to the coronary vessels and was adjusted to 7 ml · min21 · g wet wt21 at the beginning of each experiment and kept constant thereafter. A perfusion system with ‘‘constant coronary flow rate’’ instead of the more typical ‘‘constant aortic pressure’’ ensured that the flow of perfusate through the myocardium remained constant and homogeneous during changes of the contractile state and/or coronary resistance. The perfusate was not recycled through the heart, and only preparations with stable aortic pressures were selected for the study; initial pressures were in the range of 80–100 cmH2O, and final pressures at the end of the experiments changed by #5% of initial pressure. The free walls of the right ventricle and atrium were dissected open to expose the IAS and IVS and were pinned down on a Sylgard-coated horizontal chamber. The chamber was water-jacketed to control temperature, which was continuously monitored with a thermistor placed near the optical field of view. A heating coil in the chamber was used to continuously adjust the temperature of the bath via a feedback system. Surface electrograms were recorded with bipolar electrodes (Teflon-coated platinum wires, 250 µm and 1 mm apart) placed at key sites on the preparation: 1) near the SA node, 2) on the IAS near the AV ring, 3) close to the His bundle, and/or 4) on the IVS. Staining procedure. The heart was stained with the voltagesensitive dye 4-[b-[2-(di-n-butylamino)-6-naphthyl]vinyl]pyridinium (di-4-ANEPPS; Molecular Probes, Eugene, OR) by gradual injection of 200 µl of a stock solution of dye [2 mM in dimethyl sulfoxide (DMSO)] into the bubble trap over a period of 5–10 min. Other voltage-sensitive dyes, RH-421, di-8-ANEPPQ, and di-12-ANEPPQ (Molecular Probes, Eugene, OR), were also tested in attempts to obtain the highest possible signal-tonoise ratio for APs from the AV node. All four dyes were tested by preparing stock solutions in DMSO or 1:1 mixtures of DMSO and Pluronic acid. Each stock solution of dye was tested by either injecting the dye in the coronary perfusate or adding dye to the bathing solution (1–2 mM) for 30–45 min. Downloaded from http://ajpheart.physiology.org/ by 10.220.32.247 on May 2, 2017 lular coupling compared with atrial and ventricular cells. The high coupling resistance could explain the basic conduction delay of the AV node. Such decremental conduction would also predict a gradual delay across the N zone so that delay is distributed across the cell network. However, Billette (3) demonstrated that the conduction delay is not decremental in space following a premature stimulus but seems more localized in N cells, where conduction stagnates. Studies from several investigators (1, 3, 4, 8) have led to the realization that slow conduction in the AV node cannot be solely explained by active properties of N cells, such as the maximum rate of rise of the AP upstroke (dV/dtmax ), and that both passive and active properties are responsible for the inhomogeneous potential spread in the N zone. In the present report, we applied voltage-sensitive dyes and optical imaging techniques to map electrical activity across the AV node in attempts to elucidate the mechanisms responsible for the AV node delay. Voltagesensitive dyes have been extensively used by various investigators (7, 20, 22, 23) to measure optical APs in a variety of cardiac muscle preparations. Simultaneous recordings of transmembrane potential by optical and microelectrode techniques have validated the high fidelity of optical APs compared with microelectrode recordings and demonstrated that optical APs detected the classic features of atrial, pacemaker, and ventricular APs (14, 23). Optical techniques also face important limitations because the absolute value of membrane resting potential cannot be obtained unless calibrated with a microelectrode, the downstroke of the AP can be distorted by movement artifacts, and the optical AP represents the sum of APs from cells within a region of tissue and not the AP of a single cell. Despite these limitations, the technique offers important advantages in mapping the inputs to the node, detecting the activation sequence in the compact node, and identifying zone(s) of conduction delay. A CONDUCTION BARRIER REGULATES AV DELAY Fig. 1. Bipolar surface electrodes were used to measure atrial-His (AH) intervals before (A), during, and after (B) perfusion with 4-[b-[2-di-n-butylamino)-6-naphthyl]vinyl]pyridinium (di-4-ANEPPS). One bipolar electrode was placed on the interatrial septum (IAS) halfway between the sinoatrial (SA) and atrioventricular (AV) nodes to detect atrial depolarization [atrial bipolar electrogram (BE)]. A 2nd electrode was placed at junction of tendon of Todaro (TT) and crista terminalis (CT) to detect His bundle and ventricular depolarizations (His BE). from two 100-W tungsten-halogen lamps was collimated, passed through a 520 6 20-nm interference filter, and focused on the triangle of Koch. Fluorescence emission from the stained tissue was collected with a camera lens (50 mm, f1:1.4, Nikon), projected through a 630-nm cut-off filter (RG-645, Schott Glass), and focused to form an image of the preparation on the surface of a 12 3 12 photodiode array (Centronic, Newbury Park, CA). Five diodes from each corner of the array were ignored such that optical signals were monitored from 124 of 144 diodes. The image of the AV node was focused on the array at a magnification of 30.36 such that each diode detected fluorescence APs from a 0.46 3 0.46 mm area of epicardium. The depth of field of the collecting lens restricted the fluorescence measurements to a layer of cells ,100 µm from the surface. The depth of field of AP recordings was estimated by varying the staining protocol and by empirical calculations based on the magnification of the image, the numerical aperture of the lens, and the wavelength of the emitted light (19). The image of the heart focused on the array was reflected by a mirror onto a custom-made graticule with the exact dimensions of the array (Graticules, Tonbridge, UK) located on a plane parafocal with the plane of the array. Precise focusing and aligning of the heart with respect to the array was accomplished by focusing and aligning the image of the heart on the graticule. The photocurrents from 124 diodes were fed to a current-tovoltage converter, amplified, digitized (1.56 kHz per channel, 12-bit resolution per sample), and stored in a memory buffer of an IBM PC 486DX/4 100-MHz computer. The sampling rate (1.56 kHz per channel) was set to the maximum rate of the data acquisition processor (DAP 1200e, Microstar Laboratories, Bellevue, WA). A data acquisition scan consisted of 128 simultaneously recorded traces: 124 optical plus 4 instrumentation channels. A scan consisted of a continuous recording of these 128 channels for 1.2–3 s. Simultaneous AP recordings with microelectrodes and voltage-sensitive dyes. In some experiments, conventional 3 M KCl-filled microelectrodes with a resistance of 20–40 MV were used to record APs from the AV node to identify the APs that fired in synchrony with the second spike of the optical recordings. Optical APs were recorded from the AV node zone, and then the diodes on the array that detected three spikes were used to identify the mid- and lower nodal zone, which was then impaled with a microelectrode to simultaneously record APs by the two techniques. These measurements were challenging because the short working distance between the lens and the preparation made it difficult to obtain stable microelectrode impalements at a shallow angle of penetration. Data analysis. Several criteria were used to assess the ‘‘health’’ of AV node preparations: 1) the rapid rise time of atrial and ventricular AP upstrokes, 2) the short AH intervals [,80 ms at 500 ms cycle length (28)], 3) the rapid propagation of APs in the atrium (,0.1 m/s), and 4) the duration of ventricular APs, because short AP durations are indicative of ischemic injury. The activation time point at each diode was taken as the time point of the maximum rate of rise of the fluorescence (F) AP upstroke (dF/dtmax ), which represents the time when most of the cells depolarized (21, 22). Optical recordings from each channel were normalized and passed through a Butterworth filter, and the first derivative of each fluorescence signal (dF/dt) was calculated by a numerical differentiation method using a three-point Lagrangian interpolation. The dF/dtmax time point was accepted as an activation time if dF/dtmax was greater than the standard deviation (SD) of background noise. The analysis of optical APs was automated using in-house software written with IDL 3.6.1b Downloaded from http://ajpheart.physiology.org/ by 10.220.32.247 on May 2, 2017 For all four dyes, staining the hearts by injections in the coronary circulation was markedly more effective than bathing the preparation in dye solution such that greater amplitudes of optical AP upstrokes, signal-to-noise ratios, and more homogeneous staining were obtained. Di-4-ANEPPS gave the best signal-to-noise ratio and was used for the experiments described in this study. In one experimental group (n 5 3), AH intervals were measured with bipolar surface