Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
History of genetic engineering wikipedia , lookup
Site-specific recombinase technology wikipedia , lookup
Expanded genetic code wikipedia , lookup
Artificial gene synthesis wikipedia , lookup
Genetic code wikipedia , lookup
Zinc finger nuclease wikipedia , lookup
Therapeutic gene modulation wikipedia , lookup
Genome editing wikipedia , lookup
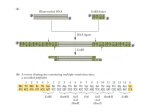
Current Biology, Vol. 15, 73–78, January 11, 2005, ©2005 Elsevier Ltd. All rights reserved. DOI 10.1016/j .c ub . 20 04 . 12 .0 4 2 A Single Amino Acid Change in the SPRY Domain of Human Trim5␣ Leads to HIV-1 Restriction Melvyn W. Yap, Sébastien Nisole,1 and Jonathan P. Stoye* Division of Virology National Institute for Medical Research The Ridgeway Mill Hill London NW7 1AA United Kingdom Summary Retroviral restriction factors are cellular proteins that interfere with retrovirus replication at a postpenetration, preintegration stage in the viral life cycle [1–3]. The first restriction activity described was the mouse Fv1 gene [4]. Three different alleles of Fv1, capable of restricting different murine leukaemia viruses (MLV), have been characterized at the molecular level [5, 6]. Two further activities, Ref1, which acts on MLV, and Lv1, which acts on lentiviruses, have been identified in other mammalian species [7–9]. Recently, it has become clear that Ref1 and Lv1 are encoded by the same gene, Trim5␣, which inhibits retrovirus replication in a species-specific manner [10–14]. A series of chimeras between the human and rhesus monkey Trim5 genes were created to map and identify these specificity determinants. The Trim5␣ SPRY domain was found to be responsible for targeting HIV-1 restriction. By contrast, N-MLV restriction appears dependent on both the coiled-coil domain and the SPRY domain. A single amino acid substitution (R332P) in the human Trim5␣ can confer the ability to restrict HIV-1, suggesting that small changes during evolution may have profound effects on our susceptibility to cross-species infection. Results and Discussion Human Trim5␣ restricts N-tropic MLV, but not B-tropic MLV or HIV-1, whereas rhesus monkey Trim5␣ restricts N-tropic MLV and HIV-1, but not B-tropic MLV [11–14]. The human and rhesus forms of Trim5␣ can also be distinguished by using the L117H mutation of N-MLV, which is restricted by human, but not rhesus monkey Trim5␣ [13]. A series of chimeras between the human and rhesus gene was constructed with overlapping PCR (Figure 1) to examine which domain of Trim5␣ was involved in determining the specificity of retroviral restriction. The structures of these chimeras (and all subsequent constructs) were confirmed by sequencing. Trim5␣ has a modular design with a series of distinct motifs. These include the RING motif, a B-Box, and a coiled-coil domain, which together form the tripartite RBCC domain characterizing Trim proteins, as well as *Correspondence: [email protected] 1 Present address: CNRS UPR 9051, Centre Hayem, Hôpital SaintLouis, 16, rue de la Grange-aux-Belles, 75010 Paris, France. a SPRY domain [15]. These different domains were exchanged between the two proteins. The chimeras HR3, HR2, and HR1 contain the human Trim5␣ RING motif with progressively longer substitutions of rhesus C-terminal residues. A reciprocal series was also made (RH3, RH2, and RH1) where parts of the rhesus Trim5␣ are substituted by human sequences. Restriction was measured by a previously described two-color FACS assay [13, 16]. Briefly, nonrestricting cells were transduced with an MLV-based vector carrying the Trim gene and EYFP. Two days posttransduction, the cells were challenged with tester viruses incorporating various capsids and encoding the EGFP marker. The percentage of Trim/ EYFP