Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
Microbial metabolism wikipedia , lookup
Mitochondrion wikipedia , lookup
Mitochondrial replacement therapy wikipedia , lookup
Metabolic network modelling wikipedia , lookup
Metabolomics wikipedia , lookup
Citric acid cycle wikipedia , lookup
Metalloprotein wikipedia , lookup
Isotopic labeling wikipedia , lookup
Evolution of metal ions in biological systems wikipedia , lookup
Peptide synthesis wikipedia , lookup
Pharmacometabolomics wikipedia , lookup
Nitrogen cycle wikipedia , lookup
Wilson's disease wikipedia , lookup
Biochemistry wikipedia , lookup
Gaseous signaling molecules wikipedia , lookup
Oligonucleotide synthesis wikipedia , lookup
Glyceroneogenesis wikipedia , lookup
Artificial gene synthesis wikipedia , lookup
Biosynthesis wikipedia , lookup
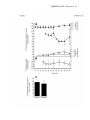
AJP-Endo Articles in PresS. Published on August 13, 2002 as DOI 10.1152/ajpendo.00246.2002 MS#E00246-2002, Nissim et al 1 Regulation of Urea Synthesis by Agmatine in the Perfused Liver: Studies With 15N Itzhak Nissim*, Oksana Horyn, Yevgeny Daikhin, Ilana Nissim, Adam Lazarow and Marc Yudkoff Children Hospital of Philadelphia, Division of Child Development and Rehabilitation, Department of Pediatrics, University of Pennsylvania School of Medicine, Philadelphia, Pennsylvania 19104 Running Title: Regulation of Hepatic Ureagenesis CONTENTS: Pages: 40 Figures: 8 Tables: 2 References: 49 * Correspondence should be addressed to: Itzhak Nissim, Ph.D. Division of Child Development Abramson Pediatrics Research Ctr. Room-510C 34th Street and Civic Center Boulevard Philadelphia, PA 19104-4318 FAX: (215)-590-5199 e-mail : [email protected] Copyright 2002 by the American Physiological Society. MS#E00246-2002, Nissim et al 2 ABSTRACT Administration of arginine or a high protein diet increases the hepatic content of Nacetylglutamate (NAG) and the synthesis of urea. However, the underlying mechanism is unknown. We have explored the hypothesis that agmatine, a metabolite of arginine, may stimulate NAG synthesis, and, thereby, urea synthesis. We tested this hypothesis in a liver perfusion system to determine: (i) The metabolism of L-[guanidino-15N2]arginine to either agmatine, nitric-oxide (NO) and/or urea; (ii) Hepatic uptake of perfusate agmatine and its action on hepatic nitrogen metabolism; and (iii) The role of arginine, agmatine or NO in regulating NAG synthesis and ureagenesis in livers perfused with 15N-labeled glutamine and unlabeled ammonia or 15NH4Cl and unlabeled glutamine. Our principal findings are: (i) [guanidino-15N2]agmatine is formed in the liver from perfusate L-[guanidino-15N2]arginine. About 90% of hepatic agmatine is derived from perfusate arginine; (ii) Perfusions with agmatine significantly stimulated the synthesis of [15N]NAG and [15N]urea from 15 N-labeled ammonia or glutamine; and (iii) The increased levels of hepatic agmatine are strongly correlated with increased levels and synthesis of 15N-labeled NAG and [15N]urea. These data suggest a possible therapeutic strategy encompassing the use of agmatine for the treatment of disturbed ureagenesis, whether secondary to inborn errors of metabolism or to liver disease. Key Words: Arginine, N-acetylglutamate, Carbamoyl Phosphate Synthetase-I, Hyperammonemia, Nitric oxide MS#E00246-2002, Nissim et al 3 INTRODUCTION The regulation of ureagenesis has posed a complex problem since the first description of the urea cycle by Krebs and Henseleit in 1932 (20). The first and most critical step in urea synthesis is the conversion of NH4+ and HCO3 - into carbamoyl phosphate by mitochondrial carbamoyl phosphate synthetase I (CPS-I), ( EC 6.3.5.5) (8, 13, 16, 25), which requires N-acetylglutamate (NAG) as an obligatory effector for its activation (8, 13, 14). Brosnan (4) has suggested that acute increases in ureagenesis are initiated by greater availability of NAG. Increased NAG synthesis and concomitant ureagenesis is observed after intake of arginine or a high protein diet (7, 19, 27, 44). However, it is not clear how and why arginine or a high protein diet increases the hepatic NAG level, and, thereby, urea synthesis. It is unknown whether arginine directly or one of its metabolites stimulates the synthesis of NAG, and, thereby, ureagenesis. As illustrated in Figure 1, L-arginine can be metabolized in the liver via several pathways: (i) Hydrolysis via arginase to form urea, the end product of the urea cycle. There are two isoenzymes of mammalian arginase (8, 17, 38, 47). Arginase I is mainly present in the cytosol of periportal hepatocytes and is linked to urea cycle activity (2, 25, 38, 47). Arginase II is found primarily in mitochondria (7, 17, 38, 47); (ii) Decarboxylation via mitochondrial arginine decarboxylase (ADC) to produce agmatine and CO2 (14, 39, 47); and/or (iii) Metabolism via NOsynthase to form nitric oxide (NO) and citrulline (21, 47). The ADC and NO-synthase reactions are quantitatively minor routes of overall arginine metabolism, but the products of these pathways (NO and agmatine) have significant roles as signaling molecules that regulate many metabolic and physiologic functions (21, 40, 41, 47). However, their role in the regulation of ureagenesis is uncertain. Understanding this relationship may provide a new therapeutic strategy for treatment of urea cycle disorders and hyperammonemia . It has been demonstrated that the ADC pathway is present in rat kidney, brain, liver and gut (40, 41). Recent studies have highlighted the importance of agmatine for a number of physiological MS#E00246-2002, Nissim et al 4 processes, including: dose-dependent release of insulin from islet cells exposed to glucose, proliferation of vascular smooth muscle, regulation of intracellular polyamine levels and cellular proliferation, regulation of neurotransmitter receptors, modulation of opioid analgesics, and inhibition of NO synthesis (2, 40, 41). Agmatine may stimulate the release of catecholamines from adrenal chromaffin cells by binding to _ 2 -adrenergic receptors (14). Thus, agmatine, which is widely distributed in mammalian tissue (37, 39), may have a role as a hormone and/or polyamine precursor for a multiple metabolic functions. However, the significance of agmatine as a signaling molecule in regulating the urea cycle remains to be defined. In the current investigation we have explored the hypothesis that dietary arginine (whether free or as a part of a high protein diet), is decarboxylated to agmatine in hepatic mitochondria, and that agmatine, not arginine, is the regulator of NAG synthesis and the resulting activation of mitochondrial CPS-I. As illustrated in Figure 1, both agmatine and NAG are synthesized in mitochondria, the latter from acetyl-CoA and glutamate by N-acetylglutamate synthetase (25). The mitochondrial location of both ADC and NAG synthetase makes agmatine a reasonable candidate for regulation of NAG synthesis, and, thereby, activation of CPS-I. We have tested the above hypothesis in three steps: (i) Initial studies that involve perfusions with L-[guanidino-15N2]arginine in order to determine the hepatic uptake of arginine and its relative metabolism to either agmatine, NO and/or urea, as indicated in Figure 1; (ii) Perfusions with agmatine to determine its hepatic uptake and the dose dependence of agmatine action on urea synthesis; and (iii) Studies to elucidate the role of arginine, agmatine or NO in the regulation of NAG synthesis and ureagenesis in liver perfused with either [2-15N]- or [5-15N]-labeled glutamine and unlabeled ammonia or 15NH4Cl and unlabeled glutamine, as we have previously described (3, 5, 31, 32). The results obtained