Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Tissue engineering wikipedia , lookup
Extracellular matrix wikipedia , lookup
Cell encapsulation wikipedia , lookup
Cytokinesis wikipedia , lookup
Organ-on-a-chip wikipedia , lookup
Cell growth wikipedia , lookup
Cell culture wikipedia , lookup
List of types of proteins wikipedia , lookup
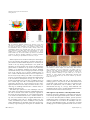
BMB reports Fluridone affects quiescent centre division in the Arabidopsis thaliana root stem cell niche 1 2 1, Woong Han , Hanma Zhang & Myeong-Hyeon Wang * 1 Department of Medical Biotechnology, College of Biomedical Science, Kangwon National University, Chuncheon 200-071, Korea, 2Centre for Plant Sciences, Institute of Integrative and Comparative Biology (IICB), Faculty of Biological Sciences, University of Leeds, Leeds, LS2 9JT, UK Plants undergo cell division throughout their life in order to maintain their growth. It is well known that root and shoot tip of plants possess meristems, which contain quiescent cells. Fluridone (1-methyl-3-phenyl-5-(3-trifluromethyl (phenyl))-4-(1H)pyridinone) is an established inhibitor of both ABA and carotenoid biosynthesis. However, the other functions of fluridone remain undiscovered. In this report, we provide experimental evidence that fluridone plays a role in the division of the quiescent centre of the Arabidopsis root meristem. This study examined the effects of exogenous fluridone and ABA on the development of the stem cell niche in Arabidopsis root. We show that fluridone promoted the division of stem cells in the quiescent centre, whereas exogenous ABA suppressed quiescent centre division. Furthermore, we established a novel regulatory function for fluridone by demonstrating that it plays an important role in postembryonic development. [BMB reports 2010; 43(12): 813-817] root stem cell niche (10, 11). Endogenous and exogenous ethylene-mediated signals control QC division during the postembryonic stage of the plant life cycle (11). Furthermore, abscisic acid (ABA) plays a key role in the developmental status of the plant over the entire life span, especially during seed development and dormancy, and it responds to diverse environmental stresses (12-14). ABA has been established as an inhibitor of root growth based on its effects on root elongation and lateral root initiation. However, few studies have reported enhanced root growth in the presence of low concentrations of ABA (15-17). It was reported earlier that fluridone is an inhibitor of phytoene desaturase, which converts phytoene to carotene (18, 19). Considering the above, this study examined the effects of exogenous fluridone and ABA on the development of the Arabidopsis root stem cell niche and investigated in detail whether or not fluridone promotes QC division in the Arabidopsis root meristem accompanied by extra columella cell layers. INTRODUCTION RESULTS AND DISCUSSION In general, the dividing cells of plants are located and distributed in regions of the roots and shoots known as meristems (1). The Arabidopsis thaliana root meristem consists of a quiescent centre (QC) and initial cells. The QC acts as an organizing centre, whereas initial cells serve as stem cells (2-4) for all tissues and promote growth in the roots (5). Ablation studies have demonstrated that QC prevents the differentiation of columella initial cells (3). To date, researchers have determined that QC plays a primary role in meristem maintenance in Arabidopsis root and that these QC are associated with the SCR, SHR, and WOX5 genes (6-9). Plant hormones are associated with QC division and maintain meristem activity in the Fluridone promotes QC division in the Arabidopsis root meristem *Corresponding author. Tel: 82-33-250-6486; Fax: 82-33-241-6480; E-mail: [email protected] DOI 10.5483/BMBRep.2010.43.12.813 Received 4 October 2010, Accepted 10 November 2010 Keywords: Abscisic acid (ABA), Fluridone, Quiescent