positive cells that were transduced with the EGFP marker was then scored against that of Trim/EYFP negative cells. Restriction was expressed as a ratio of EGFP⫹, EYFP⫹ cells to EGFP⫹, EYFP⫺ cells. A ratio of less than or equal to 0.3 was taken as restriction, whereas a ratio between 0.7 and 1 indicated the absence of restriction. A typical result is shown in Figure 1A. As observed previously, human Trim5␣ restricted N-MLV and the L117H mutant of N-MLV, but not B-MLV or HIV-1 [13]. Rhesus Trim5␣ restricted HIV-1 as well as N-MLV, but not B-MLV or N L117H. These restricting phenotypes were used as standards for comparison with the restriction profile of the chimeric molecules. All chimeras could restrict N-MLV, but not B-MLV, indicating that the proteins were still functional after exchanging domains (Figure 1B). Substitution of the SPRY domain from the human Trim5␣ with the rhesus SPRY domain in HR3 is sufficient to confer restriction to HIV-1. The reciprocal chimera, where the SPRY of rhesus Trim5␣ was replaced with the human form (RH3), lost the ability to restrict HIV-1. This implied that the SPRY domain of Trim5␣ was sufficient and necessary for recognizing and restricting HIV-1. The determinants for N-MLV restriction seemed to be more complex. Replacing the SPRY domain of rhesus Trim5␣ with the human form (RH3) enabled it to restrict N L117H in addition to N-MLV, implying that the SPRY domain was also involved in targeting the restriction of N-MLV. However, substitution of the human SPRY domain with the corresponding rhesus sequences (HR3) did not abolish its ability to restrict N L117H. It was only when the coiledcoil domain as well as the SPRY of human Trim5␣ was replaced with that from rhesus (HR2) that restriction of N L117H was lost. These results suggested that both the coiled-coil and the SPRY domains play roles in determining the specificity of MLV restriction. Two more sets of chimeras were constructed (Figure 2) to further delineate the regions of the SPRY domain involved in restriction. HR4 to HR7 consisted of human Trim5␣ with decreasing lengths of rhesus sequences substituted within the SPRY domain, whereas RH4 to RH7 formed a reciprocal series where the rhesus Trim5␣ was substituted with human SPRY sequences. As expected, all the chimeras could restrict N-MLV, but not B-MLV, indicating that they were functional with respect to restriction. HR4 was able to restrict HIV-1, but the Current Biology 74 Figure 1. Restriction of HIV-1 and MLV by Human, Rhesus, and Chimeric Trim5␣ (A) Graphical representation of restriction by human and rhesus Trim5␣. HT1080 and M. dunni cells were transduced with MLV-based vectors carrying Trim5␣ and EYFP at a multiplicity of infection of 1. Two days posttransduction, HT1080 cells were challenged with HIV-1 vectors carrying EGFP, whereas M. dunni cells were challenged with either N-MLV, B-MLV, or N L117H. The ratio of the percentage of Trim5␣⫹ cells versus that of Trim5␣⫺ cells that were infected was determined by FACS analysis 2 days after challenging with tester virus. A ratio of between 0.7 to 1 indicates the absence of restriction, whereas a ratio of less than 0.3 indicates the presence of restriction. Error bars represent the standard error of the mean. (B) Schematic representation of the human/ rhesus chimeric Trim5␣ constructs and their ability to restrict HIV-1, N-MLV, B-MLV, and N L117H. Restriction assays were performed as described in (A). White boxes indicate human derived sequences; gray boxes show rhesus monkey sequences. The amino acid positions corresponding to the human Trim5␣ are indicated on the top of the figure, whereas the ones corresponding to the rhesus sequence are indicated at the bottom. restriction was absent when the rhesus sequences after residue 374 