substantiate the hypothesis that agmatine regulates NAG and urea synthesis. MS#E00246-2002, Nissim et al 5 Materials and Methods I. Liver Perfusions: Livers from fed, male Sprague Dawley rats were perfused "in situ" in the non-recirculating mode as described by Pastor et. al (35). We employed the single-pass perfusion with antegrade flow because this model preserves the normal lobular microcirculation of the liver and physiologic flow direction. The model also avoids the problem of substrate recycling from perivenous to periportal hepatocytes. Oxygen consumption was measured continuously using a Clark electrode and the Oxygen Measuring System (Instech SYS203, Plymouth Meeting, PA). One electrode was attached into the inflow cannula and one into the outflow. This system was linked to a computer equipped with the WinDaq/Lite/Pro/Pro+ Data System (DATAQ Instrument, Akron, Ohio) for online acquisition and recording of signals corresponding to inflow and outflow levels of pO2. The basic perfusion medium was Krebs saline continuously gassed with 95% O2 / 5% CO2 and containing lactate (2.1 mM) and pyruvate (0.3 mM) as metabolic fuels. Perfusion flow rate (3-3.5 ml/min/g), hepatic temperature, pH and pO2 (in influent and effluent media) were monitored throughout, and oxygen consumption was calculated. After 15 min. of pre-perfusion with the basic perfusate, we changed to a medium that contained, in addition to the lactate and pyruvate, precursors for urea nitrogen or modulators of urea synthesis as indicated below. II. Experimental Design: To establish the role of arginine or its metabolites, NO or agmatine, in the regulation of NAG and urea synthesis, we first performed experiments to determine the rate of hepatic uptake of arginine and its relative metabolism to agmatine (ADC reaction), urea (arginase reaction) or nitric oxide (NO-synthase reaction). To this end, after 15 minutes pre-perfusion with a basic perfusate as indicated above, we changed to a perfusate that contained 0.3 mM NH4Cl, 1mM unlabeled glutamine and 0.5mM L-[guanidino-15N2]arginine ((99 mole % excess, (MPE)). Separate perfusate reservoirs, each containing different media, were used to facilitate changes in perfusate medium. Perfusion was continued for a total of 70 min. Samples were taken from the influent and MS#E00246-2002, Nissim et al 6 effluent media for chemical and GC-MS analyses. At the end of the perfusions, livers were freezeclamped, and used for measurements of metabolite concentrations and 15N enrichment in arginine and agmatine. The next series of experiments was designed to examine the effect of exogenous agmatine or nitroprusside (SNP), a donor of NO, on total urea synthesis. We also evaluated a possible dosedependent action of either arginine, agmatine or NO on urea synthesis from ammonia and glutamine. Perfusions were performed with: (i) agmatine at an increasing concentrations (µM) of 25, 50, 100, 250, 500 or 1000; (ii) SNP, at an increasing concentration (µM) of 25, 50 or 100 or (iii) arginine at an increasing concentration (µM) of 250, 500 or 1000. The dose dependence was studied in the same liver perfusion. At the indicated times, a separate perfusate reservoir containing different media was used to facilitate changes in the concentration of each modulator. To examine the role of arginine or its metabolites in the regulation of N-acetylglutamate synthesis and ureagenesis under conditions and concentrations that approximate those that exist in vivo, perfusions were performed without (control) or with addition of 100 µM agmatine, 25 µM SNP or 500 µS arginine. The 15N-labeled precursor for urea nitrogen was either: (a) glutamine (1mM), [5-15N]glutamine or [2-15N]glutamine (99 MPE) and NH4Cl (0.3mM), or (b) unlabeled glutamine (1mM) and 15 NH 4Cl (0.3mM), (99 MPE). Pre-perfusions with basic medium were performed for 15 minutes, and continued for 70 minutes with the indicated 15 N precursor or modulator. III. Chemical Analyses: Samples were taken at the indicated times from influent and effluent media for chemical determination of metabolites. At the end of the perfusion (70 min), livers were freeze-clamped. The frozen livers were extracted into perchloric acid and the neutralized extracts were used for chemical analysis. Adenine nucleotides (ATP, ADP and AMP) were determined using 31P-NMR analysis as described (30). Amino acids as well as agmatine were measured by MS#E00246-2002, Nissim et al 7 HPLC, using precolumn derivatization with o-phthalaldehyde (18). Measurements also were made of glucose (1), urea and ammonia (29), lactate (15) and pyruvate (12). IV. GC-MS Methodology: GC-MS measurements of hepatic NAG levels and 15 N isotopic enrichment in various metabolites were performed on a Hewlett Packard 5970 MSD and/or 5971 MSD coupled with a 5890 HP-GC, as described previously (3, 5, 31, 32). The following measurements were performed: i. ) Amino Acids, ammonia and urea: For measurement of 15N enrichment in urea and amino acids, samples were prepared as we have previously described (3, 5, 31, 32). Briefly, a 500 µl aliquot of effluent or liver extract was purified via an AG-50 column (H+; 100-200 mesh; 0.5 x 2.5 cm), and then converted into the t-butyldimethylsilyl (t-BDMS) derivatives. The m/z 231, 232, 233, 234 of the urea t-BDMS derivative (46) were monitored for singly (Um+1) and doubly (Um+2) labeled urea determination (3, 31, 32). Isotopic enrichment in glutamate, glutamine, aspartate and alanine was monitored using ratios of ions at m/z of 433/432, 432/431, 419/418 and 261/260, respectively. 15 NH3 enrichment was measured after conversion of ammonia to norvaline as described (5). 15N enrichment in L-[guanidino-15N2]arginine was determined after derivatization with 100 µl of tri-fluoroacetic anhydride (TFAA) and monitoring the ratio of ions at m/z 377/375 as we have previously described (28). ii.) Measurement of NAG Concentration: The level of NAG in freeze-clamped livers was determined using GC-MS and a modification of the conventional isotope dilution technique, as described (31). First, 15N-labeled NAG was synthezised (99 MPE) by reacting [15N]glutamate (99 MPE) with acetic anhydride. The 15N-labeled NAG was used to prepare standard dilution curves by mixing labeled and unlabeled NAG, and spiking samples for determination of NAG levels by isotope dilution. MS#E00246-2002, Nissim et al 8 NAG in standard solutions or samples was converted into the methyl esters as follows: Samples were dried under N2 and azeotroped with methylene chloride. Then 100 µl of methanolic HCl (3N) (Supelco) was added. Capped vials were heated at 60oC for 10 min., cooled and dried under a gentle stream of N2. The residue was extracted into 1 ml of ethyl acetate and 300 µl of H2O after vortexing for 30 sec., the organic layer was removed, dried and reconstituted in 75 µl of ethyl acetate. Usually 1-2 µl were injected into the GC-MS, and isotopic enrichment in NAG was determined using the ratio at m/z 159/158. To determine the concentration of NAG in freeze-clamped liver extracts, an aliquot (500 µl) was assayed as indicated above for 15N enrichment (I1), after 15N labeled precursor was used in the perfusion, e.g., in experiments with 15NH4Cl or [2-15N]glutamine (in this case, the value of I1 was between 20 - 30 MPE). A second aliquot (500 µl) was spiked with 5 nmoles of unlabeled NAG, and the second isotopic enrichment (I2) was determined. In experiments with [5-15N]glutamine, I1 usually was below 3 MPE. Therefore, the second aliquot of 500 µl was spiked with 5 nmoles of 15N labeled NAG and I2 was determined. NAG concentrations were calculated using the isotope dilution technique (31, 48). With each