Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Herpes simplex virus wikipedia , lookup
Schistosomiasis wikipedia , lookup
Dirofilaria immitis wikipedia , lookup
Trichinosis wikipedia , lookup
Hepatitis C wikipedia , lookup
Human cytomegalovirus wikipedia , lookup
Coccidioidomycosis wikipedia , lookup
Neonatal infection wikipedia , lookup
Sarcocystis wikipedia , lookup
Oesophagostomum wikipedia , lookup
Hepatitis B wikipedia , lookup
Mycoplasma pneumoniae wikipedia , lookup
Lymphocytic choriomeningitis wikipedia , lookup
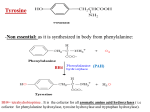
Journal of Antimicrobial Chemotherapy (2005) 56, 861–868 doi:10.1093/jac/dki331 Advance Access publication 19 September 2005 Serotonin and melatonin, neurohormones for homeostasis, as novel inhibitors of infections by the intracellular parasite chlamydia Mohd. Akhlakur Rahman1, Yoshinao Azuma1*, Hajime Fukunaga2, Tomoyuki Murakami2, Kazurou Sugi2, Hideto Fukushi3, Koshiro Miura1, Harumi Suzuki1 and Mutsunori Shirai1 1 Department of Microbiology, Yamaguchi University School of Medicine, 1-1-1, Minamikogushi, Ube, Yamaguchi 755-8505, Japan; 2Department of Clinical Research, National Sanyou Hospital, Yamaguchi 755-0241, Japan; 3Department of Veterinary Microbiology, Gifu University, 1-1 Yanagido, Gifushi, Gifu 501-1193, Japan Received 4 July 2005; returned 29 July 2005; revised 16 August 2005; accepted 19 August 2005 Objectives: Chlamydiae are obligate intracellular bacteria, causing a variety of diseases, i.e. pneumonia, sexually transmitted disease, conjunctivitis and zoonosis. Tryptophan depletion by interferon-gamma (IFN-g) is the most important host defence system against chlamydial infection. Thus chlamydial tryptophan metabolism is thought to play key roles for IFN-g resistance, persistent infection and host/tissue tropisms. We tested tryptophan derivatives for activity against chlamydia-infected cells. Methods: Rates of chlamydial infection and sizes of the inclusions were evaluated by in vitro infection using three Chlamydiaceae species, Chlamydia trachomatis, Chlamydophila pneumoniae and Chlamydophila felis, which show significant divergence of tryptophan synthesis genes and different susceptibilities to IFN-g. Results: Melatonin and serotonin, which are recognized as neural hormones for maintenance of organism homeostasis, reduced chlamydial infection but not other bacterial growth tested here. Unlike IFN-g, melatonin limited infection of all three chlamydiae and the effects were not recovered by tryptophan supplementation. Melatonin treatment only of host cells could diminish infection and the infection reduction was neutralized by a pertussis toxin, an inhibitor of G proteins. Ligands of melatonin and serotonin receptors also hampered infection. Conclusions: Inhibition mechanisms of chlamydial infection by melatonin and serotonin appear to be different from those of IFN-g and involve specific G-protein-coupled receptors. Melatonin is deemed to inhibit early progression of the chlamydial development cycle, such as establishment of intracellular infection and/or conversion from elementary body to reticulate body. Utilization of melatonin, serotonin or their derivatives may be advantageous for harmless prevention of chlamydial infection. Keywords: Chlamydophila pneumoniae, Chlamydia trachomatis, tryptophan, antimicrobial agents, G-protein coupled receptor Introduction Chlamydiae are obligate intracellular prokaryotic pathogens, exhibiting a unique developmental cycle consisting of two major cell phases: the infectious but metabolically inert elementary body (EB) and the metabolically active but non-infectious reticulate body (RB).1 The chlamydiae cause a variety of diseases ranging from asymptomatic to serious diseases in humans and animals.2 Chlamydia trachomatis is a highly prevalent cause of ocular and urogenital diseases. Chlamydophila pneumoniae is responsible for acute and chronic respiratory diseases, and is widely considered to be a cause of atherosclerosis,3 asthma,4 Alzheimer’s disease5,6 and other inflammatory processes. Chlamydophila species, which formerly belonged to Chlamydia ............................................................................................................................................................................................................................................................................................................................................................................................................................. *Corresponding author. Tel: +81-836-22-2227; Fax: +81-836-22-2415; E-mail: [email protected] ............................................................................................................................................................................................................................................................................................................................................................................................................................. 