were replaced with human ones from HR5 to HR7. This suggested that the region determining specificity for HIV-1 restriction was between amino acids 322 to 374 of the rhesus Trim5␣. RH4, which does not restrict HIV-1, lacked the rhesus sequence in this region. When the rhesus sequence was included in the chimera RH5, restriction of HIV-1 was restored, confirming the requirement of this region in HIV-1 recognition and restriction. The requirements for MLV restriction were found to be somewhat different from that of HIV-1. HR4 to HR7 could restrict N L117H. This was expected as both human Trim5␣ and HR3 (Figure 1B) could restrict this mutant. The chimeras RH4 to RH7 (Figure 2) contain decreasing amount of human SPRY sequences compared to RH3 (Figure 1B). RH4 and RH5 could restrict N L117H, but the restriction was abolished when human residues before amino acid 401 were excluded, in RH6 and RH7. This suggested that the SPRY region in human Trim5␣ influencing N-MLV restriction involved residues up to amino acid 400. Taken with the previous data, these results indicated that the specificity determinants of MLV restriction encompassed a region somewhat larger than that which governed HIV-1 restriction. We next sought to identify individual residues between amino acids 322 and 374 in the SPRY domain that affected restriction. Between the human and rhesus sequence, there was a difference of seven amino acids, two contiguous, the others scattered throughout the region. In addition, a stretch of six residues starting from amino acid 335 in the human sequence was replaced by 8 different residues in the rhesus sequence. The residues at these positions in the human Trim5␣ were replaced with the corresponding residues from rhesus monkey by site-directed mutagenesis. The mutants were then assayed for restriction of HIV-1 and MLV (Figure 3). Only two positions in this region affected restriction to HIV-1. The arginine to proline change at residue 332 and the exchange of the six human residues at position 335 for the eight amino acid rhesus sequence conferred the ability to restrict HIV-1 to the human Trim5␣. Conversely, changing each of these positions in the rhesus Trim5␣ to the corresponding residues in the human protein did not abolish restriction of HIV-1. Indeed, when both positions of the human protein were substituted by the rhesus residues, the resulting double mutant had a restricting phenotype that was similar to the two single mutants. This suggested that both of Mapping Restriction Specificity Domains of Trim5␣ 75 Figure 2. Restriction by Human/Rhesus Monkey Trim5␣ SPRY Domain Chimeras Structures of the chimeras made are shown on the left; their restriction properties on the right. The amino acid positions corresponding to the human Trim5␣ protein are indicated above the constructs, whereas the ones corresponding to the rhesus sequence are indicated at the bottom. Restriction assays were performed and scored as described for Figure 1. these positions contributed to the restriction and that they might be degenerate. Hence, the loss of one of these positions could be compensated by the presence of the other. The corresponding change from proline to arginine in position 334 in the rhesus protein did not confer restriction to N L117H, suggesting that unlike HIV-1 restriction, this position was not involved in MLV restriction. Exchanging the eight residues at position 337 of the rhesus protein with the six found in humans resulted in the restriction of N L117H, suggesting that the position was involved in MLV restriction. However, corresponding changes in the human sequence to the rhesus residues did not abolish restriction of N L117H. This suggests that other regions in the human Trim5␣ might be contributing degenerately to restricting