series of measurements, a calibration curve of NAG with a known isotopic enrichment (1-50 MPE) was prepared and analyzed by GC-MS. In nearly every preparation, we achieved an excellent agreement between the observed and the expected 15 N enrichment in NAG with r values around 0.9. iii.) Determination of 15 N-labeled Agmatine: We first synthesized [guanidino- 15 N2]agmatine as described (6). Briefly, 18mM L-[guanidino-15N2]arginine in 0.2M sodium acetate, pH 5.2, containing 5mM pyridoxal phosphate (Sigma), 0.1% BSA and 1 I.U., bacterial ADC (Sigma # A8134), was incubated at 37oC for 2 hour. Then, an additional 1 IU of ADC was added and incubation was continued for another 3 hours. The reaction was stopped by addition of 5 M KOH. Agmatine was extracted with n-butanol and dried under vacuum. Purity was determined by HPLC, MS#E00246-2002, Nissim et al 9 and yield was almost 100%. The 15 N-labeled agmatine was used to develop the GC-MS methodology for measurement of 15N-labeled agmatine in biological samples and for preparation of standard isotope dilution curves by mixing labeled and unlabeled agmatine. For GC-MS analysis of method for determination of 15 15 N-labeled agmatine we have applied a previously described N-labeled arginine in biological samples (28). To examine the accuracy of this method for measurement of 15N enrichment in agmatine a standard dilution curve of 15 N-labeled agmatine, with a known isotopic enrichment (1-50 MPE) was prepared. Samples were alkalinized with 1ml of 3 M NH4OH and then loaded into a column containing the acetate form of Dowex 1-X8 (100-200 mesh, Bio-Rad). The column was washed with 3-4 ml H2O, and agmatine was eluted with 3ml of 2N HCl. The eluate was dried under N2, azeotroped with CH2Cl2 and derivatized with 100 µl of TFAA at 100oC for 10 minutes. Usually, 2-4 µl was injected into GC-MS for analysis. Derivatized agmatine was separated from arginine and other compounds on a capillary column (15m x 0.25 I.D., ZB-1, Phenomenex # 023545). GC conditions were: injector temperature 250oC and temperature program 110 oC isothermal for 2 minutes and then 10oC/min. We have found that approximately 90% of agmatine was converted into N-tri-TFA-agmatine with a major ion at m/z 349, and about10% was converted into N-tetra-TFA with a major ion at m/z 445. We used the N-tri-TFA-agmatine and m/z 351/349 ratio to determine isotopic enrichment in [guanidino15 N2]agmatine. When the standard isotope dilution curve was analyzed, we achieved excellent agreement between the observed and the expected 15N enrichment in agmatine with r values of 0.9 or better. For determination of 15N-labeled agmatine following liver perfusion with L-[guanidino15 N2]arginine, a 1ml of perfusate or liver extract was alkalinized with 1ml of 3 M NH4OH and then loaded onto a Dowex 1-X8 column (acetate). Separation, derivatization and GC-MS analysis were completed as indicated above. MS#E00246-2002, Nissim et al 10 V. Determination of NO released in effluent: NO released (NO-2+NO-3) in the effluent during liver perfusions were determined with a Nitric Oxide Analyzer (Sievers Instruments, Boulder, CO), as indicated (36). Briefly, perfusate samples (10 µl) were injected in a reaction chamber containing a VCl3/HCl mixture heated at 90oC, and nitric oxide was detected by chemiluminescence. With each measurement, a standard curve analysis of known concentration of NaNO3 was injected and used for calculation of NO-2+NO-3 in the perfusate samples. VI. Statistical analyses: Statistical and regression analyses were carried out using In-STAT 1.14 software for the Macintosh. We used the Student-t test or ANOVA test to compare two groups or differences among groups as needed. A p value less than 0.05 was taken as indicating a statistically significant difference. VII. Materials and Animals : Male Sprague-Dawley rats (Charles-River) were fed ad lib on a standard rat chow diet. Chemicals were of analytical grade and obtained from Sigma-Aldrich. Enzymes and cofactors for the analysis of urea, lactate, pyruvate, glucose and ammonia were obtained from Sigma. 15N-labeled arginine, glutamine and ammonia (99 mole % excess) were from Isotec. MS#E00246-2002, Nissim et al 11 Results Hepatic uptake and metabolism of arginine - Production of agmatine: The initial series of experiments was designed to determine the extent to which perfusate arginine is taken up and metabolized to either urea, agmatine or nitric oxide. Figure 2 demonstrates the formation of these metabolites and oxygen consumption during perfusion with L-[guanidino-15N2]arginine. The constancy of oxygen consumption is an indication of the viability and stability of the perfused livers. The rate of L-[guanidino - 15N2]arginine uptake is about 120-140 nmoles/min/g liver (Figure 2B). At the end of 70 minutes perfusion, the isotopic enrichment of hepatic arginine was 46.3 ± 9.3 MPE (Figure 2C), indicating that approximately 50% of hepatic arginine was derived from arginine entering the liver via the portal vein . Arginine in the effluent was about 0.45 mM and that in the perfusate 0.5 mM, a finding in agreement with a previous estimate that approximately 15% of perfusate arginine is metabolized in the liver (33, 34). The major metabolites of L-[guanidino-15 N 2 ]arginine are [15N2]urea and [guanidino-15N2]agmatine (Figure 2A). The 15N enrichment in doubly labeled urea was about 10-13 MPE between 20-40 minutes and labeling decreased to 5-6 MPE by 70 minutes. The decreased enrichment in [15N2]urea after 40 minutes of perfusion reflects increased formation of unlabeled urea from ammonia and glutamine via the urea cycle. The total (labeled and unlabeled) urea output between 20-70 minutes was approximately 1000 nmoles/min/g liver. Total [15N2]urea output was about 70-90 nmoles/min/g (Figure 2A), which is about 80% of arginine uptake. Therefore, when physiologic concentrations of glutamine and ammonia are provided, approximately 7-9% of total urea in the effluent was formed from external arginine entering the liver via the portal vein. Agmatine is the second major metabolite of arginine. The output of [guanidino15 N2]agmatine in the effluent was about 14-20 nmoles/min/g (Figure 2A), indicating that approximately 10-15% of arginine uptake was recovered as agmatine in the effluent. The MS#E00246-2002, Nissim et al 12 concentration of agmatine in the liver extract was about 130 nmoles/g, and 41.1 ± 11.2 % of this was in the form of [guanidino-15N2]agmatine (Figure 2C). The ratio between [guanidino15 N2]agmatine and L-[guanidino-15N 2]arginine was 0.87, indicating that approximately 87% of hepatic agmatine was derived from external arginine. An additional pathway of arginine metabolism is via nitric oxide synthetase (21, 46). The output of NO, measured as NO 2 + NO -3 was 1.5-2 nmoles/min/g, which is about 1% of arginine uptake (Figure 2A). The current value of NO output is in good agreement with a previous study showing that, in livers perfused with 1mM arginine, the sum of NO -2 + NO 3 output was about 4 nmoles/min/g dry weight (35). Dose dependence of agmatine action on urea synthesis: Following the demonstration that agmatine is formed in the liver from external arginine, we next examined the hepatic uptake of perfusate agmatine and its action on hepatic nitrogen metabolism and ureagenesis. We also evaluated the effect of perfusate agmatine on these parameters. Figure 3 depicts the results of perfusions with unlabeled glutamine (1mM), ammonia (0.3mM), and either 25, 50 or 100 µM agmatine. We found that the maximum effect of agmatine on urea synthesis occurred at a concentration of 100 µS in the perfusate. Higher levels (not shown) had little