861 The Author 2005. Published by Oxford University Press on behalf of the British Society for Antimicrobial Chemotherapy. All rights reserved. For Permissions, please e-mail: [email protected] Rahman et al. psittaci as subspecies, cause several zoonoses, for example, Chlamydophila felis is a worldwide pathogen of cats responsible for ocular and pulmonary diseases.7 One of the most important host defence systems against the chlamydial infection is interferon-gamma (IFN-g) synthesis, which induces indoleamine 2,3-dioxygenase (IDO) and thus depletes tryptophan from human cells, since many human pathogenic chlamydiae rely on the tryptophan.8–11 Besides the function of IFN-g-inducible tryptophan shortage as a host defence, insufficient IFN-g exposure may lead the chlamydial growth to a persistent state, which is characterized by atypical inclusions that are generally smaller in diameter and contain larger aberrant RBs.12 This is why chlamydial IFN-g resistance and host/tissue tropism are thought to depend on chlamydial tryptophan synthesis.13,14 Persistent chlamydial infection is thought to be involved in the pathophysiology of some chronic inflammatory diseases, e.g. asthma and atherosclerosis for C. pneumoniae, and blindness and infertility for C. trachomatis.15 Human tryptophan catabolism mainly consists of two pathways, the kynurenine pathway and the serotonin pathway. In the kynurenine pathway, tryptophan is processed to N-formylkynurenine by two types of rate-limiting enzymes, tryptophan 2,3-dioxygenase (TDO) induced by tryptophan in the liver and the IDO induced by IFN-g during the immune response in many tissues. Tryptophan is finally metabolized to an essential cellular cofactor, nicotinamide adenine dinucleotide (NAD+). The other pathway is the serotonin pathway that produces a variety of neural activators such as a neurotransmitter, serotonin and, a pineal hormone, melatonin.10 Serotonin is a well-known ubiquitous monoamine, which is derived from tryptophan and functions as a neurotransmitter, hormone and mitogenic factor that mediates a wide range of physiological activities in different cells through multiple receptor subtypes. Serotonin is produced and stored within several tissues throughout the human body, such as the central neural cells,16 blood platelets,17 gastrointestinal enterochromaffin cells18 and prostatic neuroendocrine cells.19 Furthermore, serotonin plays a fundamental role in regulation of growth, differentiation and gene expression.20–22 Melatonin, originally identified as an effector for circadian rhythms,23 is now known to be a hormone involved in a vast range of homeostasis maintenance activities, for example seasonal timing, sexual development, the antioxidant defence system and immune response.24–26 Melatonin is synthesized from tryptophan within the serotonin pathway mainly in the pineal gland, and in a number of extrapineal organs such as retina, lens, bone marrow, intestine, skin and so on.27 To date, three mammalian melatonin receptors, G-protein coupled receptors MT1 and MT2, and a quinone reductase family receptor, MT3, have been identified.19,28,29 Recently, several chlamydial genome sequencing projects have been completed and have revealed the divergence of the gene repertoire for tryptophan synthesis.13,14,30 It is not known why chlamydial species contain different tryptophan gene repertoires, but it is possible that some host factors related to tryptophan naturally restrict gene compositions of the chlamydial tryptophan operon in chlamydial evolution. Here we report the finding that the tryptophan derivatives serotonin and melatonin reduce chlamydial infection in HEp2 cells in a manner different from the IFN-g-inducible host defence. Materials and methods Chemicals L-Tryptophan, and its derivative compounds, such as 5-hydroxy tryptophan, 5-hydroxy tryptophol, 5-methoxyindole acetic acid, melatonin and serotonin, and ligands of melatonin and serotonin receptors, IIK7, luzindole, buspirone and NAN190, were purchased from Sigma– Aldrich (Saint Louis, MO). Gentamicin, cycloheximide, DAPI and Dulbecco’s modified Eagle medium (DMEM) were purchased from Sigma–Aldrich. Pertussis toxin was purchased from List Biological Laboratories, Inc. (Campbell, CA), human recombinant IFN-g was from Peprotech Ec Ltd (London, UK), and fetal calf serum (FCS) was from Cansera International Inc. (Etobicoke, Canada). The family Chlamydiaceae-specific fluorescein isothiocyanate (FITC)-labelled antibody was from Denkaseiken (Tokyo, Japan). Cell line and bacterial strains HEp-2 cells (ATCC CCL-23) were grown in HEp-2 medium (DMEM supplemented with 10% heat-inactivated FCS and 50 mg/L gentamicin) at 37 C with 5% CO2. C. pneumoniae J138 isolated in Japan in 1994,30 C. felis Japanese strain Fe/C-5631 and C. trachomatis serovar D were used in this work. These strains were purified by sucrose-gradient centrifugation and stored at –80 C in the SPG buffer (pH 7.2), which consists of 250 mM sucrose, 10 mM sodium phosphate and 5 mM glutamate. Chlamydial titres were adjusted for C. pneumoniae J138 to 2.0 · 108 inclusion formation units (IFU)/mL, and C. felis and C. trachomatis to 1.0 · 108 IFU/mL. Other bacterial strains used here are Escherichia coli, Staphylococcus aureus, Bacillus subtilis E192, Streptococcus pneumoniae, Pseudomonas aeruginosa PAO1, Helicobacter pylori CPY3401, Mycobacterium tuberculosis H37Rv, and Legionella pneumophila ATCC33153. Chlamydial infection Chlamydial infection was performed by methods described elsewhere.32 Briefly, 2.0 · 104 HEp-2 cells in 0.10 mL of the medium were seeded into each well of flat-bottomed 96-well tissue culture plates, and allowed to adhere for 24 h prior to use. Infection was performed by addition of chlamydial EBs to achieve a multiplicity of infection (MOI) of 0.20. In this condition, inclusion formation units, number of host cells and infection rate may turn to 1 · 105 IFU/mL, 5 · 104 cells and 20%, respectively. Relative infection rate was determined as an average of at least three replicate tests and confirmed with two other independent experiments performed under the same conditions. After centrifugation at 700g for 60 min at 22 C and then incubation for 30 min at 36 C with 5% CO2, the inoculum was replaced with the post-infection medium (DMEM with 5% heat-inactivated FCS, 50 mg/L gentamicin and 1 mg/mL cycloheximide). The infected cells were incubated for 48 h at 36 C with 5% CO2, following fixation of the infected cells for 30 min in methanol. For measurement of IFUs and rates of infected host cells, the infected cells were stained with the family