N L117H. Indeed, this could involve sequences found in the coiled-coil region (Figure 1B). We have previously shown that Trim1 could also restrict N-MLV, but not HIV-1 [13]. Like Trim5, it consists of an SPRY domain after the tripartite RBCC motif. Trim1 was first identified as a homolog of the Opitz syndrome gene Trim18 (Mid1) to which it showed 83% similarity at the amino acid level [17, 18]. However Trim18, unlike Trim1, does not restrict N-MLV (Figure 4). Formally, this might be due to a lack of protein expression. Alternatively, it provides a further opportunity to examine the role of the SPRY domain in retroviral restriction. The C-terminal portion of African green monkey Trim1, Figure 3. Effects of Specific Amino Acid Changes within the SPRY Domain on the Restriction of HIV-1 and MLV The altered sequences of the SPRY domain are shown on the left, whereas the restriction properties of the variants are shown on the right. Restriction assays were performed and scored as described for Figure 1. Current Biology 76 Figure 4. Restriction of MLV by African Green Monkey Trim1/Trim18 Chimeras Residues 314 to 715 of Vero Trim1 were replaced with residues 314 to 667 of Vero Trim18 to form Vero Trim1/Trim18. Vero Trim18/Trim1 was constructed by replacing residues 314 to 667 of Vero Trim18 with residues 314 to 715 of Vero Trim1. Trim1 sequences are represented by white boxes, whereas Trim18 sequences are colored gray on the left. The ability to restrict is shown in the table on the right. Restriction assays were performed and scored as described for Figure 1. cloned from Vero cells, was used to replace the corresponding region in Trim18 beginning from residue 314 (Figure 4). This chimera (Vero Trim18/Trim1) was active against N-MLV as well as N-MLV containing the N82D mutation, but not B-MLV, a restriction phenotype typical of Trim1. The reciprocal chimera Vero Trim1/Trim18 did not restrict N-MLV and N N82D. Taken together, these results provided further evidence for the involvement of the SPRY domain as the recognition domain in the process of retroviral restriction. In owl monkeys, a psuedogene encoding the cyclophilin A (CypA) protein is inserted within the seventh intron of the Trim5␣ gene [19, 20]. It is thought that the cyclosporine A (CsA) sensitive Lv1 activity of OMK cells [21] can be explained by the presence of a fusion protein in which most of the Trim5␣ SPRY domain is replaced by CypA [19, 20]. This implies that the function provided by SPRY can be replaced by CypA; because CypA is known to bind HIV-1 CA in a CsA sensitive manner [22], this suggests that SPRY is providing binding as well as specificity functions. To examine this idea further, we replaced the SPRY domain of human and rhesus Trim5␣s with the CypA insertion of the OMK Trim5␣ gene (Figure 5). Both novel chimeras restricted wild-type HIV-1 (Figure 5), but not a mutant form that is not bound by CypA, CA G89V. In addition, the substitutions abolished activity against N-MLV, which does not bind CypA [22]. Replacing the SPRY domain of Trim5␣ from African green monkey yielded identical results (data not shown). This confirmed that CypA was performing the function of CA recognition and binding in OMK Trim5-CypA. We have previously found that an artificially created OMK Trim5␣ (V6), which contained the owl monkey SPRY domain in place of CypA, was inactive against both HIV-1 and MLV [20]. To further probe the role of SPRY in CA recognition and binding, we asked if the SPRY domain from rhesus Trim5␣ could rescue the restriction activity of OMK Trim5 V6. Replacing the SPRY of OMK Trim5 V6 in OMKRhSPRY with that from rhesus restored its activity against HIV-1 (Figure 5), suggesting that this domain alone could account for the targeting of HIV-1 CA. However, OMKRhSPRY