incremental effect on urea synthesis or oxygen consumption. Similarly, no significant differences were found with an arginine concentration above 500 µM or SNP above 25 µM (data not shown). These data indicate that agmatine is taken up by the liver, where it stimulates oxygen consumtion and urea synthesis from glutamine and ammonia. This action is dose dependent, with a maximum effect at perfusate agmatine of 100 µM. It has been shown that agmatine is taken up by system y+, the same system used for arginine and polyamine transport (6, 42). In cultured hepatocytes, approximately 10% of agmatine was converted to putrescine via the agmatinase pathway (6). In the current study, if we assume that 10% MS#E00246-2002, Nissim et al 13 of agmatine uptake was metabolized to putrescine and urea, then the expected amount of urea formed from agmatine will be about 10 nmoles/min/g, which is less than 0.1% of total urea output (Figure 3C). Thus, the amount of urea which may be derived from agmatine is negligible. This conclusion is further supported by the formation of [15N]urea from 15 N- labeled precursors, as indicated below. Effect of arginine, agmatine or nitric-oxide on hepatic metabolic activity and nitrogen balance: Viability of the livers is documented by the constancy of oxygen consumption during the course of perfusions (Figure 4). The addition of arginine or agmatine to the perfusate significantly (p<0.05) increased O2 consumption between 40-70 minutes. The increased O2 consumption was more significant with agmatine (p=0.006) than with arginine (Figure 4B). The increased oxygen consumption in perfusions with agmatine was associated with stimulated glutamine-N uptake (Table 1 and Figure 4A). Between 40-70 minutes of perfusion, glutamine-N uptake (nmoles N/min/g) was increased by approximately 4-fold in perfusions with agmatine and by 2-fold in perfusion with arginine or SNP (Table 1). There were only minor differences in ammonia-N uptake between the various experimental groups (Table 1 and Figure 4C). The appearance of nitrogen in urea, alanine and glutamate represents the major nitrogenous output (the release of other amino acids was minor). Therefore, we calculated the extent to which these three compounds could account for nitrogen balance across the liver. In control experiments the combined nitrogenous uptake of glutamine and ammonia nitrogen was 1127 nmoles of nitrogen/min/g liver. The output of nitrogen in urea, alanine and glutamate was 1031 nmoles N/min/g liver. In perfusions with agmatine, arginine or SNP, the uptake of nitrogen was 2652, 1610 or 1652 nmoles N/min/g liver, respectively (Table 1). The output of nitrogen was 1890, 1332 or 1225 nmoles N/min/g liver, in perfusion with agmatine, arginine or SNP, respectively (Table 1). Thus, in control perfusions and with the addition of arginine or NSP there was almost complete MS#E00246-2002, Nissim et al 14 recovery of nitrogen uptake in the release of urea, alanine and glutamate. In perfusions with agmatine, the uptake of nitrogen exceeded the output by approximately 30%. Table 2 presents the level of hepatic metabolites at the end of 70 minutes perfusion. We present only those metabolites that are directly related to the urea cycle. The agmatine level in control perfusions was 29.6 ± 4.2 nmoles/g, and increased by approximately 10-fold after perfusion with agmatine and 5-fold after perfusion with arginine, but relatively little with SNP. Agmatine significantly increased the level of hepatic ornithine, and most notably the level of NAG, from 28.9 ± 9.6 nmoles/g in control to 47.6 ± 16.6 nmoles/g (p<0.05) following perfusion with agmatine. At the end of arginine perfusion, there was a significant increase in hepatic content of citrulline, ornithine and arginine. The NAG level increased by approximately 20%, but this difference was not significant. In perfusions with SNP, levels of arginine, agmatine or NAG show small changes compared with control perfusions (Table 2). However, there was a significant (p<0.05) depletion of liver glutamine, glutamate, alanine and aspartate, associated with significantly increased ornithine. If 1g of liver contains 53 mg mitochondrial protein (7), the mitochondrial NAG levels would be 0.55, 0.91, 0.64 and 0.51 nmoles/mg protein in control or in perfusions with agmatine, arginine and SNP, respectively. The level of hepatic NAG in control perfusions is in good agreement with mitochondrial NAG levels previously reported (7). Measurements of the lactate/pyruvate ratio in liver extracts at the end of perfusions show little differences among the study groups (data not shown), but the sum of lactate and pyruvate uptake was approximately 1.8 ± 0.5 µmoles/min/g in control perfusions and 3.1 ± 0.2 µmoles/min/g in perfusions with agmatine (N=4, p<0.05). Little differences were found in perfusions with arginine or SNP compared with control. The increased uptake of lactate and pyruvate in perfusions with agmatine was not accompanied by increased glucose output, but a 25% increase in glucose output was found in perfusions with SNP compared with control (600 vs. 800 nmoles/min/g in control or SNP, respectively). MS#E00246-2002, Nissim et al 15 Effect of arginine, agmatine or nitric-oxide on 15N-labeled glutamine metabolism and synthesis of [15N]urea : Hepatic glutamine metabolism and ammonia formation are mediated via flux through the phosphate-dependent glutaminase (PDG) reaction (25, 29, 31, 32). In the current study we estimated flux through PDG as the sum of 15 N-labeled ammonia, urea, alanine and glutamate output in the effluent when [5-15N]glutamine was used as labeled precursor. We have found that the formation of other amino acids accounted for less than 5% of [5-15N]glutamine consumption (32). The calculated flux through the PDG reaction based upon values at 60 minutes of perfusion (Figure 5), shows rates (nmoles/min/g) of: 426 ± 113 (control), 691 ± 190 (plus agmatine, p=0.027), 584 ± 118 (plus arginine, p=0.062) and 433 ± 121 (plus SNP, p>0.05). Therefore, the addition of agmatine to the perfusate significantly stimulated flux through PDG, whereas the addition of arginine is less significant. The PDG reaction converts [5-15N]glutamine to 15NH3, which either can be released in the effluent as 15NH3, incorporated into glutamate by glutamate dehydrogenase (GDH) or into urea via the CPS-I reaction (29, 31, 32). Figure 6A illustrates the production of 15N-labeled urea (sum of Um+1 + Um+2). Between 20 and 40 minutes of perfusion, [15N]urea production from [5-15N]glutamine amounted to 150-190 nmoles N/min/g and continued to increase to 250-280 nmoles N/min/g at 60 min in control perfusions or perfusions with SNP. In perfusions with agmatine, however, [15N]urea production increased to about 431 nmoles N min/g (P=0.02) at 60 minutes. In perfusions with arginine, [15N]urea production increased by approximately 25%, to about 310 nmoles N/min/g, which is not significant (p=0.1). To distinguish between a possible effect of agmatine on glutamine metabolism via the mitochondrial PDG reaction or via the deamination of glutamate in the GDH reaction (29, 31, 32), perfusions were performed with [2-15N]glutamine. As with [5-15N]glutamine, perfusions with [215 N]glutamine demonstrate a control rate of [15N]urea production of 100-150 nmoles/min/g between 20-40 minutes and 260 nmoles/min/g from 40-60 min. In perfusions with arginine, [15N]urea output MS#E00246-2002, Nissim et al 16 was 320 nmoles N/min/g at 60 min., but these differences are not significant. However, in perfusions with agmatine [15N]urea production increased to about 600 nmoles N/min/g from 40-60 min (p=0.001) (Figure 6B). Similarly, livers