Chlamydiaceae-specific FITC-labelled antibody for 60 min at 37 C, and then stained with 0.1 mg/mL of DAPI for 10 min. Numbers of host cell nuclei and chlamydial inclusions were counted under fluorescence microscopy, BX50 (Olympus, Japan) at a magnification of ·200. Under treatments with chemicals and IFN-g, typical and atypical (or small) inclusions were observed. Here, inclusions larger than approximately 1 mm in diameter were counted as successful infections.33,34 862 Serotonin and melatonin inhibit chlamydial infection Chemical treatment (a) In most experiments, treatments with 50–200 mM test compound were started at 24 h prior to infection and ended at 48 h post-infection, otherwise treatment schedules and concentrations of chemicals are described in the figure legends for each experiment. For other bacteria, E. coli, S. aureus, B. subtilis, P. aeruginosa were cultured in LB broth, whereas S. pneumoniae was cultured in Brain Heart Infusion and H. pylori in Brucella broth supplemented with 5% horse serum and 0.1% b-cyclodextrin. Inocula of each strain were incubated in their respective media treated with melatonin, gentamicin or mock at 37 C, and then the OD at 600 nm was measured as the indication for bacterial growth up to 8 h except H. pylori to 24 h. The BACTEC MGIT 960 Automated System (Becton Dickinson, Cockeysville, MD) was adapted for susceptibility tests with M. tuberculosis and L. pneumophila. For L. pneumophila growth, Legionella BCYE growth supplement (Oxoid Ltd, Hampshire, UK) was added to the MGIT medium for M. tuberculosis. 5-hydroxyB tryptophan KynurenineC SerotoninD MelatoninE 5-hydroxyF tryptophol Relative infection rate (%) 100 80 * 60 * 40 20 0 – A B C D E F G 120 Relative infection rate (%) (c) Screening of agents against chlamydial infection in vitro 5-methoxyindoleG acetic acid 120 (b) Results Elevation of IFN-g levels by melatonin has been reported during viral infection in a mouse model.35 To investigate the relationship between melatonin and IFN-g, excess tryptophan was added independently to melatonin and IFN-g in chlamydial infection. The inhibition of chlamydial infection by IFN-g was observed as previously reported33 and was reversed by adding tryptophan, consistent with the well-known schema of tryptophan depletion by IFN-g.9,11 In contrast, tryptophan addition did not reverse melatonin-mediated inhibition (Figure 2a). This suggests that the inhibitory mechanism of melatonin is different from that of IFN-g in this case. L-formyl kynurenine NAD To analyse the production of infectious progeny under the treatment of melatonin, serotonin and IFN-g, chlamydial progeny were collected from the culture medium at 72 h post-infection by centrifugation at 10 000g for 15 min at 4 C. To measure the amount of infectious progeny, the collected chlamydiae were suspended in the DMEM buffer and adapted to the secondary infection using HEp-2 cells growing in 96-well plates. The procedures described above were used to determine the infection rates. Effect of melatonin and IFN-g on chlamydial infection Serotonin pathway 5-hydroxyindole acetaldehyde Assay of infectious progeny Tryptophan and six derivatives (Figure 1a) were administered at 50 mM onto HEp-2 cells infected with C. pneumoniae J138 at 24 h prior to infection and maintained until the infection was terminated by fixation. The rates of infected host cells were calculated and are shown in Figure 1(b). Among the compounds tested here, serotonin and melatonin reduced the chlamydial infection by 50% compared with controls. 5-Hydroxytryptophan at 200 mM also showed an inhibitory effect on chlamydial infection (data not shown). Optimum concentrations of melatonin and serotonin were analysed in a range of 25–400 mM, at which no host toxicity was observed under microscopic studies. Treatments with melatonin and serotonin caused a dose-dependent inhibition of C. pneumoniae infection of HEp-2 cells and at 100 mM both reached the optimum effect (Figure 1c). In most experiments below, melatonin and serotonin were used at 100 mM. L-TryptophanA Kynurenine pathway Melatonin Serotonin 100 80 60 40 20 0 0 25 50 100 200 Concentration (µM) 400 Figure 1. Effects of tryptophan and its metabolites on chlamydial infection in vitro. (a) Schematic presentation of tryptophan and its metabolites in kynurenine and serotonin mammal pathway. (b) Effects of tryptophan and its metabolites, indicated as A–G in panel (a), were tested at 50 mM each on infection of C. pneumoniae J138 to human HEp-2 cells. (c) Dose tests were carried out with 25–400 mM melatonin and serotonin. In both cases of panels (b) and (c), HEp-2 cells were infected with C. pneumoniae J138 at 0.2 multiplicity of infection (MOI). The rates of infected host cells were determined at 48 h post-infection by staining of chlamydial inclusions with the chlamydia-specific FITC-labelled antibody and host nuclei with DAPI. Each infection rate was normalized with controls to the relative infection rate, while the actual infection rates of the controls were 11.9% and 19.4% in panels (b) and (c), respectively. None of the chemicals showed substantial toxicity to the host cells under these conditions. The bars shown are expressed as mean – SD from triplicate and more independent experiments. *Significant differences between each experiment and control calculated by Student’s t-test (P < 0.01). Inclusion sizes under melatonin treatment varied from 2.0 to 5.5 mm in diameter. In contrast, the majority of inclusions in the control show typical sizes, 5.0 mm in diameter, and in IFN-g treatment are atypical or small sizes, 1.5 mm in diameter (Figure 2b and c). Here, successful infections showing typical and atypical 863 Rahman et al. inclusions larger than appropriate 1 mm in diameter were counted,33,34 because chlamydial infection is thought to be able