Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
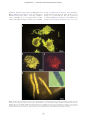
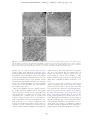
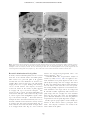
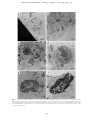
JOURNAL OF PLANKTON RESEARCH j VOLUME 33 j NUMBER 10 j PAGES 1538 – 1549 j 2011 The ultrastructure of a doliolid and a copepod fecal pellet MARION KÖSTER 1*, RABEA SIETMANN 2, ANNETTE MEUCHE 2 AND GUSTAV-ADOLF PAFFENHÖFER 3 1 6, 18565 INSEL HIDDENSEE, GERMANY, 2ERNST-MORITZ3 ARNDT-UNIVERSITÄT GREIFSWALD, INSTITUT FÜR MIKROBIOLOGIE, FRIEDRICH-LUDWIG-JAHN-STR. 15 A, 17487 GREIFSWALD, GERMANY AND SKIDAWAY INSTITUTE OF OCEANOGRAPHY, 10 OCEAN SCIENCE CIRCLE, SAVANNAH, GA 31411, USA ERNST-MORITZ-ARNDT-UNIVERSITÄT GREIFSWALD, MIKROBIELLE ÖKOLOGIE, SCHWEDENHAGEN *CORRESPONDING AUTHOR: [email protected] Received January 27, 2011; accepted in principle May 18, 2011; accepted for publication May 23, 2011 Corresponding editor: Roger Harris The goal of this study was to determine the morphology and ultrastructure of doliolid pellets using light, epifluorescence and transmission electron microscopy and compare the results to observations of calanoid copepod pellets. For (ultra)structural analyses, pellets of gonozooids of Dolioletta gegenbauri and females of the copepod Eucalanus pileatus were produced in feeding experiments at close to environmental food concentrations. Thin sections of a representative doliolid pellet revealed that these pellets were mainly composed of intact diatom valves, a few fragmented valves and intact flagellates. While the larger diatoms, Rhizosolenia alata, were completely digested (empty valves), the smaller diatoms, Thalassiosira weissflogii, were partly, or not digested at all. The phytoflagellate, Isochrysis galbana, appeared to be hardly digested. Aggregations of bacteria occurred mostly inside pellets associated closely with intact I. galbana flagellates and partly digested T. weissflogii cells; some scattered bacteria were found among fragmented valves. No, or little, bacterial colonization was associated with empty R. alata valves, and hardly digested T. weissflogii cells. Whereas doliolid fecal pellets were loosely packed and composed of fully, incompletely and/or hardly digested food particles, pellets of the copepod E. pileatus were densely packed and consisted mainly of fragmented diatom valves. Pellets of doliolids and calanoid copepods can represent a high percentage of the particulate organic carbon in the water-column on subtropical continental shelves. KEYWORDS: doliolids; calanoid copepods; fecal pellets; ultrastructure; transmission electron microscopy I N T RO D U C T I O N The occurrence and ecology of doliolids (Tunicata, Thaliacea) have been studied for more than three decades on the US southeastern continental shelf (e.g. Deibel, 1998; Gibson and Paffenhöfer, 2000; Deibel and Paffenhöfer, 2009). Doliolids have exponential growth rates, and high asexual reproduction rates (Paffenhöfer and Gibson, 1999). These energy-consuming processes are fuelled by the uptake of large amounts of phytoplankton. They are able to ingest 20% of the in situ phytoplankton standing stock per day (Paffenhöfer et al., 1995). However, their assimilation efficiency often comprises ,50% of the ingested particulate organic matter. That means that the ingested food particles are only partly digested (Paffenhöfer and Köster, 2005) and unassimilated organic matter is released as fecal pellets. Doliolids produce fecal pellets at high rates (6 – 8 pellets per gonozooid per hour, Paffenhöfer unpubl. observ.). Considering their often high concentrations .1000 zooids m23 (e.g. Paffenhöfer and Lee, 1987; Paffenhöfer et al., 1995) pellet production is ecologically doi:10.1093/plankt/fbr053, available online at www.plankt.oxfordjournals.org. Advance Access publication July 7, 2011 # The Author 2011. Published by Oxford University Press. All rights reserved. For permissions, please email: [email protected] M. KÖSTER ET AL. j ULTRASTRUCTURE OF DOLIOLID FECAL PELLETS significant. Laboratory observations of Paffenhöfer and Köster (Paffenhöfer and Köster, 2011) revealed that a laboratory-reared nurse with a 5-cm long cadophore carries maximally up to 100 trophozooids (for illustration of the different stages in the life cycle of doliolids see Fig. 1 in Paffenhöfer and Gibson, 1999). Each phorozooid when released from the nurse produces on an average 9 – 14 gonozooids per day over a period of 8 – 18 days (Paffenhöfer and Gibson, 1999). Based on an average gonozooid pellet production rate of 6 – 8 per Fig. 1. Freshly produced fecal pellets of gonozooids of Dolioletta gegenbauri and adult female Eucalanus pileatus. Fecal pellets are mainly composed of Thalassiosira weissflogii cells with a few Isochrysis galbana and Rhodomonas sp. cells. (a) Irregularly shaped fecal pellets of doliolids observed in the darkfield, (b and c) Individual doliolid fecal pellet observed in the darkfield and under green excitation light (excitation of 510– 560 nm, dichroic mirror 560 nm, emission of 575– 640 nm). Pronounced red autofluorescence of hardly digested Thalassiosira weissflogii cells is visible. (d) Fecal pellets of Eucalanus pileatus. Insert shows the posterior part of a fecal pellet (white arrow indicates the peritrophic membrane). 