infused with SNP increased [15N] urea production from [2-15N]glutamine to 385 nmoles N min/g from 40-60 min. (p=0.026). Incorporation of 15NH4Cl into urea - Regulation by agmatine: To determine whether the stimulatory action of agmatine on [15N]urea production from 15 N-labeled glutamine is directly linked to glutamine metabolism, or is subsequent to a stimulation of carbamoyl phosphate synthesis, perfusions were carried out with agmatine, 15 NH 4 Cl and unlabeled glutamine. Figure 6C demonstrates that the presence of 15NH4Cl resulted in an immediate and massive production of [15N]urea (between 20-60 minutes) of about 1000 nmoles N/min/g (control) and 1700 nmoles N/min/g (plus agmatine, p<0.05). Agmatine, independent of an effect on glutamine metabolism, increased [15N]urea production from 15NH4Cl by approximately 40% (p<0.05) even with 1mM unlabeled glutamine in the perfusate. Thus, it is reasonable to propose that agmatine stimulates the formation of 15 N-labeled carbamoyl phosphate from 15 NH 4 Cl secondary to its action on mitochondrial NAG synthesis. Synthesis of N-acetylglutamate - Regulation by agmatine: Figure 7 illustrates the relationship between hepatic agmatine and NAG levels under all experimental conditions. Data points from perfusions with [2-15N]glutamine, [5-15N]glutamine and 15NH4Cl are included in these correlation analyses. It is evident that there was a highly significant relationship between hepatic agmatine and NAG concentrations (p=0.0003), between agmatine and [15 N]urea synthesis (p=0.0002), and between NAG and [15N]urea synthesis (p=0.0002). This observation indicates that the level of NAG in the liver is strongly associated with levels of agmatine, and that agmatine may regulate the synthesis of NAG in the mitochondria. This observation suggests that agmatine MS#E00246-2002, Nissim et al 17 regulates NAG synthesis independent of an effect on flux through PDG or GDH, and this action of agmatine on NAG synthesis may play a key role in the regulation of hepatic ureagenesis. The data suggest that increased hepatic agmatine can induce NAG and urea synthesis. A related question is: Does this change reflect increased synthesis or diminished translocation of NAG from mitochondrion to cytosol, where it is degraded? To answer this question we have measured the production of 15 15 N-labeled NAG in liver perfused with 15NH4Cl and unlabeled glutamine or [2- N]glutamine and unlabeled ammonia (See Methods). With 15NH4Cl, [15N]glutamate will be formed in the mitochondria via the reductive amination of _-ketoglutarate catalyzed by glutamate dehydrogenase. In addition, some mitochondrial glutamate will be derived from unlabeled glutamine via the PDG pathway. In the case of [2-15N]glutamine, [15N]glutamate will be formed via the PDG pathway (29, 31, 32). Therefore, the 15N-labeled NAG will be derived from the overall 15 N-labeled mitochondrial glutamate pool regardless of the respective fluxes through PDG or GDH. Figure 8 illustrates the production of 15N-labeled NAG from [2-15N]glutamine. Similar observations were obtained with 15 NH4Cl (data not shown). These data clearly demonstrate that agmatine significantly (p<0.001) increased the synthesis of NAG compared with perfusions without agmatine. Arginine marginally increased (p=0.08), and SNP significantly (p=0.002) decreased the production of 15N-labeled NAG (Figure 8). MS#E00246-2002, Nissim et al 18 Discussion The liver regulates whole body nitrogen metabolism and detoxifies ammonia via conversion of excess amino-N to urea (25, 49). Although ureagenesis is a major hepatic function, the factors that regulate this process have been the subject of conflicting views and controversies (4). Understanding these mechanism(s) is important not only in terms of understanding ammonia detoxification, but also because ureagenesis has provided insight into whole body nitrogen balance and metabolism (49). The initial step in urea synthesis is the conversion of NH4+ and HCO3- to carbamoyl phosphate by mitochondrial CPS-I (28), which requires NAG as an obligatory effector (9, 13, 16, 25). Increased NAG synthesis and concomitant ureagenesis have been observed after the intake of arginine or a high protein diet (8, 19, 27, 44). However, it is unknown whether arginine itself or a metabolite of arginine stimulates the synthesis of NAG, and, thereby, ureagenesis. In the current investigation we have explored the hypothesis that agmatine, not arginine, is responsible for increased NAG synthesis and concomitant ureagenesis. The principal observations supporting our hypothesis are as follows: (i) L-[guanidino-15N2]arginine enters the liver via the portal vein and is catabolized to [guanidino-15N2]agmatine and [15N2]urea. About 90% of arginine uptake was recovered in urea and agmatine output in the effluent, (ii) Metabolism of perfusate arginine is primarily mediated via the mitochondrial ADC and arginase II reactions, (iii) Agmatine, which is naturally present in the intact liver, significantly increases in concentration following addition of arginine to the perfusate. As much as 90% of hepatic agmatine can be derived from arginine entering the liver via the portal vein, MS#E00246-2002, Nissim et al 19 (iv) The increase of hepatic agmatine strongly correlates with an increase of the newly synthesized 15N-labeled NAG and [15N]urea production from either [2-15N] or [5-15N] glutamine and unlabeled ammonia or 15NH4Cl and unlabeled glutamine as nitrogen precursors, (v) Perfusions with agmatine significantly stimulated (p<0.0001) the synthesis of [15N]NAG and [15N]urea from 15N-labeled ammonia or glutamine. However, perfusion with arginine marginally increased (p=0.08) NAG and urea synthesis. The current observations suggest that the arginine entering the liver via the portal vein is mainly metabolized through the mitochondrial ADC and arginase II reactions, and there is minimal or no equilibrium with the arginine pool that is linked with the urea cycle. Evidence of this is the observation that label in [15N]urea was about 5-10% of total urea output with 0.5 mM L-[[5N 2guanidino]arginine, unlabeled ammonia and glutamine in the perfusate. If the perfusate arginine is in equilibrium with the cytosolic pool of arginine formed via the urea cycle, then the isotopic enrichment in hepatic arginine at the end of perfusion should be similar to the isotopic enrichment of [15N2]urea in the effluent. However, the observed enrichment in hepatic [guanidino-15N2]arginine was about 50 MPE and enrichment in effluent urea was about 5-10 MPE at the end of perfusion. This is expected, since the urea cycle does not allow net production of intermediates, and arginine formed is hydrolyzed to ornithine and urea (25, 47). In addition, the percentage of [15N2]urea of the total urea in the effluent indicates that the relative activity of arginase II is about 5-10% of the total (sum of arginase I and II reactions) hepatic arginase. This estimate is in agreement with previous observations indicating that in human liver arginase II is about 2% of liver arginase (17), and in rat about 10% of total arginase activity (8). In addition to mitochondrial formation of [15 N]urea from external L-[guanidino15 N2]arginine, the current study demonstrates that approximately 15-20% of L-[guanidino- 15 N2]arginine uptake was catabolized via the mitochondrial ADC reaction to form agmatine (Figure MS#E00246-2002, Nissim et al 20 2A). O'Sullivan et al (33, 34), suggested that [U-14C]arginine is catabolized in the liver by ornithine aminotransferase following its degradation via arginase I. Their conclusion is based on the release of 14CO2 in the effluent, which was