to switch from the persistent to the productive replicative cycle when the stress to sustain persistent infection is removed.11 120 Relative infection rate (%) (a) 100 80 * * * + – – + – + – + – 60 40 20 0 Melatonin IFN-γ Tryptophan (b) – – – Control – – + Melatonin – + + IFN-γ Because of homogeneity of the bacteria, the melatonin function as an anti-chlamydial agent is suggested to be associated with host cell status rather than that of chlamydia. In the case of melatonin treatment up to 0.5 mM, no growth arrest of HEp-2 cells nor obvious morphological changes (such as cell shrinkage and chromatin condensation) were observed (Figure 2b). IFN-g exposure and melatonin treatment cause atypical chlamydial inclusions, supposedly showing the typical persistent state of chlamydial infection and producing few progeny.12,33,34 The re-infection ability of progeny produced under melatonin and serotonin treatment was analysed. At 72 h post-infection, infected host cells which were treated with melatonin and serotonin had released approximately half the infectious progeny compared with untreated control (Figure 2d), while IFN-g-treated cells had released one-fourth of the control. Chlamydial progeny in inclusions under melatonin and serotonin treatment are infectious, not in the persistent state, and are released at approximately the same time as in controls. This suggests that melatonin and serotonin may affect the early establishment stage in the chlamydial development cycle, but not intracellular growth or later development. DAPI Specificity of melatonin on microbes To determine melatonin specificity for chlamydial species, infection experiments were performed with two more chlamydiae with worldwide distribution, C. trachomatis and C. felis, which cause a human sexually transmitted disease and cat conjunctivitis, respectively. Infection rates of the two chlamydiae were reduced depending on melatonin concentrations. C. felis is resistant to IFNg but susceptible to melatonin, and C. trachomatis is relatively resistant to both melatonin and IFN-g, whereas C. pneumoniae is susceptible to both IFN-g and melatonin (Figure 3). This indicates that the mechanism of melatonin to inhibit chlamydial growth is different from that of IFN-g, and the chlamydial gene Inclusion (magnified) Distribution of inclusion sizes (%) (c) IFN-γ 30 25 Melatonin 20 15 10 5 0 (d) Control 1.0 2.0 3.0 4.0 5.0 6.0 Diameter of inclusion (µm) 7.0 Relative growth of C. pneumoniae (%) 120 100 80 60 40 20 0 0 1.0 IFN-γ 50 100 200 400 50 100 200 400 Melatonin Serotonin Concentration (µM) Figure 2. Comparison of melatonin and IFN-g on chlamydial infection. (a) C. pneumoniae J138 infection of HEp-2 cells was carried out in the presence of melatonin (100 mM) and IFN-g (1.0 ng/mL) alone, with and without tryptophan (200 mM). HEp-2 cells were infected with C. pneumoniae J138 at 0.2 MOI. The rates of infected host cells were determined at 48 h post-infection by staining of chlamydial inclusions with the chlamydia specific FITC-labelled antibody and host nuclei with DAPI. Average of the actual infection rates of the controls was 15.0%. The bars shown are expressed as mean – SD from triplicate and more independent experiments. *Significant differences between each experiment and control calculated by Student’s t-test (P < 0.01). (b) Distribution of sizes of C. pneumoniae J138 inclusions. HEp-2 cell nuclei (upper row) and C. pneumoniae J138 inclusions were visualized by staining with chlamydiaspecific FITC-labelled antibody and DAPI, respectively. Inclusions are shown in the second and third rows. The areas in the boxes of the second row are magnified three times and small inclusions are marked with arrows in the third row. White bars indicate 10 mm. This figure is available in colour in the online version of this article. (c) Distributions of inclusion sizes are illustrated under melatonin treatment (thick solid line), IFN-g treatment (thin solid line) and control (dashed line). Sizes of approximately 200 inclusions were measured in each condition using the NIH image 1.63 program. (d) Growth of C. pneumoniae J138 in HEp-2 cells was tested in the presence of melatonin (50–400 mM), serotonin (50–400 mM), and IFN-g (1.0 ng/mL). Cells were infected with C. pneumoniae for 72 h, and progenies were harvested from the triplicate independent culture supernatants for each condition. The number of IFU was individually assessed with the progeny using HEp-2 cells. The data presented are the averages and the standard deviations. A colour version of Figure 2(b) is available as online Supplementary data at http://jac.oxfordjournals.org/. 864 Serotonin and melatonin inhibit chlamydial infection concentrations of 25–200 mM. Melatonin agonist and antagonist, IIK7 and luzindole, respectively, decreased chlamydial infection as effectively as melatonin, however at high concentrations luzindole exhibited toxicity to host cells. Luzindole is generally an antagonist of a melatonin receptor, but in certain cases, it has been reported to work as a melatonin agonist as well.37 Serotonin receptor agonist and antagonist, buspirone and NAN190, respectively, partially decreased chlamydial infection. Other types of serotonin receptors, which do not bind buspirone and NAN190, may be involved in the serotonin inhibition of chlamydial infection. Relative infection rate (%) 120 100 80 60 40 20 0 Melatonin IFN-γ – – + + – – – – – – + + – – – – + – – + – – – – Discussion C. pneumoniae C. felis C. trachomatis A new function of two well-known neurohormones, serotonin and melatonin, has been found, namely inhibition by 50% of chlamydial infection of human epithelial HEp-2 cells in vitro. Both effects of melatonin and serotonin are insensitive to addition of tryptophan, which is a metabolic source of serotonin and melatonin synthesis. On