was not active against N-MLV. Intriguingly, the sequence of the coiledcoil domain of OMK Trim5 is very different from that of the rhesus protein. In the light of the earlier result (Figure 1) suggesting the involvement of the coiled-coil domain in MLV restriction, the failure of OMKRhSPRY to restrict might be due to the lack of MLV CA recognition sequences in the coiled-coil domain of OMK Trim5. Our results indicate that the SPRY domain is the primary determinant of restriction specificity and suggest that this specificity might be mediated through SPRY binding of capsid. The SPRY domain was originally characterized in proteins that regulated intracellular signaling [23]. It can occur as a subdomain within the B30.2 domain [24], which is also found in Trim1 and Trim18 [17, 18]. Although often associated with Trim proteins, SPRY domains are also found in unrelated proteins such as venom toxins and proteins with immunoglobulin-like folds [24]. The functions of B30.2 and SPRY domains are still not known but might include specific proteinprotein interactions. Our studies suggest that in Trim proteins, they play a crucial role in the interaction with infectious agents such as retroviruses to abolish infecFigure 5. Restriction Properties of Chimeric Human, Rhesus, and Owl Monkey Trim5␣ A schematic representation of the chimeras is presented on the left, whereas their ability to restrict HIV-1 and MLV is shown on the right. HuCypA and RhCypA were constructed by replacing the human SPRY domain (residues 295 to 493) or rhesus SPRY domain (residues 297 to 497) with the cyclophilin A insertion in OMK Trim5 V4 (residues 296 to 474). OMKRhSRPY was made by replacing the OMK SPRY of V6 (residues 296 to 495) with the rhesus SPRY domain (residues 297 to 497). Human sequences are represented by white boxes, whereas rhesus and owl monkey sequences are represented by light and dark gray boxes, respectively. Restriction assays were performed and scored as described for Figure 1. Mapping Restriction Specificity Domains of Trim5␣ 77 tion. The SPRY domain seems to function independently from the tripartite motif, because it can be exchanged between different primate Trim5s and can even be replaced with CypA, with associated changes in restriction specificity. It is tempting to speculate that these proteins, together with the other members of the Trim family, might be involved in innate immunity against different infectious agents. Interestingly, mutations in the B30.2 domain in Trim18 (Mid1) [25, 26] and Trim20 (Pyrin) [27, 28] are associated with genetic defects. It has been suggested [25–28] that these mutations are preserved during evolution, because they offer enhanced protection against disease-causing organisms in a manner analogous to the survival of the sickle cell alleles of haemoglobin [29]. The region imparting restriction specificity appears to lie between amino acids 332–340 of human Trim5␣. This is the region showing greatest diversity between the sequences of human, rhesus, and African green monkey [11–13], and we are currently carrying out a detailed phylogenetic analysis of this region to correlate sequence variation with changes in restriction function. Our most provocative initial finding is the observation that a single amino acid substitution (R332P) in the human Trim5␣ protein can confer the ability to restrict HIV-1 replication. Although we do not know the sequence of the ancestral Trim5, we note that rhesus and African green monkey sequences contain a proline at this position, and these species are resistant to HIV-1. This implies that during evolution, changes at this position affecting sensitivity to HIV-1 infection may have occurred. Without such changes it seems unlikely that the cross-species infection(s) leading to the