1539 JOURNAL OF PLANKTON RESEARCH j VOLUME hour, we calculated between a minimum of 144 and a maximum of 192 pellets per day produced by one gonozooid. Deibel and Paffenhöfer (Deibel and Paffenhöfer, 2009) reported a maximum gonozooid concentration of 1500 m23 on the continental US shelf. That means that daily fecal pellet production can reach between 216 000 and 288 000 pellets m23. These high numbers highlight that doliolid fecal pellets may significantly contribute to the pool of particulate organic carbon in upwelled waters where they provide nutrient-rich microaggregates for microbial colonization. An average gonozooid pellet comprises ca. 400 ng organic carbon (Brandes, pers. comm.). Thus, the water column is supplied with 86– 115 mg C m23 d21 during an intense doliolid bloom. A doliolid bloom with a period of 10 days produces 0.9 –1.2 g of organic carbon m23 enriched in fecal pellets in upwelled waters. Calanoid copepods on average usually produce fecal pellets at lower rates than D. gegenbauri gonozooids. Females of the copepod Eucalanus pileatus produce 2 pellets h21 compared with the 6 – 8 pellets h21 produced by a large gonozooid, both zooplankters being of similar weight i.e. 20– 25 mg C individual21. Younger stages of calanoids can produce maximally as many as 5 pellets h21 (Paffenhöfer and Knowles, 1979). Since previous pellet studies focused mostly on those of calanoid copepods, and since the two species developed well in summer intrusions of upwelled water on the US southeastern continental shelf, we decided to include pellets of one of the species, E. pileatus, for comparative purposes in our study on doliolid pellets. Food concentrations chosen were close to those in upwelled waters on the US southeastern shelf. Our goal was (i) to compare the (ultra)morphology of fecal pellets of doliolids and copepods and their bacterial colonization, and (ii) evaluate the degree of phytoplankton digestion (two diatom species and flagellates) using transmission electron microscopy (TEM). METHOD 33 j NUMBER 10 j PAGES 1538 – 1549 j 2011 Rhodomonas sp (4 and 8 mm ESD, respectively). Pellets were produced in 1.9 L glass jars containing the suspended food at a total concentration of 160 mg C L21 in GF/C filtered seawater. The jars were mounted on a slowly rotating (0.3 rpm) plankton wheel at 218C. Pellets of E. pileatus were produced at a total food concentration of 238 mg C L21. Food particles offered were mainly T. weissflogii and small amounts of flagellates. Gonozooids of D. gegenbauri produced between six and eight pellets per hour, whereas adult female E. pileatus produced two pellets per hour. To obtain a sufficient number of pellets for microscopic analyses, doliolids and copepods were incubated for 2 and 16 h, respectively. Pellet sampling and fixation For microscopic examination, recently produced pellets (age of ,2 and ,16 h, respectively) of D. gegenbauri gonozooids and females of E. pileatus were collected with a cut-off glass pipette and transferred into a 150 mL glass dish filled with GF/C filtered seawater. The integrity of pellets was checked under an Olympus dissecting microscope at 20-fold magnification; only intact pellets were selected for later microscopic investigations. Pellets were rinsed in 0.2-mm filtered seawater. Aliquots of 5 – 10 pellets were fixed with formaldehyde [2%(v/v) final concentration] and examined using light and epifluorescence microscopy. For TEM 10 pellets were transferred into 2 mL of 0.2-mm filtered seawater containing 1%(v/v) glutaraldehyde and 2%(v/v) paraformaldehyde. Pellets were fixed at 218C for 2 h, and then stored at 48C prior to preparation for TEM. Size and volume of pellets The size of pellets was determined under an Olympus dissecting microscope at 20-fold magnification using an ocular micrometer. The average width (w) and length (l) of D. gegenbauri and E. pileatus pellets were measured at +25 mm (n ¼ 150). For pellet volume estimations, following equations were used, Pellet production (i) For pellet production, two feeding experiments were performed in September 2009. Laboratory-reared gonozooids of D. gegenbauri had been feeding on the diatoms Thalassiosira weissflogii (8 – 12 mm ESD, Equivalent Spherical Diameter) and Rhizosolenia alata forma indica (length of 200– 300 mm, width of 12– 16 mm) with small amounts of the flagellate Isochrysis galbana and very few cells of the flagellate (ii) Volume (mm3) of D. gegenbauri pellets ¼ width (mm) length (mm) thickness (mm). Volume (mm3) of E. pileatus pellets ¼ p (width/2)2 (mm2) length (mm). Transmission electron microscopy After embedding the fecal pellets in low-gelling-temperature agarose, they were postfixed in 1%(v/v) osmium tetroxide for 2 h at room temperature (and 48C over 1540 M. KÖSTER ET AL. j ULTRASTRUCTURE OF DOLIOLID FECAL PELLETS night) followed by immersion of the specimen in 2%(v/v) uranyl acetate for 2 h. After dehydration in a graded series of ethanol the material was embedded in EPONTM epoxy resin. Sections were cut on an ultramicrotome (Reichert Ultracut, Leica UK Ltd, Milton Keynes, UK), stained with uranyl acetate and lead citrate and examined with a TEM LEO 906 (Zeiss, Oberkochen, Germany) at 80 kV. The representativeness of chosen doliolid pellets was evaluated by their shape, size, composition, and how intact they were checked under a dissecting microscope at 20-fold magnification. For copepods, the intact nature of the outer peritrophic membrane was an additional criterion to assess how representative they were. According to our previous light microscopic investigations doliolid pellets produced under defined laboratory food concentration and composition conditions revealed a morphological structure and contained food