approximately 13 nmoles/min/g in liver obtained from rats fed a normal protein diet (34), a regimen similar to that of this study. If we assume that 1 mole of agmatine formed via the ADC reaction will be accompanied by the release of 1 mole of CO2, then the rate of CO2 formation would be 14-20 nmoles/min/g, similar to the values obtained by O'Sullivan et al (33, 34). Agmatine is widely distributed in mammalian tissues (37, 40, 41), and may have a role in multiple metabolic functions. The current observations demonstrate that agmatine entering the liver via the portal vein stimulates the uptake of glutamine (Table 1), synthesis of NAG and urea (Figures 4-8). This possibility is consistent with an increased rate of oxygen consumption in perfusions with agmatine, and to a less extent with arginine (Figure 4B). The increased oxygen consumption in perfusions with agmatine may indicate (i) higher energy requirements; (ii) stimulation of substrate supply to the electron transport chain and/or (iii) increased respiratory chain activity (45). Agmatine may increase the activity of respiratory enzymes secondary to stimulated mitochondrial energyconsuming functions such as the synthesis of NAG, carbamoyl phosphate and urea itself. In line with the above suggestion are data (Figure 8) demonstrating that agmatine significantly (p<0.0001) increased formation of newly synthesized 15 15 N-labeled NAG from [2- N]glutamine. However, arginine marginally increased (p=0.08), and SNP significantly (p=0.002) decreased the production of NAG compared with control. The linear correlation between agmatine and NAG levels (Figure 7), demonstrates that synthesis of NAG and, thereby, activity of CPS-I would depend upon the concentration of agmatine. Therefore, the current data strongly support the hypothesis that agmatine is a positive effector for mitochondrial synthesis of NAG. In addition, the data in Figure 7 suggest that the concentration of intra-hepatic agmatine is an important feature in the regulation of urea synthesis and ammonia detoxification. Nevertheless, it is uncertain whether MS#E00246-2002, Nissim et al 21 increased NAG levels are secondary to increased synthesis or diminished translocation of NAG from mitochondrion into cytosol, where it is degraded. It has been shown that efflux of NAG out of mitochondria and its subsequent degradation are diminished in rats fed a high protein diet compared with rats on a protein-free diet (22, 23, 26). The current data clearly demonstrate that agmatine stimulates the synthesis of NAG (Figure 8). The diminished rate of NAG translocation in response to a high protein diet (26) may reflect the presence in this diet of arginine, which is converted to agmatine. It has been suggested that hepatic glutaminase, and concomitant glutamate production is a key factor in facilitating NAG synthesis (4). Consistent with this interpretation are the data demonstrating that agmatine stimulates glutamine uptake (Figure 4A and Table 1) and flux through glutaminase (Figure 5). However, the positive association between NAG synthesis, [15N]urea synthesis and agmatine concentration is valid whether [2-15N]glutamine, [5-15N] glutamine or 15 NH4Cl was used to monitor these metabolic processes (Figure 7). Thus, agmatine may regulate NAG synthesis independent of an effect on flux through the PDG reaction. The observed increased flux through PDG in perfusions with agmatine may be subsequent to elevated NAG levels in the mitochondrial matrix, as previously indicated (24). In addition, agmatine-stimulated glutamine uptake (Table 1), may provide an additional fuel that support NAG and/or carbamoyl phosphate synthesis, both of which require ATP. This possibility would be in line with the increased oxygen consumption associated with agmatine induced glutamine uptake (Figure 4A, 4B and Table 1). A significant decrease in hepatic glutamate level was observed following perfusions with SNP (Table 2). This finding may explain the decreased NAG synthesis in perfusions with SNP (Figure 8). The uptake of glutamine is increased with SNP (Table 1 and Figure 4), but this is not reflected in augmented output of [15N]urea and 15NH3 from [5-15N]glutamine (i.e., flux through the PDG pathway), which show minor differences compared with control (Figures 5 and 6). Measurements of 15NH3 enrichment in perfusions with [2-15N]glutamine show that, in control, 15N MS#E00246-2002, Nissim et al 22 ammonia enrichment in the effluent was about 3 atom % excess, with little differences in perfusions with agmatine or arginine. In perfusions with SNP, 15NH3 enrichment was about 11 atom % excess, indicating a stimulation of glutamate catabolism mediated via the GDH reaction. Therefore, stimulated mitochondrial glutamate catabolism in perfusions with SNP may account for the reduction of the newly synthesized NAG as shown in Figure 8. An alternative mechanism for increased NAG synthesis by agmatine is stimulation of `oxidation and/or the pyruvate dehydrogenase and pyruvate carboxylase reactions. This possibility is supported by the significant (p<0.05) increase in oxygen consumption in perfusions with agmatine (Figures 3B and 4B), and increased pyruvate and lactate uptake (µmoles/min/g) from 1.7 (control) to about 3 (plus agmatine). Stimulation of the pyruvate dehydrogenase reaction would increase availability of acetyl-CoA for NAG synthesis, and an increase in the pyruvate carboxylase activity would provide more oxaloacetate, and thereby, aspartate for synthesis of argininosuccinate, as we have previously indicated in studies with [3-13C]pyruvate (31). These possibilities are currently under investigation using 13C-labeled precursors and NMR as an analytical tool. In conclusion, the current observations provide evidence to support the hypothesis that availability of agmatine rather than arginine may have a major regulatory role in hepatic ammonia detoxification and urea synthesis. The current findings may have clinical implications for the treatment of disturbed urea synthesis and toxic hyperammonemia. Effective drugs have been introduced for removal of waste toxic nitrogen from the body (10), but agmatine might prove a valuable therapeutic adjunct. This agent has been proposed as a treatment for other disorders (40, 41). Agmatine can be useful especially in cases of hyperactivity of the hepatic GDH reaction, which is associated with the hyperinsulinism/hyperammonemia syndrome (43). In this case, hyperammonemia is secondary to diminished synthesis of NAG (43). The stimulation of NAG synthesis by agmatine (Figure 8), lends support to the importance of agmatine as a potential candidate for the treatment of hyperinsulinism/hyperammonemia syndrome in infants (43) MS#E00246-2002, Nissim et al 23 ACNOWLEDEGEMENT We thank Dr. J. T. Brosnan for helpful discussion. This work was supported by United States National Institutes of Health Grants DK-53761 and CA-79495 (to I.N). MS#E00246-2002, Nissim et al 24 REFERENCES: 1. Bergemeyer, H.U., Bernt. E, Schidt F., and Stork H. D-Glucose In: Methods of Enzymatic Analysis. H.U.Bergemeyer, Ed., Academic press, 3: PP, 1198-1201, 1974 2. Blantz R.C., Satriano J., Gabbai F., Kelly C. Biological effects of arginine metabolites, Acta Physiol. Scand 168: 21-25, 2000. 3. Brosnan J.T., Brosnan M.E., Charron R. and Nissim I. A mass isotopomers study of urea and glutamine synthesis from 15N-labeled ammonia in the perfused rat liver. J. Biol. Chem. 271: 16199-16207, 1996. 4. Brosnan J. T. Glutamate, at the interface between amino acid and Carbohydrate Metabolism J. Nut. 130: 988S-990S, 2000. 