electrograms before, during, and after perfusion of the hearts with dye solution to examine the possible pharmacological effects of DMSO with or without dye on AV conduction delays. Figure 1 shows the AH measurements from an AV node preparation. AH intervals averaged over a period of 5 min before hearts were stained were 42.7 6 2.4 ms (Fig. 1A); after hearts were stained, AH intervals were 43.3 6 2.5 ms when averaged over 5 min of dye washout (Fig. 1B; n 5 3 hearts). Thus the present staining conditions with the use of a DMSO stock solution of di-4-ANEPPS caused no detectable changes in AV node conduction. To avoid motion artifact to optical recordings, 10 mM diacetyl monoxime (DAM) solution was used to block contraction during data acquisition. Hearts were perfused in Tyrode solution containing DAM for 10–12 min and then perfused with standard Tyrode to avoid prolonged exposure to DAM, which produced time-dependent changes in AP characteristics. Exposure to DAM (10 mM) did not appear to alter AV conduction during brief periods of DAM perfusion as used in these experiments, but prolonged perfusion (.45 min) produced a gradual increase in coronary resistance in total AV delays followed by inexcitable atrial and ventricular tissue. Optical apparatus. Details of the optical and recording apparatus have been described elsewhere (22). The horizontal chamber was mounted on an X-Y-Z micromanipulator to accurately select the zone of tissue viewed by the photodiode array and to control the location of the focal plane of the optical apparatus as a function of depth in the tissue. Light H831 H832 A CONDUCTION BARRIER REGULATES AV DELAY by the operator using an interactive program. The detection of activation time points using dF/dtmax was highly reliable and rarely required correction by the investigator (,1% of activation events). The upstrokes of optical APs from the AV node were distorted by movement artifacts from vigorously contracting hearts because atrial contractions interfered with the small signals from the AV node zone such that the use of DAM (as described in Staining procedure) was essential to abate movement artifacts. Histology. Optical electrophysiological recordings were made to identify the zone of tissue that fired three sequential spikes, and then the tissue was marked with fiducial points at sites that corresponded to the edges of the array. The fiducial marks were made by impaling the tissue with a microelectrode and then placing a silk surgical thread suture at those sites to ensure that the fiducial mark was clearly identifiable even after the histological processing. The tissue containing the AV node region was excised and fixed in Bouin’s solution overnight. The fixed tissue was embedded in paraffin, and serial sections 5 µm thick were taken starting from the endocardial surface, and every tenth section was mounted on a glass slide. Mounted sections were progressively stained with Mayer’s hematoxylin, counterstained with eosin, and placed under a glass coverslip (15). Stained sections were examined under a microscope and captured in TIFF images (in 24-bit true color) using a charge-coupled device camera. Shrinkage caused by fixation (,35%) was automatically taken into account by aligning the fiducial marks on the sections with their respective locations on the edges of the array. The isochronal maps derived from optical APs were superimposed on images of the tissue derived from histological sections using CorelDraw 7.0. RESULTS Optical recordings from AV node. Figure 2A shows an anatomic sketch of the AV node region delineated by the tendon of Todaro and the crista terminalis forming the triangle of Koch and the central fibrous body. A symbolic map of the array is superimposed on the AV node to identify the region of tissue mapped by the array. The orientation of the array relative to the AV node was arbitrary but was kept constant in the present study. Figure 2B shows a set of 124 optical signals recorded from the AV node region as well as simultaneously recorded atrial and ventricular bipolar electrogram (BE) recordings. Figure 3, A–D, shows the four types of signals recorded from the AV node region and the temporal relationship between these signals and the surface electrograms. Each panel of Fig. 3 represents a more detailed tracing of recordings from diodes a–d in Fig. 2A. Each panel contains three traces, the atrial and ventricular BEs plus an optical recording from one of the diodes (a–d) viewing different sites on the preparation. Diode a (see Fig. 2A) viewed the IAS and detected atrial APs with negligible contribution from other cell types such as nodal or ventricular cells (Fig. 3A). Diode b (see Fig. 2A) viewed the IVS and detected ventricular APs (Fig. 3B). At border zones between the IAS and IVS, diode c (see Fig. 2A) detected voltagedependent optical responses with two upstrokes per cardiac beat (Fig. 3C). In these zones, the IAS overlapped ventricular tissue such that both cell types contributed to the signal recorded by the same diode, resulting in the sequential firing of atrial, and then Downloaded from http://ajpheart.physiology.org/ by 10.220.32.247 on May 2, 2017 (Interactive Data Language, Research Systems, Boulder, CO) and Borland C11 4.0. Isochronal maps and movies of activation. Optical signals from atrial and ventricular muscles exhibited the expected shapes and characteristics of atrial and ventricular APs. In contrast, signals from the AV node region (the region delineated by the tendon of Todaro and the crista terminalis) exhibited a set of three sequential spikes indicative of three distinct depolarization events separated by marked time delays (see Figs. 2–4). The first and third spikes (peaks I and III) were coincident with atrial and ventricular depolarizations, respectively. The second spike (peak II) was only detected in the AV node region and was the smallest in amplitude. Thus three cell types were found to overlap in the same zone of AV node tissue, and activation patterns were independently generated for the three spikes. The combined activation patterns represented atrial, nodal, and ventricular activation that corresponded anatomically to the atrial, nodal, and ventricular regions of the preparation. Isochronal maps were generated for the three wave fronts using a linear triangulation method (Tecplot-3DV, Amtec Engineering, Bellevue, WA). To map activation in the AV node zone, the region that exhibited peak II was extracted first and was typically delineated by 12–18 photodiodes, or areas of 4 3 3 or 6 3 3 diodes, on the array. Activation time points for all peak II were triangulated, isochronal lines were calculated for every 1-ms interval using the values of the three edges of the triangles by linear interpolation, and then points were connected by lines. With linear triangulation, 10–12 isochronal lines were typically generated in a zone delineating 2–3 mm of nodal tissue. Conduction velocity was determined from the time delays between isochronal lines and the distance traveled by the wave front detected in the two-dimensional (2-D) field of view of the array. These measurements represent ‘‘apparent’’ conduction velocities given that the precise pathways of the wave fronts are approximate, because the node is a three-dimensional (3-D) structure (0.75–1.25 mm thick according to histological analysis) and the pathway is determined from 2-D maps. As a result, conduction velocities may be underestimated if the pathway propagation spreads in depth across the thickness of the node as well as in the 2-D field of view. The spread of depolarization was animated to visualize the spread of excitation waves. The animation program was written with g11 2.7.2 and YORICK 1.2 by scaling the amplitudes of the optical recordings into a range of 8-bit, 256-color levels. The signal-to-noise ratio was sufficiently high so that filtering of the normalized signals was not necessary. One frame of the byte-scaled data from the 12 3 12 array was converted to a 120 3 120 pixel image that consisted of 12 3 12 squares (the level of depolarization was color coded and filled an array of 10 3 10 pixels). The sequence of excitation images was displayed on the monitor using X Windows System (X Consortium) ‘‘pixmaps.’’ The typical animation runs at 20 frames/s. Mapping and animation were performed on a Pentium (100 MHz) running Linux (kernel version 2.0) or an Indy workstation (Silicon Graphics, Mountain View, CA) running Irix 5.3. Movies of AP propagation across the AV node can be seen at our web site (Salama, Guy. Lab and current research: AV node conduction. [On-line] Dept. of Cell Biology and Physiology, Univ. of Pittsburgh. http://www.cbp.pitt.edu/la gs.htm); QuickTime movie player is required. The activation time point for each fluorescence AP was taken at dF/dtmax, when most of the cells viewed by a diode are depolarizing. Activation time points for all 124 APs were labeled with ‘‘tick marks’’ that could be verified and corrected A CONDUCTION BARRIER REGULATES AV DELAY ventricular, APs. The interpretation of the cell types responsible for these optical signals was validated by the electrogram recordings in that the upstrokes of atrial and ventricular optical APs were coincident with the atrial and ventricular electrograms, respectively (Fig. 3, A–C). In addition, atrial and ventricular APs were recorded optically at sites consistent with the anatomy of the preparation: they had the expected shape and time course of atrial and ventricular APs, and the delay between the firing of the two upstrokes was consistent with the expected AV delay. More interesting was a unique feature of optical signals from the AV node region shown in Fig. 3D. Signals recorded from a narrow zone between the tendon of Todaro and the crista terminalis (diode d; see Fig. 2A) exhibited a set of three sequential depolarizations per cardiac beat, peaks I, II, and III (Fig. 3D). By superimposing signals detected by other diodes and surface electrodes, peak I was again coincident with atrial depolarization and peak III was coincident with ventricular depolarization. Peak II consistently fired at intermediate time points and was only observed in a narrow (1 3 2 mm) region of the preparation. AP propagation within AV node. To elucidate the origins of peak II, intracellular microelectrodes and optical APs were simultaneously recorded from the zone that exhibited the sequence of three depolarizations. As shown in Fig. 4A, the firing of peak II was coincident with APs with the characteristic shape of N and/or NH cells because of the slow diastolic depolarization and slow upstroke (n 5 4 hearts). In contrast, microelectrode recordings of atrial, AN, His, and ventricular APs were not coincident with peak II depolarization (not shown). More detailed analysis of the spatiotemporal characteristics of peak II was carried out to reinforce the interpretation that peak II originated from the midnode and/or lower node region and to map electrical conduction across the AV node. Figure 4B shows the approach used to calculate the time delays t1 and t2 from the time points of dF/dtmax for peaks I–III. Maps of electrical activation generated from activation time points (as in Fig. 4A) were highly reproducible from beat to beat of the same heart and from heart to heart. Figure 5 shows an experiment from a spontaneously beating heart with a cycle length of 417.92 6 0.96 ms (mean 6 SD). The mean AV delay (6SD) measured with surface electrograms was 88.75 6 0.37 (n 5 5 beats). As shown in Fig. 5, the time points of dF/dtmax for peaks I–III were detected separately and used to map the spread of activation in the IAS (Fig. 5A), the AV node (e.g., the zone of tissue detecting peak II) (Fig. 5B), and the IVS (Fig. 5C). Figure 5D depicts the superimposition of all three activation patterns, showing that the three maps overlap in a small region identified as the compact node. Activation across the IAS occurred in 15 ms (Fig. 5A), and, after a substantial delay, the AV node signal appeared at 43 ms (Fig. 5B) and spread across the node in 8 ms. After another delay (e.g., the time to propagate from the His bundle to the apex and back to the base of the ventricle), the IVS fired APs at 93 ms and spread in the field of view in 4 ms (Fig. 5C). Such maps of conduction across the AV node were highly reproducible in both their patterns and temporal relationships. Table 1 lists the composite analysis of nine rabbit preparations. The means 6 SD for t1 and t2 were calculated for rabbit AV nodes (n 5 9) under sinus rhythm, with an intrinsic cycle length of 301.64 6 8.41 ms. The time for peak II to propagate across the compact node (i.e., midnode) was 10.33 6 3.21 ms, which predicts a conduction velocity of 0.162 6 0.024 m/s. The conduction velocity measured by optical techniques is faster than that inferred from intracellular microelectrode recordings because the latter measurements were not based on multiple recordings within the node and included a substantial component of t1 or AN delays. In Fig. 6, an activation pattern across the AV node is shown as a sequence of activation maps captured at different time points during a single cardiac beat. Each map is a pseudocolor map of the array for which the extent of depolarization at each site is color coded from violet to red (from least to most depolarized potential). The first map (at time t 5 0.0 ms) is violet or blue (i.e., the tissue is at resting potential). The next map, at 3.2 ms, shows the initial firing of the atrium that spreads along the IAS in the subsequent maps (from 3.2 to 25.6 ms). After a partial repolarization of the IAS (from 25.6 to 41.6 ms), the firing of nodal APs begins in the lower left zone of the array (44.8 ms) and spreads along a Downloaded from http://ajpheart.physiology.org/ by 10.220.32.247 on May 2, 2017 Fig. 2. A: sketch of AV node preparation superimposed on a symbolic map of photodiode array to delineate AV node region being viewed. Anatomic landmarks of AV node region are identified to correlate optical signals obtained from each site with origins of signals. Compact node, or midnodal region, is delineated by a speckled outline and is bounded by TT and CT, surrounded by IAS above and interventricular septum (IVS) below, and adjacent to central fibrous body (CFB). Orientation of array relative to AV node zone was arbitrary but was kept the same for all experiments shown. B: simultaneously recorded optical action potentials (APs) from 124 sites on AV node zone. A symbolic map of array is shown as 124 square boxes, each identifying the location of individual diodes. The region of tissue viewed by each diode corresponds to map in A. The optical trace recorded by each diode is shown in its respective location in a compressed time base (400 ms). Inset: BE recordings, 1 located on IAS 2 mm above TT (A; outside field of view of array; atrial BE) and 1 on IVS near apex of ventricle (ventricular BE), were simultaneously recorded along with 124 optical signals. Optical APs recorded by diodes at locations a–d (A) are shown in Fig. 3. H833 H834 A CONDUCTION BARRIER REGULATES AV DELAY narrow zone (1–2 diodes wide), at first slowly and then faster, into the His bundle (from 44.8 to 54.4). After a delay (from 60.8 to 80.0 ms), the IVS depolarizes and ventricular signals overlap the AV node zone (from 80.0 to 86.4 ms). Video animation of these maps is more effective for visualization of the propagation pathway and can be accessed from our web page (see Isochronal maps and movies of activation). Depth resolution and amplitude of AV node APs. The AV node is a 3-D structure located near the surface of the IAS, with the majority of cells comprising the compact node found ,0.06–0.5 mm below the surface of the IAS (6). Microelectrode studies have identified zones of AN that overlap zones of N and NH cells on the basis of their AP characteristics (6). The 3-D nature of this structure implies that the focal plane of the imaging system should be located below the surface of the preparation to obtain maximum signal amplitudes for AV nodal APs. As shown in Fig. 7, a given diode will detect light from different volumetric zones of tissue as the focal plane of the optical apparatus is shifted below the surface. This is a natural consequence of optical detection because light from cells located on the plane of focus is transferred to the diode with the highest efficiency, whereas light from cells above and below the focal plane is transferred to the diode with decreasing efficiency. The efficiency of light transfer as a function of depth (at any given wavelength of light) depends on the ‘‘energy transfer function’’ of the collecting lens, measured at a particular optical magnification (21). The AV node was optically sectioned by recording APs from the surface and then from deeper optical sections by shifting the plane of focus into the tissue in 250-µm steps. Changes in the relative amplitude from peak I to peak II were used to estimate the depths of the cells responsible for the generation of peak II. As shown in Table 2, the relative amplitude from peak II to peak I increased with increasing depth of the focal plane for 0, 0.25, and 0.5 mm and then decreased for 0.75 and 1.0 mm. The most likely explanation for these depth-dependent changes is that peak I emanated from the surface and peak II emanated from cells ,0.5 mm below the surface. In all experiments, the focal plane was first adjusted to maximize peak II relative to peak I at the beginning of each experiment. This approach made it possible to maximize the signalto-noise ratio for peak II, delineate the 2-D distribution of the cells that fire peak II, and map the spread of activation of peak II. Activation sequence of pacing AV node. A key feature of the AV node is that, in the absence of SA node activity, N or NH cells can become primary pacemak- Downloaded from http://ajpheart.physiology.org/ by 10.220.32.247 on May 2, 2017 Fig. 3. Classification of optical signals from AV node region. Optical APs from AV node region shown in Fig. 2B exhibited 1 of 4 possible characteristics depending on location of recording. Each class of optical AP was temporally correlated with atrial and ventricular BEs to show firing sequence of APs. A–D each show a set of 3 traces, the atrial (top) and ventricular (bottom) BEs plus an optical recording (middle), from diodes labeled a–d, respectively, in Fig. 2A. A: optical AP