particles of a digestive state as found previously (Paffenhöfer and Köster, 2005). Also, pellets of E. pileatus were similar to previous observations. Therefore, we selected only one representative pellet of each taxon for a detailed ultrastructure analysis by TEM. The selected squarish-like doliolid pellet had a length of 450 mm and the cylindrically-shaped copepod pellet a length of 500 mm and a width of 50– 60 mm. For the estimation of bacterial abundance in doliolid pellets, micrographs of selected thin sections at 800-fold magnification were used. Bacteria and microalgae (non-, partly- and completely-digested) were counted in fields of 1946 mm2. Light and epifluorescence microscopy The morphology of formaldehyde-fixed doliolid and copepod pellets was examined with an Axiophot epifluorescence microscope (Zeiss, Jena, Germany) at 50 and 100-fold magnification, respectively. Chlorophyll autofluorescence was determined under green excitation light (excitation of 510 to 560 nm, dichroic mirror 560 nm, emission of 575 – 640 nm) and used to estimate the degree of digested food particles in pellets. Micrographs were recorded with a high-resolution CCD camera 1300 BC (Vosskühler, Osnabrück, Germany) using an image analysis system (Lucia 4.81). R E S U LT S Light microscopy Light microscopy observations showed that D. gegenbauri fecal pellets produced by gonozooids had an average size of 300 mm by 600 mm (Fig. 1a). Mean pellet volumes usually ranged between 0.03 and 0.05 mm3 depending on the size of the gonozooids and the type of ingestable food particles. The shape of the gonozooid pellets was generally flat and squarish-like, but could also be irregularly shaped and show elongated threadlike structures. The thickness of the pellets varied from monolayered to multilayered cell sheets; the maximum thickness was 150 mm. Especially the thinner and more loosely structured outer regions were translucent, whereas the central regions were more densely compacted. Dolioletta gegenbauri fecal pellets consisted mainly of small centric T. weissflogii (8 – 12 mm ESD) and large cylindrical R. alata cells (length of 200– 300 mm, width of 12– 16 mm). Strong chlorophyll autofluorescence revealed that the majority of chloroplasts of T. weissflogii cells were hardly or not digested by the gut passage through D. gegenbauri (Fig. 1b and c). Light microscopy showed that the outer transparent region of some pellets consisted mainly of broken and almost fully digested T. weissflogii cells, while cells in the interior more densely packed region of the pellet seemed to be undigested. Digested food particles were embedded in mucous material preventing the membraneless pellet from disaggregation (Köster unpubl. observation). Copepod pellets produced by female E. pileatus were cylindrically shaped, and surrounded by a chitinous peritrophic membrane (Fig. 1d; see insert, white arrow) that often formed a tail at both ends of the pellet. Pellets reached lengths of 400– 600 mm and widths of up to 50– 60 mm, respectively. Compared with doliolid pellets, average copepod pellet volumes were about one order of magnitude lower (0.002 – 0.004 mm3). Pellets consisted mainly of fragmented diatom valves, were densely packed and non-translucent (Fig. 1d). As observed in a previous study, the autofluorescence of digested fragmented food particles was only weak in copepod pellets (see Fig. 1f in Paffenhöfer and Köster, 2005). Ultrastructure of fecal pellets Transmission electron microscopy of pellets revealed the fine structure and state of ingested phytoplankton cells as well as the occurrence and distribution of pellet-associated bacteria. Further, TEM serves to localize the sites of microbial attack. In thin sections from different parts of the gonozooid pellet of D. gegenbauri, we observed numerous intact diatom valves of R. alata and T. weissflogii (Fig. 2a and b). While some of the small T. weissflogii contained cell organelles such as chloroplasts and nuclei in the cytoplasm (Fig. 2a see white arrows, Fig. 3a and b) the larger R. alata cells did not contain cell organelles and only occasionally did cytoplasm remain, indicating that these cells were almost completely 1541 JOURNAL OF PLANKTON RESEARCH j VOLUME 33 j NUMBER 10 j PAGES 1538 – 1549 j 2011 Fig. 2. Transmission electron microscopy micrographs of regions inside a gonozooid fecal pellet of Dolioletta gegenbauri. Thin sections originate from the middle part of the pellet. (a and b) From longitudinally to vertically varying sections of large intact silicate valves of empty (¼digested) Rhizosolenia alata cells (R) and some smaller ones of Thalassiosira weissflogii (T) containing partly digested chloroplasts (white arrow) (c) There are also some regions with fragmented diatom valves (FV) with scarce bacterial colonization (B, bacteria; FV, fragmented valves). digested (Fig. 2b). This observation agrees with the results of earlier work (Paffenhöfer and Köster, 2005) showing that doliolids incompletely digested the ingested food particles, often assimilating ,50% of the ingested organic material. Fragmented diatom frustules were only found intermittently between intact frustules (densely compacted aggregations with a diameter of 30– 100 mm; Fig. 2a and c) because doliolids do not have mouthparts to select and crush food particles, but collect food particles with their mucous net. Most of the flagellates ingested (I. galbana) seemed to be hardly deformed. Epifluorescence micrographs ( photos not shown) showed that the chloroplasts