5. Brosnan J.T., Brosnan M.E., Yudkoff M., Nissim-Ilana, Daikhin Y., Lazarow A., Horyn O., and Nissim I: Alanine metabolism in the perfused rat liver J. Biol. Chem. 276: 31876-31882, 2001. 6. Cabella C., Gardini G., Corpillo D., Testore G., Bedino S., Solinas S.P., Cravazola C., Vargiu C., Grillo M.A., and Colombatto S. Transport and metabolism of agmatine in rat hepatocyte cultures, Eur. J. Biochem. 268: 940-947, 2001. 7. Carey G.B., Cheung Chia-Wei, Cohen N.S., Brusilow S., and Raijman L. Regulation of urea and citrulline synthesis under physiological conditions 292: 241-247, 1993. 8. Cheung C. W, and Raijman L. Arginine, mitochondrial arginase, and the control of carbamyl phosphate synthesis. Arch. Biochem. Biophys. 209: 643-649, 1981. 9. Cheung C. W., Cohen N. S. , and Raijman L. Channeling of urea cycle intermediates in situ in permeabilized hepatocytes. J. Biol. Chem. 264:4038-4044, 1989. 10. Come B., Kasumov T., Bradley P.A., Puchowicz M.S., Dahms W., Kerr D., Nissim, I., and Brunengraber H. Identification of phenylbutyrylglutamine, a new metabolite of phenylbutyrate metabolism in humans J. Mass Spectrometry. In. Press, 2002. 11. Curran R.D., Billiar T.R., Stuehr D.J., Hofmann K., Simmons R.L. Hepatocytes produce nitric oxides from L-arginine in response to inflammatory products of Kupffer cells. J. Exp. Med. 170P 1769-1774, 1989. 12. Czok, R., Lamprecht, W. Pyruvate, phosphoenolpyruvate and D-glycerate-2-phosphate In: Methods of Enzymatic Analysis. H.U.Bergemeyer, Ed., Academic press, 3: PP, 1446-1451, 1974. 13. Fahien L.,Schoder J.M., Gehred G.A. and Cohen P.P. Studies on the mechanism of action of N-acetylglutamate as an activator of carbamoyl phosphate synthetase. J. Biol. Chem. 239: 1935-1942, 1964. MS#E00246-2002, Nissim et al 25 14. Gen L., Regunathan D., Barrow C.J., Esraghi J., Cooper R., and Reis D.J. Agmatine: an endogenous clonidine-displacing substance in the brain Sicence 263: 12231-12234, 1994. 15. Gutmann, I., Wahlefeld A W. L-(+)-Lactate In: Methods of Enzymatic Analysis. H.U.Bergemeyer, Ed., Academic press, 3: PP, 1464-1472, 1974. 16. Hall L.M., Metzenberg R. L. and Cohen P.P. Isolation and characterization of a naturally ocurring cofactor of carbamyl phosphate biosynthesis. J. Biol. Chem. 230: 1013-1021, 1958. 17. Iyer R., Jenkinson C.P., Vockley J., G., Kern R., M., Grody W., W., and Cederbaum S. The human arginases and arginase deficiency, J. Inher. Metab. Dis. 21: 86-100, 1998. 18. Jones, B.N and Gilligan, J.P. Ortho-phthaldialdehyde precolumn derivatization and reversed-phase high-performance liquid-chromatography of polypeptide hydrolysates and physiological fluids J. Chromatogr. 266, 471-482, 1983. 19. Kamemoto E.S., Atkinson D.E. Modulation of the activity of rat liver acetylglutamate synthase by pH and arginine concentration, Arch. Biochem. Biophy. 243: 100-107, 1985. 20. Krebs, H. A., and Henseleit K. Investigation of urea metabolism in the animal body. Hoppe-Syeler's Z. Physiol. Chem. 210: 33-36, 1932. 21. Marletta M.A. Nitric oxide synthase structure and mechanism J Biol Chem 268: 966969, 1993. 22. Meijer A. J., and Van Woerkom G. M. Turnover of N-acetylglutamate in isolated rat hepatocytes Biochimica et Biophysica Acta 721: 240-246, 1982. 23. Meijer A. J., Van Woerkom G. M., Wanders R. J. A., and Lof C. Transport of Nacetylglutamate in rat-liver mitochondria, Eur. J. Biochem. 124: 325-330, 1982. 24. Meijer A. J., and Verhoeven A.J. Regulation of hepatic glutamine metabolism. Biochem. Soc. Trans. 14: 1001-1004, 1986. 25. Meijer A.J., Lamers W.H. and Chamuleau R.A.F.M Nitrogen metabolism and ornithine cycle function . Physiological Reviews 70: 701-748, 1990. 26. Morita T., Miura S., Mori M., Tatibana M. Transport of the precursor for rat-liver ornithine carbamoyltransferase into mitochondria in vitro E. J. Biochem. 91: 563-569, 1982. 27. Morimoto H., Brady J.F., and Atkinson D.E. Effect of Level of Dietary Protein on Arginine-Stimulated Citrulline Synthesis, Biochem. J. 272: 671-675, 1990. MS#E00246-2002, Nissim et al 26 28. Nissim I.,Yudkoff M., Terwilliger T., and Segal S. Rapid determination of [guanidine15 N]Arginine in plasma with Gas Chromatography-Mass Spectrometry: Application to human metabolic studies, Anal. Biochem.131: 75-82, 1983. 29. Nissim I., Cattano C., Nissim Ilana and Yudkoff M. Relative role of the glutaminase, glutamate dehydrogenase, and AMP-deaminase pathway in hepatic ureagenesis: studies with 15N and GC-MS. Arch. Biochem. Biophys. 292: 393-401, 1992. 30. Nissim, I, Weinberg J.M. Glycine attenuates Fanconi syndrome induced by maleate or ifosfamide in rats (1996) Kidney Int. 49: 684-695, 1996. 31. Nissim I., Yudkoff M., and Brosnan J.T. Regulation of [15N] urea synthesis from [515N] glutamine: Role of pH, hormones and pyruvate. J. Biol. Chem. 271: 31234-31242, 1996. 32. Nissim I., M.E. Brosnan,Yudkoff M., Ilana Nissim and Brosnan J.T: Studies of hepatic glutamine metabolism in the perfused rat liver with 15N-labeled glutamine. J. Biol. Chem. 274: 28958-28965, 1999. 33. O'Sullivan D., Brosnan J.B., and Brosnan M.E. Hepatic zonation of the catabolism of arginine and ornithine in the perfused rat liver, Biochem. J. 330: 627-632, 1998. 34. O'Sullivan D., Brosnan J.B., and Brosnan M.E. Catabolism of arginine and ornithine in the perfused rat liver: Effect of dietary protein and of glucagons, Am. J. Physiol. 278: E516-E521, 2000. 35. Pastor C.M., and Billiar T.R., Nitric oxide causes hyporeactivity to phenylephrine in isolated perfused livers from endotoxin-Treated Rats, Am. J. Physiol. 268: G177-G182, 1995. 36. Paxinou E., Weisse M., Chen Q., Souza J.M., Hertkon C., Selak M., Daikhin E., Yudkoff M., Sowa G., Sessa W.C., and Ischiropoulos H. Dynamic regulation of metabolism and respiration by endogenously produced nitric oxide protects against oxidative stress, Proc. Natl. Acad. Sci. 98: 11575-11580, 2001. 37. Rasch W., Regunathan S., Li G., and Reis D.J. Agmatine, the bacterial amine, is widely distributed in mammalian tissues, Life Sci. 56: 2319-2330, 1995. 38. Reczkowski R. S., and Ash D.E. Rat liver arginase: kinetic mechanism, alternate substrates, and inhibitors. Arch.. Biochem. Biophy. 312: 31-37, 1994. 39. Regunathan S., and Reis D.J. Characterization of arginine decarboxylase in rat brain and liver: Distinction from ornithine decarboxylase, J. Neurochemistry 74: 2201-2208, 2000. 40. Reis D.J., and Regunathan S. Is agmatine a novel neurotransmitter in brain? Trends Pharmacl. Sci. 5:187-193, 2000. 41. Satriano J., Kelly C.J., and Blantz R.C. An emerging role for agmatine, Kidney Inter. 56: 1252-1253, 1999. MS#E00246-2002, Nissim et al 27 42. Satriano J. Isome M., Casero R.A., Thomson S.C., and Blantz R.C. Polyamine transport system mediates agmatine transport in mammalian cells, Am. J. Physiol. Cell Physiol. 281: C329-C334, 2001. 43. Stanely C. A., Lieu BS, Hsu BYL, Alberto BB, Greenberg CR, Hopwood NJ, Perlman K, Rich BH, Zammarchi E, and Poncz M. Hyperinsulinism and hyperammonemia in infants with regulatory mutations of the glutamate dehydrogenase gene. New England J. Medicine, 338:1352-1357,1998. 44. Stewart P.M., Walser M. Short term regulation of ureagenesis. J. Biol. Chem. 255: 5270-5280, 1980. 45. Weinberg JM, Venkatachalam MA, Roeser NF, and Nissim I. Mitochondrial dysfunction during hypoxia/reoxygenation and its correction by anaerobic metabolism of citric acid cycle intermediates. Proc. Nat. Acad. Sci., USA, 97:2826-2831, 2000. 46. Wolfe R.R. Measurment of urea kinetics in vivo by means of a constant tracer infusion of di-15N-urea.. Am. J. Physiol. 240: E428-E434, 1981. 47. Wu G., and Morris S.M. Arginine metabolism: Nitric oxide and beyond, Biochem. J. 336: 1-17, 1998. 