the other hand, inhibition of chlamydial infection by IFN-g is ceased by addition of extra tryptophan. This suggests that serotonin and melatonin are not antagonists of tryptophan for protein synthesis or any other metabolic intermediate production, and the inhibition mechanisms by melatonin and serotonin are independent of IFN-g. Pre-infection melatonin treatment of host cells, but not chlamydial cells, inhibits chlamydial infection. Post-infection treatment with melatonin also inhibited chlamydial infection, illustrating that melatonin may not block interaction between cell surfaces of chlamydiae and host cells. Moreover, addition of pertussis toxin with melatonin (but not serotonin) treatment, cancelled the melatonin effect, and ligands of melatonin and serotonin receptors also inhibit chlamydial infection, suggesting that some melatonin and serotonin receptors are involved in the inhibition of chlamydial infection. Melatonin is synthesized within the serotonin pathway from tryptophan mainly in the pineal gland and also in a number of tissues, such as retina, intestine, and so on.27 Three mammalian melatonin receptors have been identified to date. MT1 and MT2 receptors are G-protein-coupled receptors sensitive to the pertussis toxin28,29 and MT3 receptor is a quinone reductase family receptor.19 It is reported that the receptors are widely exhibited on peripheral tissues, such as brain, liver and kidney, and an epithelial cell line, HEK-293.38,39 Gene expression of MT1 and MT2 receptors was not detected by RT–PCR in our experiments using a human epithelial cell line, HEp-2 (data not shown). In addition, distribution analysis of inclusion sizes demonstrates distinctive groups of inclusions. One of these may be a group (50%) in which susceptibility to melatonin is high and prevented from visible inclusion formation. The other two groups may be resistant and partially susceptible to melatonin, resulting in typical and small inclusion formations, respectively. It may implicate the existence of a melatonin-specific receptor(s), whose signal transduction system or gene expression determines the susceptibility to melatonin relying on the host cell cycle. In contrast to melatonin, chlamydial growth inhibition by serotonin is insensitive to pertussis toxin. However, it is still highly possible that the inhibitory mechanism of serotonin involves specific receptor(s) coupled with G-proteins resistant to pertussis toxin, firstly because ligands of serotonin receptors have inhibited chlamydial infection, and secondly because most Figure 3. Host susceptibilities to different chlamydial species were tested in response to 25, 50 and 100 mM of melatonin treatment, compared with 1.0 ng/mL of IFN-g treatment. Prior to use, C. felis Fe/C-56 and C. trachomatis serovar D were adapted to HEp-2 cells by three passages and numbers of IFU were assessed in HEp-2 cells. C. pneumoniae J138, C. felis Fe/C-56 and C. trachomatis serovar D were used for infection to HEp-2 cells at 0.2 MOI. The rates of infected host cells were determined at 48 h post-infection. Each infection rate was normalized with controls to the relative infection rate, while the actual infection rates of the controls were 25.3%, 25.0% and 16.6% for C. pneumoniae, C. felis and C. trachomatis, respectively. The bars shown are expressed as means of duplicate independent experiments. repertoires, but not tryptophan synthesis, may affect susceptibility to melatonin. To examine whether or not melatonin affects only intracellular chlamydiae, eight other bacteria (E. coli, S. aureus, B. subtilis, S. pneumoniae, P. aeruginosa, H. pylori, L. pneumophila and M. tuberculosis) were tested separately at different melatonin concentrations in suitable culture medium. These bacteria represent different groups within the kingdom Eubacteria, including normal resident microflora and pathogenic bacteria, and Gram-positive and -negative bacteria. In contrast to gentamicin, to which all bacteria tested were susceptible, none of them showed effects of melatonin treatment on their growth (data not shown). Target(s) of melatonin and serotonin It has been reported that melatonin influences the hydrophobic characteristics of Escherichia coli cell surfaces.36 Therefore, chlamydial cells and host cells were pre-treated with melatonin and serotonin before infection. Chlamydial infection was reduced by pre-treatment of HEp-2 cells with melatonin and also serotonin. Pre-treatment of chlamydial cells showed no effect (Figure 4a). In addition, post-infection treatment with melatonin also reduced chlamydial infection (data not shown). This suggests that melatonin and serotonin interact with human cells but not with chlamydial cells, and some melatonin and serotonin receptors are involved in the resistance to chlamydial infection. Pertussis toxin is a well-known inhibitor of signal transduction through G protein-coupled receptors.28,29 The melatonin-mediated reduction in chlamydial infection in HEp-2 cells was abrogated by treatment with 20 ng/mL of pertussis toxin. No such inhibitory effect was observed in the case of serotonin (Figure 4b). Because of these results implying the involvement of certain G proteincoupled melatonin receptors, agonists and antagonists of melatonin and serotonin receptors were tested in chlamydial infection at 865 Rahman et al. (b) 100 80 60 * * 40 20 0 Melatonin Serotonin Treatment to 120 Relative infection rate (%) 120 Relative infection rate (%) (a) 100 80 * 60 * 40 * 20 0 – – + – – + – – Host cells + – – + Melatonin Serotonin Pertussis toxin – – – – – + + – – + – + – + – – + + Chlamydial cells Relative infection rate (%) (c) 120 Melatonin 100 IIK7 80 Luzindole 60 Serotonin 40 Buspirone 20 0 NAN190 0 25 50 100 Concentration (M) 200 Figure 4. Targets of melatonin and serotonin in HEp-2 cell infection