centre (QC), Stem cell niche http://bmbreports.org The central part of plant root stem cells contains a QC surrounded by initial cells, which in total are referred to as the meristem (Fig. 1a). The QC rarely divides under normal conditions, but initial cells are able to divide (e.g., for plant growth) throughout their life span. To determine the effect of fluridone in Arabidopsis, we germinated Col-0 wild-type in PANG2 medium for 3 days and then transferred it to PANG2 medium containing fluridone (Flu+). Seedlings of Arabidopsis roots grown in PANG2 medium (Fig. 1b) did not show QC division even after 10 days of germination, whereas it was clearly observed in seedlings germinated in Flu+ medium (Fig. 1c, white arrows). Transverse section of germinated seedling showed a four-celled QC in longitudinal view. In addition, we also determined the QC division ratio of seedlings incubated in 10 μM fluridone-supplemented medium for 10 DAG. QC division (60%) was observed in the roots of these seedlings, whereas wild-type did not show any such division (n = 50) (Fig. 1d). BMB reports 813 Fluridone promotes quiescent centre division Woong Han, et al. Fig. 1. Exogenous fluridone enhances QC division in the stem cell niche of Arabidopsis roots. (a) Schematic representation of the stem cell niche of Arabidopsis root meristems. Co/En, cortical and endodermis initial; P, pericycle initials; LRC/Ep, lateral root cap/epidermis initial; CSC, columella stem cells. (b, c) Col-0 seedlings cultured for 10 DAG (Days After Germination) on control medium (b) or medium containing 10 μM fluridone (c). Arrow head indicates QC and arrow indicates QC division. (d) Fluridone promotes QC division. Seedlings cultured for 10 DAG on PANG2 alone or containing 10 μM fluridone. Scale bars, 50 μm. WOX5 expression was initiated in embryonic cells and gave rise to QC formation. This finding is consistent with other reports that also found that WOX5 expression is consistently observed in the QC during postembryonic root growth (8). Therefore, we used a WOX5p::GFP construct to determine the general effects of fluridone on QC division and specifically whether or not fluridone stimulates QC division. Seedlings showed that WOX5p::GFP was strongly expressed in the single layer of QC cells (Fig. 2a). In contrast, seedlings grown on Flu+ medium indicated that WOX5p::GFP was expressed in the extra layer of QC cells (Fig. 2b). SCR is a transcription factor that is accumulated in the QC and has been demonstrated to be essential for development (20) (Fig. 2c). SCR was detected in the double layer of QC cells in seedlings grown on Flu+ medium (Fig. 2d). SHR is a transcription factor usually expressed in the stele (21), and its expression was also investigated and observed in our study on PANG2 and Flu+2 medium (Fig. 2e). Seedlings cultured in Flu+ medium clearly revealed QC division (Fig. 2f). ABA affects QC quiescence in the Arabidopsis root meristem, which in turn affects QC division. Differentiated columella cells accumulate starch within amyloplast organelles in the root tip for root gravitropism (22, 23). To determine whether or not the extra cells that form in Flu+ medium function as QC cells, we tested their ability to regulate differentiation in the surrounding initial cells (4). Ablation studies have previously revealed that QC cells in Arabidopsis roots promote earlier differentiation of columella initial cells and have detected (by Lugol staining) the starch grains of columella cells 814 BMB reports Fig. 2. Fluridone promotes QC division in the Arabidopsis thaliana root meristem. Confocal images of 10 DAG (Days After Germination) of WOX5p:: GFP (a, b), SCRp::GFP (c, d), SHRp:: GFP (e, f) seedlings, with their structures displayed (propidium iodide staining). g-i: Col-0 seedlings cultured for 10 DAG on Control medium (g), medium containing 0.5 μM of ABA (h), or 10 μM of fluridone (i). Lugol staining marks differentiated columella cells (g-i). Arrow head indicates QC and arrow indicates QC division. Scale bars, 20 μm. instead of columella initial cells (24). In the present study, seedlings grown in the presence of ABA and fluridone and stained with Lugol revealed a single layer of unstained cells below the supernumerary cells in PANG2, ABA, and fluridone-treated roots (Fig. 2e, f and g). These additional cells produced in the QC in Flu+ medium displayed both functions and characteristics similar to those of the QC. ABA suppresses QC division in a dose-dependent manner Exogenous exposure to fluridone is expected to promote division in the QC during plant development. However, fluridone, as a common ABA biosynthesis inhibitor, presents questions regarding the ability of exogenous ABA to suppress QC division and whether or not it affects the Arabidopsis root meristem in a dose-dependent manner. To confirm this, we tested seedlings grown in media supplemented with various concenhttp://bmbreports.org Fluridone promotes quiescent centre division Woong Han, et al. Fig. 3. ABA suppresses QC division. (a) ABA suppresses QC division in a dose-dependent manner. Seedlings cultured for 10 DAG on PANG2 medium containing different concentrations of ABA and 10 μM fluridone. (b) ABA-deficient mutant displays QC division. Seedlings cultured for 10 DAG on PANG2 medium. Error bars, standard deviations. Fig. 4. Fluridone results in production of extra columella cell layers. (a) Confocal laser microscopic images of columella cells. (b) Number of columella cell layers containing different concentrations of fluridone. All seedlings were cultured for 10 DAG. PANG2, control medium; Flu, control medium containing 10 μM fluridone. Error bars, standard deviations. Scale bars, 50 μm. trations of ABA and predicted a 10% decrease in the QC division ratio with 5 nM (least concentration) ABA (Fig. 3a). Interestingly, the QC division ratio slightly decreased with increased ABA concentrations, and there was 0% QC division at 5,000 nM ABA. Therefore, it was concluded that ABA suppressed QC in a dose-dependent manner. There is a growing hypothesis that fluridone might also participate in other pathways that control QC division and not really act as an inhibitor of ABA. Therefore, we investigated whether or not QC division could be controlled in ABA-deficient mutants (e.g., aba1-1, aba3-2) (25) as well as any other reasons for this phenomenon. If the QC division phenotype of fluridone was caused by the absence of ABA, then ABA-deficient mutants would be characterized by increased supernumerary divisions in the QC. We observed extra QC cell division in ABA-deficient mutants grown in PANG2 medium without fluridone supplementation (Fig. 3b), which suggests that ABA and QC division were associated. as a measure of cell division in the QC. We observed that the seedlings grown in Flu+ medium had more layers of columella cells than those grown in basic medium (Fig. 4a). This additional division in Flu+ medium resulted in increased development of columella layers compared to the seedlings grown in PANG2 (Fig. 4b). Plants grown in media without fluridone showed 4.83 ± 0.37 (SD) layers, whereas seedlings cultured in the presence of fluridone (1 μM of flu) displayed 5.3 ± 0.59 (SD) layers. The results shown here suggest that cell division in the QC was regulated by both fluridone and ABA. Our results reveal that fluridone modulated QC division and enhanced columella cell layer formation, whereas exogenous ABA suppressed QC division. The results also suggest that fluridone and ABA were involved in Arabidopsis root meristem maintenance. Fluridone changes cellular pattern of the stem cell niche Seeds of Arabidopsis thaliana ecotype Col-0, Ler-0, ABA-deficient mutants, aba1-1, and aba3-2 (25) were used in the experiments. SHRp::GFP and SCRp::GFP were obtained from Philip N. Benfey (Duke University), and WOX5p::GFP from Ben Scheres and Renze Heidstra (University of Utrecht). Arabidopsis seeds were surface-sterilized with 80% ethanol and 1% active NaClO for 10 min, followed by three washes with 100% ethanol. The seeds were dried in a flow hood and placed on the surface of agar-solidified medium. All media (PANG2) used in this study was of the following basic composition: 1.5 mM CaCl2, 1.0 mM NaH2PO4, 0.2 mM MgSO4, 0.4 mM KNO3, 1.8 μM KI, 20 μM H3BO3, 3.0 μM ZnSO4, 0.06 We also investigated fluridone-induced cell division of the QC in Arabidopsis root and also whether or not such cell division increases the number of columella cell layers. Plants grown in the presence of aminoethoxyvinylglycine (AVG) display inhibited ethylene synthesis and reportedly develop fewer columella layers (11). To identify the effects of fluridone on columella cell layers, we monitored the Arabidopsis root stem cell niche. We hypothesised that the addition of fluridone would induce additional QC division, which could affect the number of columella cell layers. Therefore, we counted the number of columella cell layers in Flu+ and PANG2 medium http://bmbreports.org MATERIALS AND METHODS Plant materials and growth conditions BMB reports 815 Fluridone promotes quiescent centre division Woong Han, et al. μM CuSO4, 0.4 μM Na2MoO4, 4.0 μM CoCl2, 0.04% (v/v) Sigma’s Ferrous sulfate chelate solution (F-0518), 0.5% (w/v) 2-(N-morpholino) ethanesulphonic acid (MES), 0.5% (w/v) sucrose, and 10% (w/v) agar-agar (A-1080, Fisher Chemicals). The pH levels of all media were adjusted to 5.7 using 1.0 M o KOH, followed by autoclaving at 121 C for 20 min. Fluridone (Sigma 59756-60-4) and ABA were added from filter-sterilized o stock solutions after autoclaving at 50 C. Stock solutions of fluridone (10 mM) and ABA (1 mM) were made in 100% ethanol. Seedlings were grown on the surfaces of these agar media in vertically-oriented Petri dishes. Dishes were then sealed with Parafilm, with three openings of 0.3-0.5 cm to allow air exchange during growth, and kept in a growth room at o 22 C under a 16-h/8-h light/dark cycle using overhead lighting. Growth conditions in the dark were identical except for the absence of light. Confocal microscopy Whole seedlings were stained with 10 μg ml-1 of propidium iodide (PI; Sigma), mounted in water under glass cover slips, and then analyzed for PI using a confocal laser scanning microscope (Carl Zeiss LSM 510 META NLO, Germany). 7. 8. 9. 10. 11. Starch staining For starch staining of roots, tissues were incubated in Lugol’s solution (Sigma) for 5 to 10 min and then mounted in 20% glycerol for microscopic analysis. Acknowledgements We thank the Korea Basic Science Institute (Chuncheon Center) for assistance with the confocal equipment. This work was partially supported by a grant from the Institute of Biosciences and Biotechnology, Kangwon National University, Chuncheon, South Korea. REFERENCES 1. Dolan, L., Janmaat, K., Willemsen, V., Linstead, P., Poethig, S., Roberts, K. and Scheres, B. (1993) Cellular organization of the Arabidopsis root. Development 119, 71-84. 2. Wildwater, M., Campilho, A., Perez-Perez, J. M., Heidstra, R., Blilou, I., Korthout, H., Chatterjee, J., Mariconti, L., Gruissem, W. and Scheres, B. (2005) The RETINOBLASTOMARELATED gene regulates stem cell maintenance in Arabidopsis roots. Cell 29, 1337-1349. 3. van den Berg, C., Willemsen, V., Hendriks, G., Weisbeek, P. and Scheres, B. (1997) Short-range control of cell differentiation in the Arabidopsis root meristem. Nature 390, 287-289. 4. Kidner, C., Sundaresan, V., Roberts, K. and Dolan, L. (2000) Clonal analysis of the Arabidopsis root confirms that position, not lineage, determines cell fate. Planta 211, 191-199. 5. Stahl, Y. and Simon. R. (2005) Plant stem cell niches. Int. J. Dev. Biol. 49, 479-489. 6. Benfey, P. N., Linstead, P. J., Roberts, K., Schiefelbein, J. 816 BMB reports 12. 