and even the flagellae of I. galbana were preserved. These observations were obtained from T. weissflogii and I. galbana composed pellets that were transferred to sterile (0.2-mm filtered) seawater, formaldehyde (2% v/v)fixed, sonicated (3 7 s, 35 W) to disrupt the pellet structure and stained with acridine orange (final concentration 1:10 000). Transmission electron microscopy analyses confirmed that these flagellates reveal hardly any cellular alterations after being ingested by D. gegenbauri (Fig. 4d–f ). We estimate that the residence time of ingested cells exposed to digestive enzymes does not surpass 30 min at a release rate of 8 pellets h21. That implies a limited period of exposure to digestive fluids during which a flagellate membrane would have to be lysed. Overview cross-sections of calanoid copepod fecal pellets revealed that most of the ingested T. weissflogii cells were fragmented (Fig. 5). The few intact valves did not contain any cell constituents. The peritrophic membrane that is produced in the copepod gut and wraps the ingested food particles (Gauld, 1957; Hansen and Peters, 1997/1998) remained intact in most of the freshly produced pellets up to 16 h after release (Fig. 1d, insert). Transmission electron microscopy micrographs at 800-fold magnification (Fig. 5) clearly showed the integrity of the peritrophic membrane. The membrane was 50– 70-nm thick and consisted of multilayered chitinous microfibrils oriented in different directions (TEM micrographs at 28 000–60 000-fold magnification, not shown). 1542 M. KÖSTER ET AL. j ULTRASTRUCTURE OF DOLIOLID FECAL PELLETS Fig. 3. Transmission electron microscopy micrographs of hardly digested Thalassiosira weissflogii cells inside fecal pellets of Dolioletta gegenbauri. Longitudinal (a) and cross-section (b) of intact cells of Thalassiosira weissflogii. Note the intact cell organelles (N, nucleus; No, nucleolus; Ch, chloroplasts). There are hardly any bacteria (B) in the surroundings of these cells. (c and d) longitudinal sections of partly digested cells that contain degenerated cell organelles and cytoplasm (Cp). These cells reveal a dense bacterial colonization outside their silicate valves. Bacterial colonization of fecal pellets In loosely structured doliolid pellets, bacteria occurred both close to the external surfaces and within pellets. Thin sections of a gonozooid pellet of D. gegenbauri revealed that sizes and distribution of bacteria were variable. Most of the bacteria were rods with lengths ranging from 0.4 to 1.5 mm; dividing cells made up ,0.5% of total bacteria. The majority of bacteria occurred in clusters in the vicinity of partly digested T. weissflogii cells (up to 54 bacteria 1000 mm22 thin section area Fig. 3c and d), and ingested flagellates (up to 82 bacteria 1000 mm22 thin section area; Fig. 4a and b), and in regions with unidentifiable organic matter (up to 160 bacteria 1000 mm22). We found numerous examples for these observations. Extremely scarce or no bacterial colonization was found in the vicinity of intact T. weissflogii cells (with nuclei and chloroplasts; Fig. 3a and b) and also not inside and outside of empty valves of T. weissflogii and R. alata (Fig. 2b). A few scattered bacteria were found among fragmented valves (,25 bacteria 1000 mm22; Fig. 2c). A Kruskal – Wallis Test (non-parametric analysis of variance by ranks; Conover, 1980) was applied to bacterial abundances near partly digested as well as completely digested (empty) cells of T. weissflogii, and cells of R. alata. The null hypothesis that bacterial abundances were the same in each of the three cases was rejected. The ensuing multiple comparison test revealed that bacterial abundances near empty cells of T. weissflogii and R. alata did not differ significantly while bacteria near partly empty cells of T. weissflogii were more abundant than near the empty cells of the two phytoplankton species. Bacterial abundances near cells of I. galbana far surpassed those near the partly digested T. weissflogii. Freshly egested copepod pellets revealed hardly any bacteria on their exterior surfaces ( peritrophic membrane). The majority of bacteria were found in the interior lumen of the pellets. 1543 JOURNAL OF PLANKTON RESEARCH j VOLUME 33 j NUMBER 10 j PAGES 1538 – 1549 j 2011 Fig. 4. Transmission electron microscopy micrographs of hardly digested phytoflagellates [Isochrysis galbana (Ig); Rhizosolenia alata (Rho)] inside Dolioletta gegenbauri fecal pellets. (a–c) Note the numerous bacteria (B, bacteria; BC, bacterial cluster) in the vicinity of the flagellates (d) Isochrysis galbana. The flagellates’ two chloroplasts (Ch) with thylachoid membranes, the pyrenoid (P), the nucleus (N) and mitochondria (M) are visible. (e and f ) Longitudinal sections of Rhodomonas sp. Note the bacteria in the surroundings and the Isochrysis galbana cell that is attached to the outer membrane of Rhodomonas sp. 1544 M. KÖSTER ET AL. j ULTRASTRUCTURE OF DOLIOLID FECAL PELLETS copepods, their pellet morphology differs in many respects. Ultrastructure of fecal pellets Fig. 5. Transmission electron microscopy micrographs of fecal pellets of female Eucalanus pileatus. (a and b) Fecal pellets contained mainly fragmented valves of Thalassiosira weissflogii (Fd). There were only a few intact diatom valves (Id). The white arrows indicate the outer peritrophic membrane (Pm) of the pellet. DISCUSSION The morphology of fecal pellets influences their bacterial colonization and degradation (as well as their sedimentation behavior (residence time) and thus their contribution to the pelagic food web and vertical carbon flux (e.g. Hofmann et al., 1981; Wexels Riser et al., 2007; Giesecke et al., 2010; Patonai et al. 2011). Our TEM observations revealed differences in the ultrastructure of a doliolid and a copepod fecal pellet as well as the condition of ingested phytoplankton cells, and the occurrence of bacteria on/within the pellets. Owing to different feeding mechanisms of doliolids and A range of papers have covered the ultrastructure of zooplankton fecal pellets by scanning electron microscopy (SEM, e.g. Turner, 1984a, b; Beaumont et al., 2002; Jansen and Bathmann, 2007). This technique enables the surfaces of both fragmented/broken and intact pellets to be observed (e.g. Jansen et al., 2006). Turner (Turner, 1984b) stripped off the peritrophic membrane to make the interior of the pellets visible. Alternatively, TEM can be applied to investigate the ultrastructure of fecal pellets. Thin sections allow the presence and distribution of both internal and membrane-associated bacteria in different regions of a pellet to be investigated. In addition, the detailed ultrastructure of digested food particles including their cell organelles and membranes is visible. We used the detailed information on their state to evaluate the degree of digestion. Especially, as concerns the intactness of the peritrophic membrane as an indicator for pellet integrity, the application of TEM is helpful. Although TEM has the potential to provide valuable information on the ultrastructure of pellets, it has been rarely used previously. To the authors’ knowledge, the only TEM investigations are analyses of the composition of pellets of salps and pteropods (Silver and Bruland, 1981) and unidentified copepod pellets (Gowing and Silver, 1983). The latter study presents TEM micrographs of the structure of cylindrical and ellipsoid pellets (most likely originating from copepods) obtained from traps, and from shipboard-feeding crab larvae. They showed damaged as well as intact phytoplankton cells and bacteria. Our study describes the ultrastructure of doliolid and copepod pellets. Our observations revealed seemingly intact, partly empty and entirely empty phytoplankton cells in doliolid fecal pellets. While the R. alata valves were empty, most of the T. weissflogii cells seemed to be not or hardly affected (well preserved chloroplasts and other cell organelles). Some T. weissflogii cells contained cytoplasm remains, and a few were empty. So, why does the integrity of diatom cells differs between taxa, and why does the percentage of hardly digested T. weissflogii cells far exceed that of R. alata cells? We assume that digestive enzymes of D. gegenbauri gonozooids might have had an easier access to R. alata cells than to T. weissflogii cells. Differences in the fine structure of silicate valves (e.g. number and size of pore sizes, stability of valves; shielding surface polymers around algae) probably result in a higher percentage of the smaller 1545 JOURNAL OF PLANKTON RESEARCH j VOLUME T. weissflogii cells surving gut passage and they may have been more resistant to the digestive enzymes of D. gegenbauri than the larger R. alata cells. Whether ingested viable bacteria may supplement the hydrolysis of food components, which the doliolid cannot hydrolyze because it lacks the necessary enzymes, remains unclear. Fecal pellets of E. pileatus contained numerous broken valves among the occasional undamaged valves of T. weissflogii (Fig. 5). This explains the high assimilation efficiency of calanoid copepods ingesting diatoms when compared with doliolids ingesting the same cells (e.g. Paffenhöfer and Köster, 2005). Bacterial colonization of fecal pellets The initial studies on thaliacean fecal pellets and associated bacteria included observations on pellets of D. gegenbauri (Pomeroy and Deibel, 1980; Pomeroy et al., 1984). The latter stated ‘They (the pellets) contain large amounts of undigested or partly-digested materials and consequently are excellent substrates for bacteria’. Pellets of D. gegenbauri, which had been feeding on enriched natural seston for 6 h, had by that time few bacteria, but numerous large bacteria after 21 h. Similarly, D. gegenbauri pellets, from feeding on a natural suspension of the small diatom Thalassiosira subtilis for 6 h, had no bacteria; however, after 21 h they had large motile rods which were also found in the surrounding water (Pomeroy and Deibel, 1980). Overall, microscopic examination of such feces ‘showed a rapid increase in bacteria within the feces during the first day’. This the authors attributed to excess DOM, associated with the partly digested food particles within fecal particles, which is leached as indicated by primary amines (Pomeroy et al., 1984). Kiørboe et al. (Kiørboe et al., 2002) reported bacterial colonization rates of artificial organic-rich (leaky) particles even in the order of minutes. These authors described bacterial colonization of aggregates ( pellets) as a highly dynamic process depending on the motility of bacteria, their attachment/ detachment behavior, as well as the size and hydrodynamic environment of the aggregates. As doliolids create a cilary current to draw food particles into their mouth (Bone et al., 1997), a certain percentage of free-living bacteria should also be retained in their mucous filter via filter feeding. Hitherto no data on the mesh size of the filter of doliolids are available (due to unsuccessful fixation of the