recorded from atrial cells located on IAS by diode a. B: ventricular AP recorded from IVS by diode b. C: optical APs from AV boundary were recorded by diode c. At such sites, atrial and ventricular cells overlapped, resulting in optical signals with 2 spikes, synchronous with atrial and ventricular BEs, respectively. D: optical recordings from AV node at diode d invariably consisted of 3 sequential depolarizations (peaks I–III). Peaks I and III were coincident with atrial and ventricular BEs, and peak II fired at an intermediate time point, indicating that AV node was origin of signals that exhibited 3 sequential spikes. DF/F, fractional fluorescence change. A CONDUCTION BARRIER REGULATES AV DELAY ers. To investigate the spread of activation in hearts, we removed the SA node so that the AV node became the primary pacemaker (n 5 4). Figure 8A shows the optical signals detected from the AV node zone when pacemaker activity was initiated at the AV node. In this case, the initiation of the heart beat begins in the node [AV node (midnodal)] at a time delineated by the time line labeled a (Fig. 8A). As a primary pacemaker, the AV node trace (or peak II) fired first, and the atrial and ventricular BE are delayed (,28 ms) and are coincident with the time line labeled c (Fig. 8A). Note that, as expected, the optical recording of ventricular AP was coincident with the ventricular BE. On the other hand, the recovery of atrial AP was coincident with the time line labeled b (Fig. 8A) and precedes the atrial BE because the optical signal recorded APs near the AV node at an early time of activation, whereas the BE was located on the IAS, near the SA node. When the AV node is the dominant pacemaker, the amplitude and slope of the first depolarization was markedly decreased as expected because peak II is now the leading wave of depolarization originating from the AV node. In Fig. 8B, activation maps indicate that impulses were initiated in a narrow zone within the triangle of Koch, and the first site to activate fired in the node as indicated by the asterisk. The signal spread in 10 ms within the node, and after a substantial delay (27.96 ms; see arrow in Fig. 8B), activation spread across the IAS. The zone of activation delineated by peak II during sinus rhythm (not shown) was the same as that delineated by the leading wave of AP upstrokes when the AV node became the pacemaker (Fig. 8B). The marked conduction delay from the AV node zone to the IAS (Fig. 8B) indicates that propagation is not smooth but is discontinuous in both the anterograde and retrograde directions. This barrier to conduction (arrow) between the compact node and the IAS is dramatically evident in all the activation maps (Fig. 8B). The activation pattern within the ventricular tissue was intentionally omitted from the activation maps of Fig. 8B so that AV nodal activation would not be obscured. Superimposition of activation maps and histological sections of AV node. In four hearts, optical maps of APs were recorded from the AV node region and the tissue was labeled with fiducial marks, fixed, embedded in paraffin, serially sectioned, and mounted on microscope slides for histological analysis. Figure 9A shows the anatomic landmarks of the AV node seen from longitudinal cross sections taken from the endothelial surface at increasing depths in the tissue. The conventional anatomic landmarks of the AV node are all resolved at a depth of 100 µm (Fig. 9A, left), namely, the IAS, the pale cells of the compact node, the IVS, and the His bundle. As shown in Fig. 9, A and C, zones of fat cells are found adjacent to the AV node region. These zones of adipose cells are typical of mammalian AV nodes and extend continuously at all depths of the preparation. In this heart, the dimensions of the AV node zone were largest at a depth of 150 µm (Fig. 9A, middle) and decreased at a depth of 200 µm (1-mm scale at right applies to A–C). Activation maps recorded from the same preparation are shown in Fig. 9B as a set of maps taken every 3.2 ms. In this case, the SA node was removed and the AV node became the primary pacemaker. The superimposition of isochronal maps derived from optical recordings and the histological section taken at a 150-µm depth showed that the compact node is the tissue underlying the optical map-identified activation patterns across the AV node. Moreover, the earliest sites to fire APs in Downloaded from http://ajpheart.physiology.org/ by 10.220.32.247 on May 2, 2017 Fig. 4. Detection of activation time points in AV node. A: simultaneous optical and microelectrode recordings from AV node. Optical signals detected a sequence of 3 depolarizations, and peak II was coincident with an AP recorded from a microelectrode impaled in a midnodal or distal node cell, either an N or NH cell. The shape and time course of the microelectrode AP show that peak II fired in synchrony with an N and/or an NH cell. B: 1st derivatives of all 4 classes of signals (dF/dt) were taken to analyze spread of activation across AV node and to determine time point of activation at each site. For signals from AV node, dF/dt identified 3 separate activation time points. Top: DF/F recorded from a diode viewing AV node exhibited a set of 3 upstrokes (arrows) per cardiac beat (peaks I–III). Bottom: delays between peaks I and II (t1 ) and between peaks II and III (t2 ) were determined from time points of maximum dF/dt (dF/dtmax ) of voltagesensitive fluorescent signals. dF/dt of peak II allowed identification of activation time points for cells comprising AV node that were used to map activation patterns across node. H835 H836 A CONDUCTION BARRIER REGULATES AV DELAY these maps were located in the proximal region of the compact node (i.e., posterior), and subsequent APs fired in the distal zone of the AV node (i.e., anterior), followed by the His region, with increasingly faster propagation velocity. The superimposition of optical isochrones and histological images (Fig. 9C) provides compelling evidence that these optical signals originate from the firing of APs in the AV node and delineate the shape and boundaries of the AV node. Figure 10 shows another example of activation isochrones across the AV node toward the His bundle and depicts the marked increase in conduction velocity from the proximal to the distal portion of the node. The traces (Fig. 10A) show the Table 1. Analysis of voltage-dependent fluorescent signals from atrioventricular node Parameter t1 , ms t2 , ms Conduction time of peak II, ms QN , m/s 49.50 6 6.59 57.57 6 5.15 10.33 6 3.21 0.162 6 0.024 Values are means 6 SD for n 5 5 optical signals recorded from 9 rabbit atrioventricular (AV) node preparations. Delays between peaks I and II (t1 ) and between peaks II and III (t2 ), conduction time of peak II, and AV node conduction velocity (QN ) were calculated and grouped for rabbit hearts under normal sinoatrial node activation, with a cycle length of 301.64 6 8.41 ms. QN was determined from time delay and distance measured from isochronal maps of midnodal region of AV node as shown in Fig. 5 and did not include lower nodal or His bundle region shown in other experiments (see Figs. 9 and 10). QN , conduction velocity, and total time to propagate within node were highly reproducible from heart to heart. kinetics of optical signals recorded from these two regions. The top trace (Fig. 10A), recorded from the proximal region of the AV node trace, exhibited the typical sequence of three depolarizations. Here, peak II originated from the midnodal region, which initiated waves of depolarization (Fig. 10A, top arrow). The bottom trace, recorded from a more distal region, exhibited a sequence of two depolarizations, a His followed by a ventricular depolarization. Effect of TTX on AV node signals and conduction. The major depolarizing current of N and NH cells is a slow inward current (most likely Ca21 current) that is insensitive to the voltage-gated Na1-channel blocker TTX (12). In contrast, atrial, AN, His, and ventricular cells are somewhat sensitive to TTX, which, at 5–10 µM, decreases dV/dtmax. We investigated the effect of TTX (10 µM) on the propagation times across the AV node, that is, of peak II and His bundle depolarization, in the same manner as in Fig. 10A (n 5 3). Figure 10B shows an example of activation maps measured from the node before (left) and after (right) 20 min of TTX perfusion. TTX reduced the rise time of atrial and ventricular APs (not shown) but did not alter the spatiotemporal characteristics of peak II (compare the proximal zone of the AV node in Fig. 10, A and B). On the other hand, TTX produced a marked decrease in apparent conduction velocity in the distal zone of the activation map (Fig. 10B, right, TTX-sensitive zone). The sensitivity of the distal zone to TTX supports its identification as a His bundle region from the histological analysis (Fig. 9) and the spatiotemporal distribu- Downloaded from http://ajpheart.physiology.org/ by 10.220.32.247 on May 2, 2017 Fig. 5. Isochronal maps of activation across AV node. From activation time points obtained as described in Fig. 4, activation sequences of atrial (peak I), nodal (peak II), and ventricular (peak III) regions were triangulated