current AIDS epidemic [30] would have occurred. In this context, it is tempting to speculate that the R332P and/or RYQTFV335LFTFPSLT derivatives of human Trim5␣ might be suitable candidate(s) for gene therapy of HIV-1 infection, perhaps introduced in combination with agents that target other steps in the viral life cycle such as the Vif-insensitive, K128D derivative of human APOBEC3G [31–33] or HIV-1 siRNA [34]. Acknowledgments This work was supported by the UK Medical Research Council. Received: October 19, 2004 Revised: November 4, 2004 Accepted: November 5, 2004 Published: January 11, 2005 References 1. Bieniasz, P.D. (2003). Restriction factors: a defense against retroviral infection. Trends Microbiol. 11, 286–291. 2. Goff, S.P. (2004). Genetic control of retrovirus susceptibility in mammalian cells. Annu. Rev. Genet. 38, 61–85. Published online May 26, 2004. 10.1146/annurev.genet.38.072902.094136 3. Stoye, J.P. (2002). An intracellular block to primate lentivirus replication. Proc. Natl. Acad. Sci. USA 99, 11549–11551. 4. Lilly, F. (1970). Fv-2: identification and location of a second gene governing the spleen focus response to Friend leukemia virus in mice. J. Natl. Cancer Inst. 45, 163–169. 5. Best, S., Le Tissier, P., Towers, G., and Stoye, J.P. (1996). Positional cloning of the mouse retrovirus restriction gene Fv1. Nature 382, 826–829. 6. Stevens, A., Bock, M., Ellis, S., LeTissier, P., Bishop, K.N., Yap, M.W., Taylor, W., and Stoye, J.P. (2004). Retroviral capsid determinants of Fv1 NB and NR tropism. J. Virol. 78, 9592–9598. 7. Towers, G., Bock, M., Martin, S., Takeuchi, Y., Stoye, J.P., and Danos, O. (2000). A conserved mechanism of retrovirus restriction in mammals. Proc. Natl. Acad. Sci. USA 97, 12295–12299. 8. Cowan, S., Hatziioannou, T., Cunningham, T., Muesing, M.A., Gottlinger, H.G., and Bieniasz, P.D. (2002). Cellular inhibitors with Fv1-like activity restrict human and simian immunodeficiency virus tropism. Proc. Natl. Acad. Sci. USA 99, 11914– 11919. 9. Besnier, C., Takeuchi, Y., and Towers, G. (2002). Restriction of lentivirus in monkeys. Proc. Natl. Acad. Sci. USA 99, 11920– 11925. 10. Stremlau, M., Owens, C.M., Perron, M.J., Kiessling, M., Autissier, P., and Sodroski, J. (2004). The cytoplasmic body component TRIM5alpha restricts HIV-1 infection in Old World monkeys. Nature 427, 848–853. 11. Hatziioannou, T., Perez-Caballero, D., Yang, A., Cowan, S., and Bieniasz, P.D. (2004). Retrovirus resistance factors Ref1 and Lv1 are species-specific variants of TRIM5alpha. Proc. Natl. Acad. Sci. USA 101, 10774–10779. 12. Keckesova, Z., Ylinen, L.M., and Towers, G.J. (2004). The human and African green monkey TRIM5alpha genes encode Ref1 and Lv1 retroviral restriction factor activities. Proc. Natl. Acad. Sci. USA 101, 10780–10785. 13. Yap, M.W., Nisole, S., Lynch, C., and Stoye, J.P. (2004). Trim5alpha protein restricts both HIV-1 and murine leukemia virus. Proc. Natl. Acad. Sci. USA 101, 10786–10791. 14. Perron, M.J., Stremlau, M., Song, B., Ulm, W., Mulligan, R.C., and Sodroski, J. (2004). TRIM5alpha mediates the postentry block to N-tropic murine leukemia viruses in human cells. Proc. Natl. Acad. Sci. USA 101, 11827–11832. 15. Reymond, A., Meroni, G., Fantozzi, A., Merla, G., Cairo, S., Luzi, L., Riganelli, D., Zanaria, E., Messali, S., Cainarca, S., et al. (2001). The tripartite motif family identifies cell compartments. EMBO J. 20, 2140–2151. 16. Bock, M., Bishop, K.N., Towers, G., and Stoye, J.P. (2000). Use of a transient assay for studying the genetic determinants of Fv1 restriction. J. Virol. 74, 7422–7430. 17. Buchner, G., Montini, E., Andolfi, G., Quaderi, N., Cainarca, S., Messali, S., Bassi, M.T., Ballabio, A., Meroni, G., and Franco, B. (1999). MID2, a homologue of the Opitz syndrome gene MID1: similarities in subcellular localization and differences in expression during development. Hum. Mol. Genet. 