filter net for electron microscopy, Bone et al., 2003). In our study, many freeliving bacteria smaller than 1 mm were found within D. gegenbauri pellets (Figs. 3 and 4) suggesting that the filter net might be capable of trapping submicron particles. The bacteria trapped in the filter net provide, in 33 j NUMBER 10 j PAGES 1538 – 1549 j 2011 addition to the attached bacteria of ingested food particles, an additional food source for the doliolids. Whether bacteria survive the doliolids’ gut passage and whether their digestive enzymes play a role in the decomposition of ingested phytoplankton is still a subject of investigation. At the same time bacteria living in the water surrounding the recently ejected pellets should be attracted by various dissolved organic compounds that are released by the partly digested phytoplankton cells (Pomeroy, 1992). We want to point out that while Pomeroy and Deibel (Pomeroy and Deibel, 1980) offered natural particulate matter to doliolids, we offered phytoplankton cells grown in non-axenic conditions in the laboratory. Such phytoplankton cultures contain organic matter degrading bacteria of unknown origin, physiology and ecology. Transmission electron microscopy observations of doliolid pellets revealed that bacterial colonization varied in the vicinity of ingested phytoplankton species of different state (Figs. 3 and 4). The degree of bacterial colonization is probably caused by the availability of assimilable DOC. When assimilable organic material leaks through the valves of partly digested diatoms and/or flagellates, nutrient-rich microhabitats are created around the algae/flagellates serving as nutrient sources for heterotrophic bacteria. This is reflected in increased bacterial numbers in these highly nutritive areas (see Figs. 3 and 4). The majority of bacteria in the vicinity of partly digested food particles were ,1 mm, but there always appeared some large rods with lengths of 1.5 – 2.0 mm and widths of 0.5 mm. As the age of our pellets was mostly in the range of several hours (between 2 and 16 h), bacteria would have sufficient time to grow within the pellets, and/or position themselves in the vicinity of digested food items containing organic matter. Dividing cells were not observed among the bacteria inside pellets indicating that the residence time of fecal pellets in the gut was shorter than the generation time of food-associated bacteria. Contrary to bacteria-enriched areas, nearly completely digested microalgae such as empty valves of T. weissflogii and R. alata do not provide any valuable/ assimilable substrates for bacteria. Therefore, these areas were far less colonized by bacteria. The fact that no bacterial colonization was observed inside numerous diatom valves and among fragmented valves confirms our assumption that the digestion of algal cells was caused by hydrolytic digestive enzymes of the doliolids’ gut and not by bacteria. In 50 thin slices of a freshly egested copepod pellet (,16 h) we found hardly any bacteria (1 bacteria 1000 mm22 thin section area) on the peritrophic 1546 M. KÖSTER ET AL. j ULTRASTRUCTURE OF DOLIOLID FECAL PELLETS membrane but quite a few (,25 bacteria 1000 mm22 thin section area) within the pellet (Fig. 5). Several authors (Lautenschlager et al., 1978; Gowing and Silver, 1983) also reported that the surfaces of freshly ejected (age ,7 h) crustacean pellets ( planktonic copepods and gammarids, respectively) are devoid of bacteria. With ongoing degradation, copepod fecal pellets were densely colonized by bacteria (‘bacterial lawn’) as also has been observed by other investigators (cf. Turner, 1979; Gowing and Silver, 1983; Turner, 2002). Whether the majority of copepod pellet associated internal bacteria originate from the enteric flora of the copepod and/or bacteria attached to ingested food particles remains uncertain (compare e.g. Nagasawa and Nemoto, 1988; de Troch et al., 2010). Lawrence et al. (Lawrence et al., 1993) reported that calanoid copepods produced bacteria-free pellets when axenic food was offered. They concluded that pellet-associated bacteria did not originate from the copepods’ gut but were derived from ingested food particles. Furthermore, the diversity of the bacterial community of fecal pellets of harpacticoid copepods is assumed to be broader than that of their food source (diatoms). This increased bacterial richness depends largely from the ingested food, the copepod species and the age of fecal pellets (de Troch et al., 2010). Pellet morphology, sinking velocity and potential degradation The first fundamental microbiological studies of the degradation of doliolid fecal pellets go back to Pomeroy in the early 1980s (Pomeroy and Deibel, 1980; Pomeroy et al., 1984). Later and recent investigations that have applied more sensitive methods to determine microbial pellet degradation (e.g. Thor et al., 2003, enzymatic degradation activity; Olsen et al., 2005; microcalorimetry; Shek and Liu, 2010, oxygen consumption measurements by microsensors) were exclusively devoted to copepod pellets. None of these process-oriented studies were accompanied by microscopic methods. Thus, there is a need to combine process-oriented investigations with direct microscopic methods to assess the degradation state of doliolid and copepod pellets and their contents. As pellet characteristics (shape, packaging, density etc.) differ strongly, we expect different degradation pathways for the pellets of the doliolid D. gegenbauri and the calanoid copepod E. pileatus. The