13. 14. 15. 16. 17. 18. 19. 20. 21. W., Hauser, M. T. and Aeschbacher. R. A. (1993) Root development in Arabidopsis: four mutants with dramatically altered root morphogenesis. Development 119, 57-70. Scheres, B., Laurenzio, L. D., Willemsen, V., Hauser, M. T., Janmaat, K., Weisbeek, P. and Benfey. P. N. (1995) Mutations affecting the radial organization of the Arabidopsis root display specific defects throughout the radial axis. Development 121, 53-62. Haecker, A., Gross-Hardt, R., Geiges, B., Sarkar, A., Breuninger, H., Herrmann, M. and Laux, T. (2004) Expression dynamics of WOX genes mark cell fate decisions during early embryonic patterning in Arabidopsis thaliana. Development 131, 657-668. Sarkar, A. K., Luijten, M., Miyashima, S., Lenhard, M., Hashimoto, T., Nakajima, K., Scheres, B., Heidstra, R. and Laux, T. (2007) Conserved factors regulate signalling in Arabidopsis thaliana shoot and root stem cell organisers. Nature 446, 811-814. Thomann, A., Lechner, E., Hansen, M., Dumbliauskas, E., Parmentier, Y., Kieber, J., Scheres, B. and Genschik. P. (2009) Arabidopsis CULLIN3 genes regulate primary root growth and patterning by ethylene-dependent and -independent mechanisms. PLoS Genet. 5, doi:10.1371/journal. pgen.1000328. Ortega-Martínez, O., Pernas, M., Carol, R. J. and Dolan, L. (2007) Ethylene modulates stem cell division in the Arabidopsis thaliana root. Science 317, 507-510. Seo, M. and Koshiba. T. (2002) Complex regulation of ABA biosynthesis in plants. Trends Plant Sci. 7, 41-48. Chinnusamy, V., Gong, Z. and Zhu, J. K. (2008) Abscisic acid-mediated epigenetic processes in plant development and stress responses. J. Integr. Plant Biol. 50, 1187-1195. Wan, X. R. and Li, L. (2006) Regulation of ABA level and water-stress tolerance of Arabidopsis by ectopic expression of a peanut 9-cis-epoxycarotenoid dioxygenase gene. Biochem. Biophys. Res. Commun. 347, 1030-1038. De Smet, I., Signora, L., Beeckman, T., Inzé, D., Foyer, C. H. and Zhang, H. (2003) An abscisic acid-sensitive checkpoint in lateral root development of Arabidopsis. The Plant J. 33, 543-555. Fukaki, H., Okushima, Y. and Tasaka, M. (2007) Auxinmediated lateral root formation in higher plants. Int. Rev. Cytol. 256, 111-137. Fukaki, H. and Tasaka. M. (2009) Hormone interactions during lateral root formation. Plant Mol. Biol. 69, 437449. Bartels, P. G. and Watson, C. W. (1978) Inhibition of carotenoid synthesis by fluridone and norflurazon. Weed Sci. 26, 198-203. Fong, F. J. and Schiff, A. (1979) Blue-light-induced absorbance changes associated with carrotenoids in Euglena. Planta 146, 119-127. Di Laurenzio, L., Wysockadiller, J., Malamy, J. E., Pysh, L., Helariutta, Y., Freshour, G., Hahn, M. G., Feldmann, K. A. and Benfey, P. N. (1996) The SCARECROW gene regulates an asymmetric cell division that is essential for generating the radial organization of the Arabidopsis root. Cell 86, 423-433. Nakajima, K., Sena, G., Nawy, T. and Benfey, P. N. (2001) Intercellular movement of the putative transcription factor http://bmbreports.org Fluridone promotes quiescent centre division Woong Han, et al. SHR in root patterning. Nature 413, 307-311. 22. Sack, F. D. (1997) Plastids and gravitropic sensing. Planta 203, S63-S68 23. Blancaflor, E. B., Fasano, J. M. and Gilroy, S. (1998) Mapping the functional roles of cap cells in the response of Arabidopsis primary roots to gravity. Plant Physiol. 116, 213-222. 24. van den Berg, C., Weisbeek, P. and Scheres, B. (1998) http://bmbreports.org Cell fate and cell differentiation status in the Arabidopsis root. Planta 205, 483-491. 25. Koornneef, M., Jorna, M. L., Brinkhorst-van der Swan, D. L. C. and Karssen, C. M. (1982) The isolation of abscisic acid (ABA) deficient mutants by selection of induced revertants in non-germinating gibberellin sensitive lines of Arabidopsis thaliana (L.) heynh. Theor. Appl. Genet. 61, 385-393. BMB reports 817