with C. pneumoniae. (a) HEp-2 cells and C. pneumoniae J138 were separately treated with melatonin and serotonin. Treatments of chlamydial cells were performed at 200 mM for 90 min before infection in the same culture media as used for HEp-2 cells. Both C. pneumoniae J138 cells treated and mock treated were used for infection at 0.2 MOI to untreated HEp-2 cells and the actual infection rates of the controls were 6.7% [right half of panel (a)]. Treatments of host cells were performed at 200 mM for 24 h and the chemicals were washed with the culture medium before infection. Host cells treated or mock treated were infected with C. pneumoniae J138 at 0.2 MOI [left half of panel (a)]. The actual infection rates of the controls were 8.8%. (b) Effects of the pertussis toxin, an inhibitor of G-protein (20 ng/mL), were tested on the melatonin and serotonin treatments. Treatments with the pertussis toxin, melatonin and serotonin were carried out from 24 h prior to infection to 48 h post-infection. HEp-2 cells were infected with C. pneumoniae J138 at 0.2 MOI. The actual infection rate of control was 17.3%. No substantial toxicity was observed under these conditions. (c) Antagonists and agonists were applied to the C. pneumoniae J138 in vitro infection. Treatments at 25–200 mM of each chemical were carried out from 24 h prior to infection to 48 h post-infection and C. pneumoniae J138 was infected at 0.2 MOI. The actual infection rate of control was 23.0%. No substantial toxicity was observed under these conditions, except with 100 and 200 mM luzindole. In the cases of panels (a) to (c), the rates of infected host cells were determined at 48 h post-infection. The bars shown in panels (a) and (b) and points in (c) are expressed as mean – SD from triplicate and more independent experiments. *Significant differences between each experiment and control calculated by Student’s t-test (P < 0.01). physiological serotonin reactions are processed through G protein-coupled serotonin-specific receptors. In the large family of G protein-coupled serotonin receptors, Ga12/13, has already been reported to be a type of G protein insensitive to pertussis toxin.40 As common features, the melatonin and serotonin signal transductions through each specific receptor result in an increase in intracellular Ca2+ concentration and regulation of adenylyl cyclase activity, which produces adenosine 30 :50 -cyclic monophosphate (cAMP). Both concentrations of Ca2+ and cAMP produced by adenylyl cyclase subsequently modulate the gene expression status of the host cell.41,42 Interestingly, chlamydial infection is closely related to intracellular levels of cAMP and Ca2+. Chlamydial infection is inhibited by high concentrations of extracellular cAMP or cGMP.43 Ca2+ mobility across the cell membrane is rapidly increased by chlamydial attachment on to host cells.43 Recently, Ca2+-dependent phagolysosomal fusion has been illustrated as a host defence strategy, in which host cells may monitor Ca2+ influx accompanying injuries by invasion of intracellular pathogens and the Ca2+ influx may promote lysosomal exocytosis.44 The type III secretion system is a candidate to injure the host cell membrane and is thought to be an invasion system of chlamydiae as well. It is likely that melatonin modulates intracellular Ca2+ and/or cAMP levels and leads to increased phagolysosomal activity of the host cells against the chlamydial infection. To detect modification of gene expression in HEp-2 cells by the melatonin and serotonin treatments, microarray analysis was carried out with AceGene oligo-DNA chips (HitachiSoft, Japan) arraying approximately ten thousand known human genes. Positively and negatively regulated genes in the treated cells were nominated when the signals are more than 1.5 times or less than 0.6 times compared with untreated cells, respectively. Melatonin and serotonin treatments resulted in modifying mRNA accumulation moderately, except for some genes, e.g. myosin and kinesin family members and platelet-derived growth factors, implying that the melatonin and serotonin treatments may remodel the intracellular matrix to a state inadequate for chlamydial intracellular growth (data not shown). Melatonin, originally identified as an effector for circadian rhythms,23 is now known as a hormone involved in a vast range of homeostasis maintenance, i.e. human mood,24 seasonal timing,45 866 Serotonin and melatonin inhibit chlamydial infection anti-ageing,46 oncostatic,47 antioxidant defence,27,48 and immunomodulation.49 Many of the melatonin effects seem to relate to mitochondrial functions as a direct scavenger for a variety of toxic oxygen and nitrogen-based reactants.50 Enhancements of the nonspecific and specific immune responses by exogenous melatonin are reported as stimulation for proliferation/production of NK cells and monocytes,51 and prevention from the reduction in B- and Tcell proliferation and Th1 cytokine secretions caused by retroviral infection.52 Despite the many investigations on melatonin and serotonin that have been carried out, they have not been generally recognized as antimicrobial agents, apart from a few reports for melatonin showing protective effects in mice against viral infections by Venezuelan equine encephalomyelitis virus,53 Semliki Forest virus54 and West Nile virus,55 and against infection by a helminth parasite, Schistosoma mansoni.56 Melatonin is used in Mycobacterium tuberculosis infection to increase the chemotherapeutic activity of isoniazid (pyridine-4-carbohydrazide), which is an inhibitor of tryptophan metabolism to NAD.57 So, the inhibition of infection for the intracellular bacterium chlamydia is a new function of melatonin and serotonin. Variation of tryptophan biosynthesis genes is the most remarkable feature in the comparison of gene repertoires among