loose packaging of doliolid pellets (low density), their flat shape and the intermittent occurrence of threadlike structures are responsible for their low sinking velocities and long retention times in the water column. Patonai et al. (Patonai et al., 2011) determined average sinking velocities of 20– 60 m d21 for pellets of 6 – 7 mm long gonozooids. Since doliolid pellets will therefore remain in the water column for a relatively long period, and seawater heterotrophic bacteria have immediate access to the membraneless pellets, those will most likely be rapidly mineralized in the water column. In comparison, pellets of females of E. pileatus are more densely packed than doliolid pellets, and have much smaller volumes than the latter (Fig. 1). While their sinking rates are in the same range as those of doliolid pellets (Patonai et al., 2011), observations by Ploug et al. (Ploug et al., 2008) reveal an average sinking rate of 322 m d21of pellets from the calanoid copepod Temora longicornis (females) feeding at 158C on T. weissflogii at 430 mg C L21. Those pellets’ volumes at 106 mm3 were slightly lower than those of E. pileatus pellets also feeding on T. weissflogii in our experiment. It appears from their detailed study as well as Feinberg and Dam (Feinberg and Dam, 1998) that ‘pellet size and density depend on copepod species, food concentration and food source.’ The early decomposition of copepod pellets is initiated via mechanical breakdown and/or microbial attack of the chitinous peritrophic membrane. As soon as the pellets are egested their peritrophic membranes provide a colonizable substratum (surface) for bacteria occurring in the surrounding water. While the peritrophic membrane of up to 16 h old E. pileatus pellets was intact (Fig. 5) and colonized by only a very few bacteria, the peritrophic membrane of pellets of Acartia tonsa was already disrupted within 1.5 h (Hansen et al., 1996). With ongoing degradation, we observed microscopically that pellets became more porous and lighter, indicating to the activity of organic matter degrading bacteria with different physiological potential. The degree and rate of membrane and pellet degradation depends on coprophagy, bacterial colonization and pellet composition (Hansen et al., 1996). Up to the present, it is still a methodological challenge to distinguish between internal-origin and external-origin pellet-associated bacteria responsible for membrane and pellet degradation. A first indication of the origin of bacteria is the TEM micrographs showing bacteria inside pellets with an integral peritrophic membrane (Fig. 5). One of the next steps to consider would be to conduct similar studies on doliolid pellets produced under in situ conditions. This would be accompanied by quantifying abundances of phytoplankton and heterotrophic organisms (including bacteria and protists, zooplankton) in the pellets and the surrounding water. In addition, pellets of smaller (body length of 2 – 3 mm) zooids (e.g. small gonozooids released by phorozooids), which are dominant in blooms, ought to be included in future considerations. New staining techniques (e.g. Cell 1547 JOURNAL OF PLANKTON RESEARCH j VOLUME Tracker Green) will enable our approach of the ultrastructure of fecal pellets to be combined with functional analyses of the viability of pellet associated cells (undigested, digested phytoplankton species, bacteria). 33 j NUMBER 10 j PAGES 1538 – 1549 j 2011 Gibson, D. M. and Paffenhöfer, G.-A. (2000) Feeding and growth rates of the doliolid, Dolioletta gegenbauri Uljanin (Tunicata, Thaliacea). J. Plankton Res., 22, 1485– 1500. Giesecke, R., González, H. E. and Bathmann, U. (2010) The role of the chaetognath Sagitta gazellae in the vertical carbon flux of the Southern Ocean. Polar Biol., 33, 293– 304. Gowing, M. M. and Silver, M. W. (1983) Origins and microenvironments of bacteria mediating fecal pellet decomposition in the sea. Mar. Biol., 73, 7 –16. AC K N OW L E D G E M E N T S We thank Captain Raymond Sweatte and the crew of R/V Savannah for their professional cooperation and their support collecting doliolids and copepods on the continental SE shelf of the USA. We greatly acknowledge Dr Gesine Roth (Deutscher Akademischer Austausch Dienst, German Academic Exchange Service) for the support of Dr M. Köster in building up an American-German cooperation between the Skidaway Institute of Oceanography, and the University of Greifswald. We are grateful for the constructive comments of three anonymous reviewers. Hansen, B., Fotel, F. L., Jensen, N. J. et al. (1996) Bacteria associated with a marine planktonic copepod in culture. II. Degradation of fecal pellets produced on a diatom, a nanoflagellate or a dinoflagellate diet. J. Plankton Res., 18, 275–288. Hansen, U. and Peters, W. (1997/1998) Structure and permeability of the peritrophic membranes of some small crustaceans. Zool. Anz., 236, 103– 108. Hofmann, E. E., Klinck, J. M. and Paffenhöfer, G.