individually and isochronal lines of activation were drawn 1 ms apart. Gray scale from bright to dark represents ‘‘early’’ to ‘‘late’’ activation times. Axes are length in millimeters. A: depolarization sequence of IAS generated from peak I. B: activation sequence of the AV node generated from peak II. C: activation sequence of IVS generated from peak III. D: composite isochronal map of AV node region generated from superimposition of activation maps in A–C reveals details about supraventricular activation and spread of activation across AV node and IVS. Note that from last depolarization of peak I to first depolarization detected through peak II, there is a 30- to 33-ms interval during which firing of APs is not detected anywhere in the preparation. A CONDUCTION BARRIER REGULATES AV DELAY H837 Downloaded from http://ajpheart.physiology.org/ by 10.220.32.247 on May 2, 2017 Fig. 6. Pseudocolor maps of activation across AV node region. A sequence of array images recorded at different time points during a single cardiac beat was generated as an alternative approach to display activation maps. Levels of local depolarization were linearly converted to rainbow color map (color scale 0–255), where least to most depolarized levels produced a color shift from blue to red, respectively. Successive images taken every 3.2 ms were generated from optical recordings as described in MATERIALS AND METHODS. Inset: orientation of images. tion of the signals (Fig. 10A). Thus the posterior zone consists of TTX-insensitive cells (most likely N and/or NH cells), and the anterior zone contains TTX-sensitive cells (most likely His and perhaps NH cells), in line with intracellular microelectrode recordings. Effect of His bundle cut. Figure 11A shows the atrial and ventricular BEs and optical recordings from the IAS, the AV node, and the IVS following a cut of the His bundle. The His bundle forms the only electrical connection between the AV node and the ventricle such that cutting fibers below the AV node effectively disconnected electrical coupling between the ventricular myocardium and inputs from the AV node. As shown in Fig. 11A, severing the His bundle eliminated the synchronous firing of ventricular BE and the detection of ventricular APs. This finding reinforced our interpreta- H838 A CONDUCTION BARRIER REGULATES AV DELAY tion that APs from the lower left zone of the preparation originate from ventricular cells. Under our experimental conditions, the His bundle cut produced a 2:1 block between atrial and nodal activation (Fig. 11A, node). As a result, some atrial activation waves (Fig. 11A, I) failed to entrain an AV node activation (Fig. 11B; note the lack of AV node activation), whereas the next beat (Fig. 11A, II) was coupled to an AV node activation (Fig. 11C). Not all activity ceased in the ventricular tissue because ectopic beats were occasionally observed, but these were out of phase with atrial and nodal APs. An advantage of His bundle cuts is the absence of ventricular APs, which interfered with our ability to detect the repolarization phase of N-cell APs. A conduction barrier between atrial and nodal cells regulates AV delay. AV delay is physiologically regulated by heart rate, parasympathetic, and sympathetic activity. To investigate the mechanisms responsible for the physiological regulation of the AV node delay, activation maps across the node were analyzed as a Table 2. Ratio of peak II to peak I as a function of depth of focus Depth, µm Peak II/Peak I 0 250 500 750 1,000 0.301 6 0.011 0.331 6 0.036 0.490 6 0.108 0.424 6 0.154 0.387 6 0.136 Values are means 6 SD of ratios of peak II to peak I from normalized signals recorded from 4 rabbit AV node preparations, with 3 cardiac beats averaged per preparation. Optical APs were recorded with focal plane at various depths below surface of AV nodal zone. Amplitudes of peaks I and II were measured from data printouts. Ratio of peak II to peak I first increased with increasing depth of focus (from 0 to 0.5 mm) then decreased (from 0.5 to 1.0 mm). Initial increase in ratio was due to an increase of peak II with increasing depth of focus rather than a decrease of peak I and was statistically significant (P , 0.05) by analysis of variance. DISCUSSION The study presents the first measurements of electrical propagation across the AV node using voltagesensitive dyes and imaging techniques. A number of technical difficulties were overcome, including signal-tonoise ratio, depth of focus, detection and analysis of three sequential optical upstrokes, and display of activation maps. Several lines of evidence were presented to support the interpretation that peak II represented AV nodal AP and could be used to map activation within the node. 1) The occurrence of peak II was anatomically correlated with the location of the midnodal and lower nodal regions (Fig. 9). 2) The temporal relationship between peak II and electrograms recorded from the IAS and IVS indicated that peak II originated from cells Downloaded from http://ajpheart.physiology.org/ by 10.220.32.247 on May 2, 2017 Fig. 7. Ratio of peak II to peak I as a function of depth. Schematic of a transverse histological section of rabbit AV node indicates different layers of cell types found as a function of depth in tissue [adapted from Janse et al. (10a)]. Volumetric elements (shaded cubes) superimposed on section represent volume of cells that are primary sources of fluorescence signals (F) detected by a diode when focal plane is shifted below surface in a stepwise manner. At different focal planes, different layers of cells become primary sources of signal. hn, Excitation light. function of cycle length. The heart rate was controlled by removing the SA node and pacing the right atrium with bipolar electrodes placed near the SA node. In this experimental group (n 5 9), cycle length was varied from 270 to 350 ms; shorter cycle lengths made it difficult to temporally separate the overlapping peaks I–III for analysis, and longer cycle lengths were possible except that extra beats occasionally interfered with the measurements. In this range of cycle lengths, activation maps across the IAS, AV node, and IVS maintained the same pattern. AV delay measured with atrial and ventricular electrograms changed with cycle length, as shown in Fig. 12, and was highly correlated with changes in t1, with a correlation coefficient of 0.98, but correlated poorly with changes in t2, with a correlation coefficient of 0.42 (each data point corresponds to the mean 6 SD). Heart rate and AV delay are regulated physiologically by sympathetic and parasympathetic innervation to the heart. Although much is known regarding the mode of action of neurotransmitter at the SA node, the mechanisms responsible for AV node regulation are less well understood. In a separate experimental group (n 5 5), we altered AV delay by perfusing the heart with ACh (0.1 µM). In all five experiments, the major change in the optical map was an experimentally significant increase in t1 by 19 6 2.3 ms and a slight increase in the time to activate the node [e.g., a decrease in apparent conduction velocity in the node region (QN )]. Figure 13, A and B, depicts the maps of activation of the IAS before and after perfusion with ACh, respectively. Note the similarities in activation patterns across the IAS and the node; the only marked change caused by ACh is an increase in the delay between atrial and nodal activation, with t1 increasing by 39.4% from 44.2 to 61.6 ms (Fig. 13, A and B). The prolongation of t1 with a slight decrease in QN suggests that ACh hyperpolarizes N cells such that the atrial current injected across a high-resistance barrier is less effective at reaching suprathreshold potential in the AV node. Thus a longer interval of current injection is required to reach threshold and elicit AV node activation. The increased AV delay caused by an ACh-induced hyperpolarization can be attributed to an injection of current across a highresistance barrier with passive electrical properties. A CONDUCTION BARRIER REGULATES AV DELAY H839 firing in the correct time frame for AV nodal APs. 3) The spread of activation analyzed from peak II reproducibly (in .28 hearts) produced a zone of slow conduction distinguishable from either atrial or ventricular propagation. 4) Under conditions in which the AV node was the primary pacemaker, peak II became the first upstroke to fire during a cardiac beat and produced the same spatiotemporal zone of activation as that observed when the AV node is driven by atrial inputs. 5) After the His bundle was cut, ventricular activation was blocked, but the spatiotemporal characteristics of peak II did not significantly change. 6) The signal amplitude of peak II as a function of depth of focus was consistent with the interpretation that peak II originated from the firing of APs by cells ,0.2–0.5 mm below the surface of the preparation. 