chlamydial genomes. Considering that tryptophan depletion by IFN-g is a crucial host defence mechanism against chlamydiae, it evokes all evolutional relationships between strategies of host defence and parasite infection, leading to chlamydial host and tissue tropisms. There is an apparent paradox: C. trachomatis and C. pneumoniae, worldwide common parasites to human but not to animals, contain partial and no tryptophan biosynthesis genes in their genomes, respectively, and they are susceptible to both IFN-g and melatonin; on the other hand, C. felis, a worldwide common parasite of cats but rarely humans, seems to possess an almost complete gene set for tryptophan biosynthesis (Y. Azuma, H. Hirakawa, A. Yamashita, Y. Cai, M. A. Rahman, H. Suzuki, S. Mitaku, H. Toh, T. Murakami, K. Sugi, H. Hayashi, H. Fukushi, M. Hattori, S. Kuhara and M. Shirai, unpublished data) and show marked resistance to IFN-g, but not to melatonin. Whereas, tryptophan might play a key role for the molecular basis of host–parasite interaction in terms of IFN-g resistance, the gene repertoires of tryptophan biosynthesis cannot entirely explain host tropisms of individual chlamydial species. The different susceptibilities to melatonin or serotonin, or any tryptophan derivatives may be involved in determination of the host tropisms. Different melatonin susceptibilities among chlamydial species may depend on gene variations of the chlamydial species responsible for host defence systems through melatonin receptors. Genes discriminating C. trachomatis from C. felis and C. pneumoniae consist of 69 genes, whose products are annotated as 60 unknown proteins, a membrane thiol protease, two transporters, two polymorphic outer membrane proteins and four phospholipase D superfamily proteins. Although our studies have provided evidence of possible anti-chlamydial functions of melatonin and serotonin, further investigations are still needed to determine the chlamydial factors that are involved in the various diseases and host/tissue tropism. Acknowledgements This study was supported by the Japan Society for the Promotion of Science, Research for the Future Program (JSPS-RETF 00L01411), a Grant-in-Aid for Scientific Research from the Ministry of Education, Culture, Sports, Science and Technology of Japan (KAKENHI: 14013043, 15019069, 15659172, 16012245, 16790255), the New Frontier Project of Yamaguchi University School of Medicine, and the Venture Business Laboratory, Yamaguchi University. Supplementary data Figure 2(b) is available in colour as online supplementary data at http://jac.oxfordjournals.org/. References 1. Moulder JW. Interaction of chlamydiae and host cells in vitro. Microbiol Rev 1991; 55: 143–90. 2. Rottenberg ME, Gigliotti-Rothfuchs A, Wigzell H. The role of IFN-gamma in the outcome of chlamydial infection. Curr Opin Immunol 2002; 14: 444–51. 3. Rosenfeld ME, Blessing E, Lin TM et al. Chlamydia, inflammation, and atherogenesis. J Infect Dis 2000; 181 Suppl 3: S492–7. 4. Hahn DL, Dodge RW, Golubjatnikov R. Association of Chlamydia pneumoniae (strain TWAR) infection with wheezing, asthmatic bronchitis, and adult-onset asthma. JAMA 1991; 266: 225–30. 5. Robinson SR, Dobson C, Lyons J. Challenges and directions for the pathogen hypothesis of Alzheimer’s disease. Neurobiol Aging 2004; 25: 629–37. 6. Itzhaki RF, Wozniak MA, Appelt DM et al. Infiltration of the brain by pathogens causes Alzheimer’s disease. Neurobiol Aging 2004; 25: 619–27. 7. Baker KA. A virus from a pneumonia of cats and its possible relation to the cause of a typical pneumonia in man. Science 1942; 96: 475–6. 8. Pantoja LG, Miller RD, Ramirez JA et al. Inhibition of Chlamydia pneumoniae replication in human aortic smooth muscle cells by gamma interferon-induced indoleamine 2,3-dioxygenase activity. Infect Immun 2000; 68: 6478–81. 9. Mehta SJ, Miller RD, Ramirez JA et al. Inhibition of Chlamydia pneumoniae replication in HEp-2 cells by interferon-gamma: role of tryptophan catabolism. J Infect Dis 1998; 177: 1326–31. 10. Moffett JR, Namboodiri MA. Tryptophan and the immune response. Immunol Cell Biol 2003; 81: 247–65. 11. Beatty WL, Byrne GI, Morrison RP. Morphologic and antigenic characterization of interferon gamma-mediated persistent Chlamydia trachomatis infection in vitro. Proc Natl Acad Sci USA 1993; 90: 3998–4002. 12. Belland RJ, Nelson DE, Virok D et al. Transcriptome analysis of chlamydial growth during IFN-gamma-mediated persistence and reactivation. Proc Natl Acad Sci USA 2003; 100: 15971–6. 13. Kalman SMW, Marathe R, Lammel C et al. Comparative genomes of Chlamydia pneumoniae and C. trachomatis. Nat Genet 1999; 21: 385–9. 14. Xie GBC, Bonner CA, Jensen RA. Dynamic diversity of the tryptophan pathway in chlamydiae: reductive evolution and a novel operon for tryptophan recapture. Genome Biol 2002; 3: 0051.1–17. 15. Schachter J. Chlamydial infections (first of three parts). New Engl J Med 1978; 298: 428–35. 16. Shields PJ, Eccleston D. Evidence for the synthesis and storage of 5-hydroxytryptamine in two separate pools in the brain. J Neurochem 1973; 20: 881–8. 17. Marmaras VJ, Mimikos N. Enzymic formation of serotonin in mammalian blood platelets and red cells. Experientia 1971; 27: 196–7. 18. Sjolund K, Sanden G, Hakanson R et al. Endocrine cells in human intestine: an immunocytochemical study. Gastroenterology 1983; 85: 1120–30. 867 Rahman et al. 19. Abrahamsson PA, Wadstrom LB, Alumets J et al. Peptide-hormoneand serotonin-immunoreactive cells in normal and hyperplastic prostate glands. Pathol Res Pract 1986; 181: 675–83. 20. Seuwen K, Pouyssegur J. Serotonin as a growth factor. Biochem Pharmacol 1990; 39: 985–90. 21. Buznikov GA, Lambert HW, Lauder JM. Serotonin and serotoninlike substances as regulators of early embryogenesis and morphogenesis. Cell Tissue Res 2001; 305: 177–86. 