-A. (1981) Concentrations and vertical fluxes of zooplankton fecal pellets on a continental shelf. Mar. Biol., 61, 327–335. Jansen, S. and Bathmann, U. (2007) Algae viability within copepod faecal pellets, evidence from microscopic examinations. Mar. Ecol. Prog. Ser., 337, 145–153. Jansen, S., Riser, C. W., Wassmann, P. et al. (2006) Copepod feeding behaviour and egg production during a dinoflagellate bloom in the North Sea. Harmful Algae, 5, 102– 112. FUNDING The research for this paper was supported by two grants from the U.S. National Science Foundation (OCE 0825999, OCE 1031263). Kiørboe, T., Grossart, H. -P., Ploug, H. et al. (2002) Mechanisms and rates of bacterial colonization of sinking aggregates. Appl. Environ. Microbiol., 68, 3996– 4006. REFERENCES Lawrence, S. G., Ahmed, A. and Azam, F. (1993) Fate of particlebound bacteria ingested by Calanus pacificus. Mar. Ecol. Prog. Ser., 97, 299 –307. Beaumont, K. L., Nash, G. V. and Davidson, A. T. (2002) Ultrastructure, morphology and flux of microzooplankton faecal pellets in an East Antarctic fjord. Mar. Ecol. Prog. Ser., 245, 133–148. Bone, Q., Braconnot, J. -C. and Carré, C. (1997) On the filter feeding of Doliolum (Tunicata, Thaliacea). J. Exp. Mar. Biol. Ecol., 214, 179–193. Bone, Q., Carré, C. and Chang, P. (2003) Tunicate feeding filters. J. Mar. Biol. Assoc. UK, 83, 907 –919. Lautenschlager, K. P., Kaushik, N. K. and Robinson, J. B. (1978) The peritrophic membrane and faecal pellets of Gammarus lacustris limnaeus Smith. Freshw. Biol., 8, 207– 211. Nagasawa, S. and Nemoto, T. (1988) Presence of bacteria in guts of marine crustaceans and on their fecal pellets. J. Plankton Res., 10, 559 –564. Olsen, S. N., Westh, P. and Hansen, B. W. (2005) Real-time quantification of microbial degradation of copepod fecal pellets monitored by isothermal microcalorimetry. Aquat. Microb. Ecol., 40, 259–267. Conover, W. J. (1980) Practical Nonparametric Statistics. John Wiley & Sons, New York. Paffenhöfer, G.-A., Atkinson, L. P., Lee, T. N. et al. (1995) Distribution and abundance of thaliaceans and copepods off the southeastern USA during winter. Cont. Shelf Res., 15, 255–280. Deibel, D. (1998) The abundance, distribution and ecological impact of doliolids. In Bone, Q. (ed.), The Biology of Pelagic Tunicates. Oxford University Press, Oxford, England, pp. 171– 186. Paffenhöfer, G.-A. and Gibson, D. M. (1999) Determination of generation time and asexual fecundity of doliolids (Tunicata, Thaliacea). J. Plankton Res., 21, 1183– 1189. Deibel, D. and Paffenhöfer, G.-A. (2009) Predictability of patches of neritic salps and doliolids (Tunicata, Thaliacea). J. Plankton Res., 31, 1571– 1579. Paffenhöfer, G.-A. and Knowles, S. C. (1979) Ecological implications of fecal pellet size, production and consumption by copepods. J. Plankton Res., 37, 35– 49. de Troch, M., Cnudde, C., Willems, A. et al. (2010) Bacterial colonization on fecal pellets of harpacticoid copepods and on their diatom food. Microbiol. Aquat. Syst., 60, 581– 591. Paffenhöfer, G.-A. and Köster, M. (2005) Digestion of diatoms by planktonic copepods and doliolids. Mar. Ecol. Prog. Ser., 297, 303–310. Feinberg, L. R. and Dam, H. G. (1998) Effects of diet on dimensions, density and sinking rates of fecal pellets of the copepod Acartia tonsa. Mar. Ecol. Prog. Ser., 175, 87–96. Paffenhöfer, G.-A. and Köster, M. (2011) From one to many, On the life cycle of Dolioletta gegenbauri Uljanin (Tunicata, Thaliacea). J. Plankton Res., 33, 1139– 1145. Gauld, D. T. (1957) A peritrophic membrane in calanoid copepods. Nature, 179, 325–326. Paffenhöfer, G.-A. and Lee, T. N. (1987) Development and persistence of patches of Thaliacea. S. Afr. J. Mar. Sci., 5, 305–318. 1548 M. KÖSTER ET AL. j ULTRASTRUCTURE OF DOLIOLID FECAL PELLETS Patonai, K., El-Shaffey, H. and Paffenhöfer, G.-A. (2011) Sinking velocities of copepod and doliolid fecal pellets. J. Plankton Res., 33, 1146– 1150. Ploug, H., Iversen, M. H., Koski, M. et al. (2008) Production, oxygen respiration rates, and sinking velocity of copepod fecal pellets: direct measurements of ballasting by opal and calcite. Limnol. Oceanogr., 53, 469–476. Pomeroy, L. R. (1992) The microbial food web. Oceanus, 35, 28– 35. Pomeroy, L. R. and Deibel, D. (1980) Aggregation of organic matter by pelagic tunicates. Limnol. Oceanogr., 25, 643– 652. Pomeroy, L. R., Hanson, R. B., Mc Gillivary, P. A. et al. (1984) Microbiology and chemistry of fecal products of pelagic tunicates, rates and fates. Bull. Mar. Sci., 35, 426– 439. Shek, L. and Liu, H. (2010) Oxygen consumption rates of fecal pellets produced by three coastal copepod species fed with a diatom Thalassiosira pseudonana. Mar. Poll. Bull., 60, 1005–1009. Silver, M. W. and Bruland, K. W. (1981) Differential feeding and fecal pellet composition of salps and pteropods, and the possible origin of the deep-sea flora and olive-green ‘cells’. Mar. Biol., 67, 193–199. Thor, P., Dam, H.-G. and Rogers, D. R. (2003) Fate of organic carbon released from decomposing copepod fecal pellets in relation to bacterial production and ectoenzymatic activity. Aquat. Microb. Ecol., 33, 279 –288. Turner, J. T. (1979) Microbial attachment to copepod fecal pellets and its possible significance. Trans. Am. Microsc. Soc., 98, 131– 135. Turner, J. T. (1984a) Zooplankton feeding ecology, contents of fecal pellets of the copepods Temora turbinata and T. stylifera from continental shelf and slope waters near the mouth of the Mississippi River. Mar. Biol., 82, 73– 83. Turner, J. T. (1984b) Zooplankton feeding ecology, contents of fecal pellets of the copepods Eucalanus pileatus and Paracalanus quasimodo from continental shelf waters of the Gulf of Mexico. Mar. Ecol. Prog. Ser., 15, 27– 46. Turner, J. T. (2002) Zooplankton fecal pellets, marine snow and sinking phytoplankton blooms. Aquat. Microb. Ecol., 27, 57–102. Wexels Riser, C., Reigstad, M., Wassmann, P. et al. (2007) Export or retention? Copepod abundance, faecal pellet production and vertical flux in the marginal ice zone through snap shots from the northern Barents Sea. Polar Biol., 30, 719–730. 1549