7) Peak II signals (Fig. 3D) recorded from the posterior zone of the activation map were insensitive to TTX. Late signals (observed in ,50% of the preparations) consisting of a small depolarization preceding a ventricular AP represented the firing of His bundle APs and were TTX sensitive (Fig. 10). Peak II is not movement artifact. A major concern was the possibility that peak II was primarily caused by movement artifact (MA) rather than voltage-dependent fluorescence signals. Extensive measurements were carried out to negate that possibility. First, all measurements were obtained with the judicious use of DAM, an uncoupler of excitationcontraction coupling. It was important to find suitable conditions for the use of DAM (see Staining procedure) because perfusion with DAM for .30 min prolonged AV delays and reduced excitability of the preparation, and continued DAM perfusion eventually blocked all electrical activity (measured optically or with electrodes). On the other hand, DAM effectively blocked contractions after 8–10 min and could be washed out to reverse its effects. In this way, DAM perfusion for 10 min gave us 15–20 min to acquire data before washing out DAM. Second, the dF/dt of peak II produced a sharp, unique time point, whereas analysis of MAs by the same signal-processing technique produced neither a sharp maximum dF/dt (see Fig. 4) nor a single maximum time point so that an activation map could not be generated from MAs. Third, analysis of activation patterns from peak II were highly reproducible (.27 hearts) in the anatomic location, the direction of propagation, and the propagation velocity. MAs are typically unpredictable, and such a coincidence is statistically negligible. Fourth, in a few experiments, the Ca21 concentration (0.2–5 mM) was varied to produce vastly different levels of MAs, yet the spatiotemporal characteristics of peak II did not vary significantly. Fifth, when the AV node was the primary pacemaker, peak II fired first and no interference from MA from atrial and ventricular contractions was possible. In the latter case, the signals produced maps of electrical activity similar to those obtained when the AV node fired after the atrial activation. Finally, microelectrode impalements in the midnodal zone recorded N- or NH-cell APs that were coincident with the rise of peak II signals, which demonstrates that peak II depolarization originated from the firing of N and/or NH cells. Taken together, these findings give a high degree of confidence that peak II represented the electrical activity of a group of cells that fired APs in the correct anatomic location and at the right time. Downloaded from http://ajpheart.physiology.org/ by 10.220.32.247 on May 2, 2017 Fig. 8. Optical recordings from a pacing AV node. SA node was removed so that AV node became primary pacemaker. A: atrial and ventricular BEs were used to monitor firing of IAS and IVS and were temporally correlated with optical APs recorded from various regions of AV zone preparation: IAS (atrial AP), posterior boundary of AV node (transitional cells), AV node (midnodal cells), and IVS (ventricular AP). Earliest activation occurred at AV node (time line a) and propagated to atrium (time line b), His bundle, and ventricle (time line c). B: activation maps of AV node and IAS generated from atrial and nodal APs (axes are length in millimeters). Activation sequence in IVS was intentionally omitted to allow visualization of conduction through node and delay between nodal and IAS activation. Impulses initiated in node (*) and propagated across node in 10 ms and to IAS after a 17.96-ms delay. Step delay in excitation wave occurred across a conduction barrier delineated by closely packed isochronal lines (arrow). Isochronal lines were 2 ms apart. For web site access to QuickTime movie of propagation sequence, see MATERIALS AND METHODS. H840 A CONDUCTION BARRIER REGULATES AV DELAY Identification of various AV node cell types. Previous electrophysiological and morphological studies have identified at least five different cell types in the AV node zone. A major limitation of the optical technique is the difficulty of resolving single cell APs by optical mapping. However, drug interventions such as TTX proved to be useful in identifying the cell types comprising the activation zone delineated by peak II and His depolarization (Figs. 9 and 10). For example, TTX had no effect on propagation in the proximal region (Fig. 10A) but slowed propagation in the distal region. This suggests that peak II consists primarily of N and/or NH cells with few TTX-sensitive AN or His cells, whereas the distal region contained TTX-sensitive His cells. Another limitation of the optical techniques was the inability to measure AP durations because signals from the compact node consisted of superimposed atrial, nodal, and ventricular APs. We attempted to resolve the complete time course of N-cell APs (including the repolarization phase) by increasing the optical magnification and varying the depth of focus. With the present optical configuration, we failed to selectively measure N-cell APs, even at the expense of decreased signal-tonoise ratio. An alternative approach used here was to cut the His bundle to block ventricular APs and measure N-cell AP durations. However, cutting the His Downloaded from http://ajpheart.physiology.org/ by 10.220.32.247 on May 2, 2017 Fig. 9. Superimposition of activation maps and histological landmarks of AV node. A: charge-coupled device images of 3 longitudinal sections of same AV node taken at different depths. Anatomic landmarks are IAS, IVS, CT, fat deposits (F), midnodal region (N), and His bundle. B: activation maps recorded from same AV node showing sequence of depolarization at 3.2-ms intervals, as in Fig. 6. Within a single cardiac beat, progression from earliest (top left) to latest (bottom right) activation map shows sequence of depolarization. C: superimposition of activation map derived from peak II depolarization on a longitudinal cross section of same AV node preparation. Optical and histological maps were aligned as described in MATERIALS AND METHODS and indicate that a barrier to electrical propagation forms a collar at posterior edge of AV node (may include zones of adipose tissue) and that a bridge with a high electrical resistance appears to separate IAS from midnodal region. A CONDUCTION BARRIER REGULATES AV DELAY H841 bundle could change the electrotonic coupling between NH and His fibers and thereby alter N-cell AP durations. Despite these limitations, optical mapping techniques offer a new approach for studying the organization and coupling of A, AN, N, NH, and His cells in ways that cannot be resolved by conventional electrode techniques. Activation patterns across AV node region. The depolarizations of peaks I–III delineate the spread of AP propagation across the atrial, nodal, and ventricular tissue, respectively. The earliest depolarizations (peak I) were synchronous with the atrial electrogram and mapped the sequence of depolarization across the IAS, overlapping the AV node region. The time course, distribution, and propagation velocity of these APs identified them as atrial in nature, with AN characteristics as they overlap the AV node zone. After the last AN cell depolarization, there was a delay of 30–40 ms (during which no other cell depolarizations could be detected) followed by depolarizations within the node, detected by peak II. This delay between peak I and II indicated that conduction was discontinuous between AN and midnodal cells. The zone delineated by peak II represented the firing of midnodal cells in the posterior region of the node and lower nodal cells in the more anterior region of the AV node (Fig. 9). In ,50% of the preparations, His bundle APs were detected as small depolarizations that preceded ventricular APs and de- lineated a narrow track of electrically active tissue contiguous with the lower nodal region. In line with this interpretation, there was a progressive increase in conduction velocity from the posterior region (midnode) to the anterior track (His bundle) of the AV node. Thus, for the first time, optical techniques provided detailed maps of the sequence of activation across the AV node and described the organization of the various cell types comprising the node. The data also indicated that there was a discontinuity in conduction, with a well-defined anatomic location between atrial and midnodal cell depolarization. Mechanism(s) responsible for AH intervals. A number of important conclusions can be extracted from the present findings. One major finding is that the conduction through the AV node is not decremental but discontinuous, and we conclude that a conduction barrier exists between the atrial and midnodal cells. Features of this barrier are consistent with the presence of an inexcitable gap across which activation proceeds electrotonically through a high-resistance pathway. Although a barrier consisting of resistance and capacitance components is consistent with the data, we have not excluded the possibility of a barrier with more complex electrical properties, with possible modulation by neurotransmitters. The superimposition of activation maps on longitudinal cross sections of the AV node suggests that the electrical barrier surrounds Downloaded from http://ajpheart.physiology.org/ by 10.220.32.247 on May 2, 2017 Fig. 10. Effect of TTX (10 µM) on activation across AV node and His bundle. Excitation maps across AV node were generated