22. Moiseiwitsch JR, Raymond JR, Tamir H et al. Regulation by serotonin of tooth-germ morphogenesis and gene expression in mouse mandibular explant cultures. Arch Oral Biol 1998; 43: 789–800. 23. Binkley SA. Circadian rhythms of pineal function in rats. Endocr Rev 1983; 4: 255–70. 24. Lieberman HR, Waldhauser F, Garfield G et al. Effects of melatonin on human mood and performance. Brain Res 1984; 323: 201–7. 25. Krause DN, Dubocovich ML. Regulatory sites in the melatonin system of mammals. Trends Neurosci 1990; 13: 464–70. 26. Reiter RJ. The mammalian pineal gland: structure and function. Am J Anat 1981; 162: 287–313. 27. Tan DX, Manchester LC, Hardeland R et al. Melatonin: a hormone, a tissue factor, an autocoid, a paracoid, and an antioxidant vitamin. J Pineal Res 2003; 34: 75–8. 28. Carlson LL, Weaver DR, Reppert SM. Melatonin signal transduction in hamster brain: inhibition of adenylyl cyclase by a pertussis toxinsensitive G protein. Endocrinology 1989; 125: 2670–6. 29. Chan AS, Lai FP, Lo RK et al. Melatonin mt1 and MT2 receptors stimulate c-Jun N-terminal kinase via pertussis toxin-sensitive and -insensitive G proteins. Cell Signal 2002; 14: 249–57. 30. Shirai MHH, Kimoto M, Tabuchi M et al. Comparison of whole genome sequences of Chlamydia pneumoniae J138 from Japan and CWL029 from USA. Nucleic Acids Res 2000; 28: 2311–4. 31. Cai Y, Fukushi H, Koyasu S et al. An etiological investigation of domestic cats with conjunctivitis and upper respiratory tract disease in Japan. J Vet Med Sci 2002; 64: 215–9. 32. Matsushima H, Shirai M, Ouchi K et al. Lymphotoxin inhibits Chlamydia pneumoniae growth in HEp-2 cells. Infect Immun 1999; 67: 3175–9. 33. Pantoja LG, Miller RD, Ramirez JA et al. Characterization of Chlamydia pneumoniae persistence in HEp-2 cells treated with gamma interferon. Infect Immun 2001; 69: 7927–32. 34. Byrne GI, Ouellette SP, Wang Z et al. Chlamydia pneumoniae expresses genes required for DNA replication but not cytokinesis during persistent infection of HEp-2 cells. Infect Immun 2001; 69: 5423–9. 35. Valero N, Bonilla E, Pons H et al. Melatonin induces changes to serum cytokines in mice infected with the Venezuelan equine encephalomyelitis virus. Trans R Soc Trop Med Hyg 2002; 96: 348–51. 36. Uberos J, Augustin C, Liebana J et al. Comparative study of the influence of melatonin and vitamin E on the surface characteristics of Escherichia coli. Lett Appl Microbiol 2001; 32: 303–6. 37. Cabrera J, Quintana J, Reiter RJ et al. Melatonin prevents apoptosis and enhances HSP27 mRNA expression induced by heat shock in HL-60 cells: possible involvement of the MT2 receptor. J Pineal Res 2003; 35: 231–8. 38. Chan CW, Song Y, Ailenberg M et al. Studies of melatonin effects on epithelia using the human embryonic kidney-293(HEK-293) cell line. Endocrinology 1997; 138: 4732–9. 39. Naji L, Carrillo-Vico A, Guerrero JM et al. Expression of membrane and nuclear melatonin receptors in mouse peripheral organs. Life Sci 2004; 74: 2227–36. 40. Berg KA, Maayani S, Goldfarb J et al. Effector pathway-dependent relative efficacy at serotonin type 2A and 2C receptors: evidence for agonist-directed trafficking of receptor stimulus. Mol Pharmacol 1998; 54: 94–104. 41. Reppert SMGC, Mahle CD, Weaver DR et al. Molecular characterization of a second melatonin receptor expressed in human retina and brain: the Mel1b melatonin receptor. Proc Natl Acad Sci USA 1995; 92: 8734–8. 42. De Vivo M, Maayani S. Stimulation and inhibition of adenylyl cyclase by distinct 5-hydroxytryptamine receptors. Biochem Pharmacol 1990; 40: 1551–8. 43. Ward ME, Salari H. Control mechanisms governing the infectivity of Chlamydia trachomatis for hela cells: modulation by cyclic nucleotides, prostaglandins and calcium. J Gen Microbiol 1982; 128: 639–50. 44. Roy D, Liston DR, Idone VJ et al. A process for controlling intracellular bacterial infections induced by membrane injury. Science 2004; 304: 1515–8. 45. Pevet P. Melatonin: from seasonal to circadian signal. J Neuroendocrinol 2003; 15: 422–6. 46. Karasek M, Reiter RJ. Melatonin and aging. Neuroendocrinol Lett 2002; 23 Suppl 1: 14–6. 47. Sanchez-Barcelo EJ, Cos S, Fernandez R et al. Melatonin and mammary cancer: a short review. Endocr Relat Cancer 2003; 10: 153–9. 48. Reiter RJ. Functional aspects of the pineal hormone melatonin in combating cell and tissue damage induced by free radicals. Eur J Endocrinol 1996; 134: 412–20. 49. Zhang Z, Araghi-Niknam M, Liang B et al. Prevention of immune dysfunction and vitamin E loss by dehydroepiandrosterone and melatonin supplementation during murine retrovirus infection. Immunology 1999; 96: 291–7. 50. Leon J, Acuna-Castroviejo D, Sainz RM et al. Melatonin and mitochondrial function. Life Sci 2004; 75: 765–90. 51. Currier NL, Sun LZ, Miller SC. Exogenous melatonin: quantitative enhancement in vivo of cells mediating non-specific immunity. J Neuroimmunol 2000; 104: 101–8. 52. Bonilla E, Valero N, Chacin-Bonilla L et al. Melatonin and viral infections. J Pineal Res 2004; 36: 73–9. 53. Bonilla E, Valero-Fuenmayor N, Pons H et al. Melatonin protects mice infected with Venezuelan equine encephalomyelitis virus. Cell Mol Life Sci 1997; 53: 430–4. 54. Maestroni GJ, Conti A, Pierpaoli W. Pineal melatonin, its fundamental immunoregulatory role in aging and cancer. Ann NY Acad Sci 1988; 521: 140–8. 55. Ben-Nathan D, Maestroni GJ, Lustig S et al. Protective effects of melatonin in mice infected with encephalitis viruses. Arch Virol 1995; 140: 223–30. 56. El-Sokkary GH, Omar HM, Hassanein AF et al. Melatonin reduces oxidative damage and increases survival of mice infected with Schistosoma mansoni. Free Radic Biol Med 2002; 32: 319–32. 57. Wiid I, Hoal-van Helden E, Hon D et al. Potentiation of isoniazid activity against Mycobacterium tuberculosis by melatonin. Antimicrob Agents Chemother 1999; 43: 975–7. 868