8, 1397–1407. 18. Perry, J., Short, K.M., Romer, J.T., Swift, S., Cox, T.C., and Ashworth, A. (1999). FXY2/MID2, a gene related to the X-linked Opitz syndrome gene FXY/MID1, maps to Xq22 and encodes a FNIII domain-containing protein that associates with microtubules. Genomics 62, 385–394. 19. Sayah, D.M., Sokolskaja, E., Berthoux, L., and Luban, J. (2004). Cyclophilin A retrotransposition into TRIM5 explains owl monkey resistance to HIV-1. Nature 430, 569–573. 20. Nisole, S., Lynch, C., Stoye, J.P., and Yap, M.W. (2004). A Trim5cyclophilin A fusion protein found in owl monkey kidney cells can restrict HIV-1. Proc. Natl. Acad. Sci. USA 101, 13324–13328. 21. Towers, G.J., Hatziioannou, T., Cowan, S., Goff, S.P., Luban, J., and Bieniasz, P.D. (2003). Cyclophilin A modulates the sensitivity of HIV-1 to host restriction factors. Nat. Med. 9, 1138–1143. 22. Luban, J., Bossolt, K.L., Franke, E.K., Kalpana, G.V., and Goff, S.P. (1993). Human immunodeficiency virus type 1 Gag protein binds to cyclophilins A and B. Cell 73, 1067–1078. 23. Ponting, C., Schultz, J., and Bork, P. (1997). SPRY domains in ryanodine receptors (Ca(2⫹)-release channels). Trends Biochem. Sci. 22, 193–194. 24. Seto, M.H., Liu, H.L., Zajchowski, D.A., and Whitlow, M. (1999). Protein fold analysis of the B30.2-like domain. Proteins 35, 235–249. 25. Quaderi, N.A., Schweiger, S., Gaudenz, K., Franco, B., Rugarli, E.I., Berger, W., Feldman, G.J., Volta, M., Andolfi, G., Gilgenkrantz, S., et al. (1997). Opitz G/BBB syndrome, a defect of midline development, is due to mutations in a new RING finger gene on Xp22. Nat. Genet. 17, 285–291. 26. Gaudenz, K., Roessler, E., Quaderi, N., Franco, B., Feldman, G., Current Biology 78 27. 28. 29. 30. 31. 32. 33. 34. Gasser, D.L., Wittwer, B., Horst, J., Montini, E., Opitz, J.M., et al. (1998). Opitz G/BBB syndrome in Xp22: mutations in the MID1 gene cluster in the carboxy-terminal domain. Am. J. Hum. Genet. 63, 703–710. Consortium, T.F.F. (1997). A candidate gene for familial Mediterranean fever. Nat. Genet. 17, 25–31. Consortium, T.I.F. (1997). Ancient missense mutations in a new member of the RoRet gene family are likely to cause familial Mediterranean fever. Cell 90, 797–807. Ashley-Koch, A., Yang, Q., and Olney, R.S. (2000). Sickle hemoglobin (HbS) allele and sickle cell disease: a HuGE review. Am. J. Epidemiol. 151, 839–845. Gao, F., Bailes, E., Robertson, D.L., Chen, Y., Rodenburg, C.M., Michael, S.F., Cummins, L.B., Arthur, L.O., Peeters, M., Shaw, G.M., et al. (1999). Origin of HIV-1 in the chimpanzee Pan troglodytes troglodytes. Nature 397, 436–441. Bogerd, H.P., Doehle, B.P., Wiegand, H.L., and Cullen, B.R. (2004). A single amino acid difference in the host APOBEC3G protein controls the primate species specificity of HIV type 1 virion infectivity factor. Proc. Natl. Acad. Sci. USA 101, 3770– 3774. Schrofelbauer, B., Chen, D., and Landau, N.R. (2004). A single amino acid of APOBEC3G controls its species-specific interaction with virion infectivity factor (Vif). Proc. Natl. Acad. Sci. USA 101, 3927–3932. Xu, H., Svarovskaia, E.S., Barr, R., Zhang, Y., Khan, M.A., Strebel, K., and Pathak, V.K. (2004). A single amino acid substitution in human APOBEC3G antiretroviral enzyme confers resistance to HIV-1 virion infectivity factor-induced depletion. Proc. Natl. Acad. Sci. USA 101, 5652–5657. Humeau, L.M., Binder, G.K., Lu, X., Slepushkin, V., Merling, R., Echeagaray, P., Pereira, M., Slepushkina, T., Barnett, S., Dropulic, L.K., et al. (2004). Efficient lentiviral vector-mediated control of HIV-1 replication in CD4 lymphocytes from diverse HIV⫹ infected patients grouped according to CD4 count and viral load. Mol. Ther. 9, 902–913.