from activation time points of peak II, as in Fig. 3, and isochronal lines were drawn 1 ms apart. A: in some preparations, activation maps generated from intermediate (peak II) depolarization revealed a pattern of depolarization across midnodal, lower nodal, and His bundle regions (left). Optical signals (right) measured from midnodal region (AV node) exhibited typical response characteristics of peak II. Signals from lower node region (not shown) were similar to midnodal signals. Signals from His bundle were typically recorded from zones exhibiting 2 sequential depolarizations: upstrokes generated by His firing followed by firing of ventricular cells. Time lines labeled A, N, and H represent the depolarization time of the IAS, node, and His bundle, respectively. B: to confirm interpretation of these signals, TTX was used to identify nature of cells producing optical signals. Activation maps were drawn in absence of TTX (left) and then recorded after 20 min of perfusion with 10 µM TTX. Note that conduction across posterior segment of node was TTX insensitive, whereas conduction through anterior segment slowed markedly, supporting interpretation that these responses originated from His bundle. H842 A CONDUCTION BARRIER REGULATES AV DELAY the node and lies at the boundary of the midnode and the IAS. Zones of fat deposits (adipose cells) adjacent to the posterior and anterior zones of the AV node surround the AV node in 3-D, according to serial longitudinal sections, and are typically found in rabbit (6) and Fig. 12. Correlation of t1 and AV delay at various cycle lengths. A heart was paced from 270 to 350 ms with bipolar electrodes placed near SA node; t1 and t2 were measured as described in Fig. 4. Total AV delay (measured with surface electrograms), t1, and t2 were plotted as a function of cycle length. Each set of data points was fitted with a line drawn using polynomial curve fit. Step delay in conduction between A (and/or AN) and N cells (t1 ) changed with rate, whereas t2 was within experimental error and rate independent. Correlation coefficient between AV and t1 delay as a function of cycle length was 0.98, and that between AV and t2 was 0.39. human AV nodes (personal communication, Dr. L. C. Nichols, School of Medicine, University of Pittsburgh, PA). These fat deposits likely form an effective electrical barrier that insulates the AV node from the IAS. As a result, electrical coupling to the AV node can only occur via narrow bridges: one bridge connects to the IAS and the other to the cristae terminalis that are unobstructed by fat deposits. A mathematical simulation of optical signals was developed to predict the signal characteristics under different conditions such as synchronous, decremental, or discontinuous conduction. The model consisted of 15 cells of equal dimensions firing optical APs of equal amplitude and time course, and all 15 are detected with one diode. When all 15 cells fired APs synchronously, the resulting optical AP had a sharp upstroke with a rise time similar to that of the single cell upstroke (Fig. 14A). When the cells fired with a smooth conduction delay (50 ms across 15 cells), as in decremental conduction, the resulting optical AP had a slower rise time indicative of the time-averaged upstrokes of asynchronous depolarizations (Fig. 14B). Only when there was an abrupt delay (e.g., 45 ms) between APs from the first 10 cells and the last 5 cells did the resulting optical APs exhibit two distinguishable upstrokes (Fig. 14C, arrow). Thus decremental conduction can be excluded as an AV node delay mechanism, because the resulting optical APs from the atrium and the node would blend into a single upstroke (not 3 temporally distinct spikes) representing the time-averaged sum of APs (Fig. 14B). Decremental conduction is also incompatible with our Downloaded from http://ajpheart.physiology.org/ by 10.220.32.247 on May 2, 2017 Fig. 11. Effect of cutting His bundle on electrical activity was investigated in 4 rabbit AV node preparations. A: simultaneous recordings of atrial and ventricular BEs and optical APs from IAS (atrium), AV node, and IVS (ventricle) are shown after His bundle was cut. As expected, His bundle cut blocked excitation to IVS, abolishing ventricular BEs and APs. In absence of coupled ventricular depolarization, AV node signals fired 2 sequential spikes, an atrial and a nodal spike; the latter was not followed by a ventricular AP, revealing plateau and repolarization phases of AV node AP. Atrial APs were synchronous with atrial BE, but nodal APs were markedly reduced at every other beat, suggesting a 2:1 block of AV conduction. Activation maps of 2 successive beats (beats I and II on nodal AP) are shown, respectively, in B and C. Activation map produced by beat I produced an IAS excitation pattern (B), whereas beat II mapped IAS and nodal activation. Note that IAS activation patterns were similar for beats I and II. Isochronal lines were 1 ms apart; axes are length in millimeters. A CONDUCTION BARRIER REGULATES AV DELAY H843 Fig. 13. Effect of neurotransmitter ACh on AV node delay was investigated by perfusing heart with 0.1 µM ACh. AV delay increased within seconds after ACh perfusion, and data were acquired 2–5 min after ACh injection (n 5 5). Isochronal lines were 2 ms apart; axes are length in millimeters. A: control activation map recorded immediately before ACh injection. B: activation map recorded after 3 min of perfusion with ACh. First nodal activation occurred at 44.2 ms in control (A) and at 61.6 ms in presence of ACh (B). node conduction (11, 13, 16). Discontinuous conduction was also tested by applying a local perturbation [i.e., current (27), high K1 (5), freezing (26), or a sucrose gap (2)] in otherwise uniform cardiac fibers (e.g., Purkinje or ventricular). In such experimental models, the perturbation produced a step change in delay and ‘‘stagnation’’ similar to that observed in the node. Step delays were caused by an interruption of active transmission of an impulse as it arrived at the inexcitable gap. The electrotonic transmission through the inexcitable gap slowly charged distal cells until the resting potential of the distal cells reached threshold and ignited active transmission in the distal cells. It is critically important that the step delay is determined by the time needed to inject the electrotonic current necessary to induce active transmission in the distal cells. James et al. (10) also argued against decremental conduction because AV delays were not proportional to the size of the AV node in various mammalian hearts. They proposed that pacemaker cells are involved in the AV delay as coupled relaxation oscillators modulated by electrotonic atrial inputs. The location and nature of the inexcitable gap and of discontinuous conduction are still controversial. Early studies (18) indicated that the major component of AV delay occurred between the firing of late AN and early N cells. Subsequent findings by Billette et al. (3, 4) Fig. 14. An experimentally recorded atrial AP was used to simulate optical AP recordings during synchronous, decremental, and discontinuous activation of 15 cells with equal dimensions and signal amplitudes. A: synchronous activation of 15 cells. Optical recording is a single spike representing sum of 15 APs. B: decremental conduction of 15 cells. Cells were coupled through a high-resistance barrier, resulting in graded delays (50 ms) evenly distributed from first to last cell. Optical AP is a single spike with a slow rise time caused by asynchronous depolarization of the 15 cells. C: discontinuous activation between first 10 cells (‘‘early activating cells’’) and last 5 cells (‘‘late activating cells’’). This model was based on a single step delay due to a high-resistance, inexcitable barrier, resulting in discontinuous conduction (10 early activating and 5 late activating cells). Time to propagate across early and late activating cells was 5 ms, and there was a 45-ms delay between early and late cells. In discontinuous conduction, optical AP will exhibit 2 distinct upstrokes that are temporally separated according to step delay across inexcitable gap. Downloaded from http://ajpheart.physiology.org/ by 10.220.32.247 on May 2, 2017 measurements of QN (Table 1). For instance, if conduction through the node is decremental (with no ‘‘step’’ changes in conduction) and accounts for AV delays, then QN should be at least 10 times slower than in the atrium. From the analysis of optical signals, the total AV delay was the sum of 1) the interval between A- and N-cell activation (t1 ), 2) the time to propagate across the node, and 3) the time between N- and V-cell activation (t2; see Table 2). The conduction velocity in the node was 0.162 6 0.024 m/s, approximately onethird that of atrial tissue (0.76 6 0.062 m/s). However, the delay due to conduction through the node was #14 ms, an insufficient interval to account for the AV delay. On the other hand, delay due to discontinuous conduction between peaks I and II was 44.24 ms or ,50% of AV delays. On the basis of these results, it can be postulated that the major AV nodal delay is not due to the slow conduction inside the node but to discontinuous conduction