48. Yang D., Puchowicz A., David F., Powers L., Halperin M.L., and Brunengraber H. 15N enrichment of ammonium, glutamine-amide and urea, measured via Mass Isotopomer analysis of hexamethylenetetramine, J. Mass Spectrom. 34: 1130-1136, 1999. 49. Young V.R., and Ajami A.M. Glutamine: The emperor or his clothes? J. Nutr. 131: 2449S-2459S, 2001. MS#E00246-2002, Nissim et al 28 TABLE 1 Effect of Agmatine, Arginine or SNP on Nitrogen Balance. Experimental Group Control (+) Agmatine (+) Arginine (+) SNP ( nmoles / g /min ) Nitrogen Uptake From Glutamine-N 466 ± 64 1877 ± 150 812 ± 50 1020 ± 218 From Ammonia-N 661 ± 24 775 ± 29 798 ± 20 632 ± 19 As Urea-N 961 ± 87 1745 ± 99 1228 ± 120 1146 ± 24 *As Amino-N 70 ± 13 145 ± 24 104 ± 15 79 ± 18 Nitrogen Output ` Values are means ± S.D of data points presented in Figure 4 between 40-70 minutes of perfusions. Values of total glutamine and urea N are nmoles/min/g X 2. *Amino-N represents the sum of glutamate and alanine nitrogen. MS#E00246-2002, Nissim et al 29 Table 2 Hepatic Content of Agmatine, N-Acetylglutamate and Amino Acids Following 70 Minutes of Perfusion With Either Agmatine, Arginine or SNP Experimental Group Metabolite Control (n=15) (+)Agmatine (n=18) (+)Arginine (n=9) (+)SNP (n=6) Agmatine1 29 ± 4 311 ± 61.8** 129 ± 40** 27 ± 3 N-Acetylglutamate1 28.9 ± 8.6 Ornithine1 46.2 ± 15.7 71.9 ± 33.7** 134 ± 37** 74 ± 8** Citrulline1 66 ± 25 57 ± 26 96 ± 34** 52 ± 15 Arginine1 51 ± 21 69 ± 17 148 ±37** 75 ± 22 Glutamine2 2.4 ± 0.7 2.7 ± 0.8 1.8 ± 0.3 1.6 ± 0.4** Glutamate2 1.31 ± 0.4 1.6 ± 0.7 1.5 ± 0.5 0.8 ± 0.2** Alanine2 0.9 ± 0.2 0.9 ± 0.3 0.8 ± 0.1 0.5 ± 0.1** Aspartate2 0.29 ± 0.1 0.36 ± 0.1 0.28 ± 0.1 0.19 ± 0.1** 47.6 ± 16.6** 33.6 ± 8.5 26.8 ± 6.7 Perfusions performed as described in Materials and Methods. Livers were perfused with either agmatine (100 µM), arginine (500 µM) or SNP (25 µM). Perfusions were terminated as described and processed for quantitation of the indicated amino acid and agmatine. Perfusate also included glutamine (1 mM), NH4Cl (0.3 mM), lactate (2.1 mM) and pyruvate (0.3 mM). Each result is expressed as the mean value ± SD. The number of experimental determinations is given in parentheses for each experimental group. ** P < 0.05, that is, significantly different from control perfusions. 1 – nmol/g wet weight 2 - µmol/g wet weight MS#E00246-2002, Nissim et al 30 FIGURE LEGENDS FIGURE 1: Schematic presentation of external L-[guanidino-15N2]arginine metabolism and formation of [guanidino-15N2]agmatine and [15N 2]urea in the liver. This schema reflects perfusions with L-[guanidino-15N2]arginine and unlabeled glutamine (1mM) and ammonia (0.3mM). Enzymatic steps presented are: 1, nitric oxide synthetase; 2, arginine decarboxylase; 3, arginase II; 4, N-acetylglutamate synthetase; 5, glutamate dehydrogenase (GDH); 6, phosphate-dependentglutaminase (PDG); 7, carbamoyl phosphate synthetase I (CPS-I); and 8, arginase I. ? indicates a questionable pathway ; + , indicates stimulatory action or co-factor. FIGURE 2: Uptake of perfusate L-[guanidino-15N2]arginine (------), and B, the output in the effluent, of [15N2]urea (---L--); [guanidino - 15N2]agmatine (---6--) ; and nitric oxide as NO-2+NO-3 (------). Also is shown oxygen consumption during the course of perfusions (----). Bottom panel C, represents the 15N enrichment in arginine and agmatine in liver extracts at the end of 70 minutes perfusions. Bars are means ± S.D for three livers. FIGURE 3: Dose dependence of agmatine uptake (A), oxygen consumption (B) and total urea N output (C), during the course of perfusions with increasing concentration of agmatine. Perfusions were carried out with a basic perfusate for 15 minutes and then perfusate was replaced by perfusate contained 1mM glutamine, 0.3 mM ammonia and the indicated concentration of agmatine. The dose dependence was studied in the same liver, and at the indicated time a separate perfusate reservoir containing the indicated agmatine concentration was used. Bars are means ± S.D for three livers. MS#E00246-2002, Nissim et al 31 FIGURE 4: Total nitrogen balance and O2 consumption in perfused liver. Livers were perfused without (control) or with either agmatine (100 µM), SNP (25 µM) or arginine (500 µM) and 1mM [2-15N]- or [5-15N]-labeled glutamine and 0.3 mM unlabeled ammonia or 15NH4Cl and unlabeled glutamine as nitrogen precursors. Since all perfusions were performed with the same concentrations of substrates (i.e., 1 mM glutamine, 0.3 mM NH4Cl, 2.1 mM lactate and 0.3 mM pyruvate), it was possible to combine those with 15N-labeled glutamine or ammonia. The data are the means ± S.D, for control perfusions (--O--, N=15), plus agmatine (--G--, N=18), plus arginine (-6,--, N=9) and plus SNP (--▲--, N=6). Values of total glutamine and urea N are nmoles/min/g of these metabolites times 2. Panel A, represents glutamine N uptake; B, O2 consumption; C, ammonia uptake; and D, nitrogen output. FIGURE 5: Flux through the PDG reaction during the course of liver perfusions. Values for PDG were estimated from perfusions with [5-15N]glutamine as detailed in (31, 32). The data are the means ± S.D, for control perfusions (--O--, N=5), plus agmatine (--G--, N=5), plus arginine (-6,--, N=5) and plus SNP (--▲--, N=4). FIGURE 6: Total [15N]urea production (sum of Um+1 and Um+2), during the course of liver perfusion with [5-15N]glutamine (panel A), [2-15N]glutamine (panel B) and 15NH4Cl (panel C). Perfusion conditions are as indicated in Figure 4. The output of 15N-labeled urea is the product of 15 N enrichment (mole % excess/100) times one half of total urea N (nmoles N/min/g) for Um+1, and times total urea N for Um+2. The data are the means ± S.D, for control perfusions (--O--, N=3-5), plus agmatine (--G--, N=4-6), plus arginine (--6,--, N=3-5) and plus SNP (--▲--, N=3-4). MS#E00246-2002, Nissim et al 32 FIGURE 7: The dependence of NAG levels and urea synthesis on agmatine concentrations in freeze-clamped liver at the end of perfusions. Data points included in these correlations are from perfusions with [2-15N]glutamine, [5-15N]glutamine and 15NH4Cl of control, plus agmatine, arginine and SNP perfusions. The bottom panels represent the relationship between hepatic agmatine or NAG in freeze-clamped liver extracts and [15N]urea in the effluent at the end of perfusions. FIGURE 8: Appearance of 15N-labeled N-acetylglutamate in extracts of livers that were freeze-clamped at the end of perfusions with [2-15N]glutamine. Amount of 15N-labeled NAG is the product of isotopic enrichment (MPE/100) times total concentration (nmoles/g). The data are the means ± S.D, for control perfusions (N=4), plus agmatine (N=4), plus arginine (N=3) and plus SNP ( N=3). p values indicate the degree of significance compared with control level. MS#E00246-2002, Nissim et al 33 Figure 1 Nissim et. al. Cytosol 15NO Mitochondrion + [15N] Citrulline 3 [15N2] Urea + Ornithine 1 L-[guanidino-15N2] Arginine 15N 2 2- Arginine [guanidino-15N2] Agmatine CO2 (?) } (?)[15N2] Urea [14N2] Urea 8 Urea ArginineCycle Ornithine NAG HCO–3 CP 7 + NH NH4Cl Glutamine + Acetyl-CoA 4 + 5 4 + Glutamine 6 Glutamate MS#E00246-2002, Nissim et al 34 MS#E00246-2002, Nissim et al 35 MS#E00246-2002, Nissim et al 36 MS#E00246-2002, Nissim et al 37 MS#E00246-2002, Nissim et al 38 Nissim et. al. Total [15N] Urea Nitrogen Output (nmoles/min/g) Figure 6 600 550 500 450 400 350 300 250 200 150 100 50 0 A 15 20 25 30 35 40 45 50 55 60 Total [15N] Urea Nitrogen Output (nmoles/min/g) Time (min) 600 550 500 450 400 350 300 250 200 150 100 50 0 B 15 20 25 30 35 40 45 50 55 60 Total [15N] Urea Nitrogen Output (nmoles/min/g) Time (min) 2500 2250 C 2000 1750 1500 1250 1000 750 500 250 0 15 20 25 30 35 40 45 50 55 60 Time (min) 65 70 MS#E00246-2002, Nissim et al 39 MS#E00246-2002, Nissim et al 40