between peaks I and II. Moreover, when the AV node became the primary pacemaker, the conduction barrier at the posterior margin of the AV node was still responsible for the major component of AV delay during retrograde as well as anterograde propagation (n 5 3). The concept of an inexcitable gap in the node, resulting in discontinuous conduction, was inferred from intracellular electrode studies (3) and simulations of AV H844 A CONDUCTION BARRIER REGULATES AV DELAY The authors are grateful for the inspiration of the late Richard A. Lombardi. The authors thank Dr. Gregory Kloehn for technical assistance and William Hughes, departmental machinist, for construction of the heart chamber and manipulators to adjust the focal plane of the optical apparatus and to position stimulating and surface recording electrodes. Thanks are also due Drs. Ronald L. Hamilton and Lawrence C. Nichols (Dept. of Pathology, University of Pittsburgh, PA) for technical support and guidance regarding analysis and interpretation of the AV node histology. Address for reprint requests: G. Salama, Dept. of Physiology, School of Medicine, Univ. of Pittsburgh, 3500 Terrace St., Room 2, 314 Biomedical Science Tower, Pittsburgh, PA 15261. Received 7 February 1997; accepted in final form 22 August 1997. REFERENCES 1. Anderson, R. H., M. J. Janse, F. J. L. Van Capelle, J. Billette, A. E. Becker, and D. Durrer. A combined morphologi- cal and electrophysiological study of the atrioventricular node of the rabbit heart. Circ. Res. 35: 909–922, 1974. 2. Antzelevitch, C., and G. K. Moe. Electrotonically mediated delayed conduction and reentry in relation to ‘‘slow response’’ in mammalian ventricular conduction tissue. Circ. Res. 49: 1129– 1139, 1981. 3. Billette, J. Atrioventricular nodal activation during premature stimulation of the atrium. Am. J. Physiol. 252 (Heart Circ. Physiol. 21): H163–H177, 1987. 4. Billette, J., M. J. Janse, F. J. L. van Capelle, R. H. Anderson, P. Touboul, and D. Durrer. Cycle-length-dependent properties of AV nodal activation in rabbit hearts. Am. J. Physiol. 231: 1129–1139, 1976. 5. Cranefield, P. F., H. O. Klein, and B. F. Hoffman. Conduction of the cardiac impulses. 1. Delay, block, and one-way block in depressed Purkinje fibers. Circ. Res. 28: 199–219, 1971. 6. DeFelice, L. J., and C. E. Challice. Anatomical and ultrastructural study of the electrophysiological atrioventricular region of the rabbit. Circ. Res. 24: 457–474, 1969. 7. Efimov, I. R., D. T. Huang, J. M. Rendt., and G. Salama. Optical mapping of repolarization and refractoriness from intact hearts. Circulation 90: 1469–1480, 1994. 8. Gettes, L. S., J. W. Buchanan, Jr., T. Saito, Y. Kagiyama, S. Oshita, and T. Fujino. Studies concerned with slow conduction. In: Electrophysiology and Arrhythmias, edited by D. P. Zipes and J. Jalife. Orlando, FL: Grune and Stratton, 1985, p. 81–87. 9. Hoffman, B. F., and P. F. Cranefield. Electrophysiology of the Heart. New York: McGraw-Hill, 1960. 10. James, T. N., K. Kawamura, F. L. Meijler, S. Yamamoto, F. Terasaki, and T. Hyashi. Anatomy of the sinus node, AV node, and His bundle of the heart of the sperm whale (Physeter macrocephalus), with a note on the absence of an os cordis. Anat. Rec. 242: 355–373, 1995. 10a.Janse, M. J., F. J. L. van Capelle, R. H. Anderson, P. Touboul, and J. Billette. Electrophysiology and structure of the atrioventricular node of the isolated rabbit heart. In: The Conduction System of the Heart: Structure, Function and Clinical Implications, edited by H. J. J. Wellens, K. I. Lie, and M. J. Janse. Leiden, The Netherlands: Stenfert Kroese, 1976, p. 296–315. 11. Kinoshita, S., and G. Konishi. Mechanisms of atypical atrioventricular Wenchebach periodicity. A theoretical model derived from the concepts of inhomogeneous excitability and electrotonically mediated conduction. J. Electrocardiol. 22: 227–233, 1989. 12. Kokubu, S., M. Nishimura, A. Noma, and H. Irisawa. Membrane currents in the rabbit atrioventricular node cell. Pflügers Arch. 393: 15–22, 1982. 13. LeBlanc, A. R., and B. Dube. Propagation in the AV node: a model based on a simplified two-dimensional structure and a bidomain tissue representation. Med. Biol. Eng. Comput. 31: 545–565, 1993. 14. Loew, L., L. Cohen, J. Dix, E. Fluhler, V. Montana, G. Salama, and W. Jian-Young. A naphthyl analog of the aminostyryl pyridinium class of potentiometric membrane dyes shows consistent sensitivity in a variety of tissue, cell and model membrane preparation. J. Membr. Biol. 130: 1–10, 1992. 15. Luna, L. G. Manual of Histological Staining Methods of the Armed Forces Institute of Pathology (3rd ed.). New York: McGrawHill, 1968, p. 36–38. 16. Malik, M., D. Ward, and A. J. Camm. Theoretical evaluation of the Rosenblueth hypothesis. Pacing Clin. Electrophysiol. 11: 1250–1261, 1988. 17. Meijler, F. L., and M. J. Janse. Morphology and electrophysiology of the mammalian atrioventricular node. Physiol. Rev. 68: 608–647, 1988. 18. Paes de Carvalho, A., and D. F. De Almeida. Spread of activity through the atrioventricular node. Circ. Res. 8: 801–809, 1960. 19. Piller, H. Microscope Photometry. New York: Springer-Verlag, 1977, p. 16. 20. Salama, G. Optical measurements of transmembrane potentials in heart. In: Spectroscopic Probes of Membrane Potential, edited by L. Lowe. Boca Raton, FL: CRC Uniscience, 1988, p. 132–199. Downloaded from http://ajpheart.physiology.org/ by 10.220.32.247 on May 2, 2017 showed that, at short cycle lengths, the interval between an atrial electrogram and the latest AN cell had not changed; the delay occurred between the latest AN cell and the earliest NH cell. Moreover, at a short cycle length (130 ms), N-cell AP dissociated into two components: the first component coincided with the AP upstroke of late AN cells, and the second component coincided with the AP upstroke of the earliest NH cells. No AP upstrokes were found in between. The conclusion was that, at a short cycle length, the N zone (approximately equivalent to the compact node) became inexcitable and only transmitted electrotonic currents that slowly brought distal NH cells to threshold. This implied that N cells were the actual barrier (3, 4). The present optical data are in agreement with the early studies of AV propagation (18) and showed that, under a wide range of physiological rates (250- to 500-ms cycle lengths), the major component of delay resides between the latest AN and earliest N cells. In our intact heart preparations, attempts to drive the AV node at higher rates produced unstable AV coupling, making it unsuitable to reexamine the observation of Billette et al. (4). The present study provides robust evidence for a step jump in conduction across a well-defined anatomic gap. The regulation of the time delay across this gap as a function of cycle length and ACh indicates that delays across this gap play a major mechanistic role in the physiological control of AV delay. Prolongation of AV delay by ACh is consistent with the notion proposed by James et al. (10) that the AV node is a pacemaker modulated by electrotonic inputs. The latter interpretation is based on a hyperpolarization of AV nodal cells by ACh such that a greater input of current is required across the inexcitable gap to obtain a suprathreshold depolarization that can activate the AV node. Thus the present data are incompatible with a decremental conduction mechanism and provide direct evidence for the existence of a conduction barrier, its specific anatomic location, and a measurement of the step delay (t1 ) involved in the physiological regulation of AV delay. This study raises new questions regarding the mechanisms underlying t1 and AV delays and demonstrates the potential of optical techniques to elucidate basic problems in AV node physiology and salient clinical problems. A CONDUCTION BARRIER REGULATES AV DELAY 21. Salama, G., A. Kanai, and I. R. Efimov. Subthreshold stimulation of Purkinje fibers interrupts ventricular tachycardia in intact hearts. Circ. Res. 74: 604–619, 1994. 22. Salama, G., R. Lombardi, and J. Elson. Maps of action potential and NADH fluorescence in intact working hearts. Am. J. Physiol. 252 (Heart Circ. Physiol. 21): H384–H394, 1987. 23. Salama, G., and M. Morad. Merocyanine-540 as an optical probe of transmembrane electrical activity of the heart. Science 191: 485–487, 1976. 24. Sano, T., M. Tasaki, and T. Shimamoto. Histologic examination of the origin of the action potential characteristically obtained from the region bordering the atrioventricular node. Circ. Res. 7: 700–704, 1959. H845 25. Tawara, S. Das Reizleitungs System des Herzens. Jena, Germany: Fisher, 1906. 26. Waxman, M. B., E. Downar, and R. W. Wald. Unidirectional block in Purkinje fibers. Can. J. Physiol. Pharmacol. 58: 925– 933, 1980. 27. Wennemark, J. R., V. J. Ruesta, and D. A. Brody. Microelectrode study of delayed conduction in the canine right bundle branch. Circ. Res. 23: 753–769, 1968. 28. Young, M.-L., B. M. Ramaza, R. C. Tan, and R. W. Joyner. Adenosine and hypoxia effects on atrioventricular node of adult and neonatal rabbit hearts. Am. J. Physiol. 253 (Heart Circ. Physiol. 22): H1192–H1198, 1987. Downloaded from http://ajpheart.physiology.org/ by 10.220.32.247 on May 2, 2017