Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
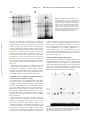
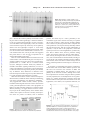
Activation of Heat-Shock Factor by Stretch-Activated Channels in Rat Hearts Jiang Chang, MD, PhD; Jeremy S. Wasser, PhD; Richard N.M. Cornelussen, PhD; A.A. Knowlton, MD Downloaded from http://circ.ahajournals.org/ by guest on June 12, 2017 Background—Previously, we have observed that the isolated, erythrocyte-perfused rabbit heart has increased levels of heat-shock protein (HSP) 72 after a mild mechanical stress. We hypothesized that stretch-activated ion channels (SACs) mediated this increase. Methods and Results—To test this hypothesis, we subjected isolated, perfused rat hearts to mechanical stretch. Gel mobility shift assay showed that heat-shock factor (HSF) was activated in hearts with mechanical stretch, but not in controls. Supershift experiments demonstrated that HSF1 was the transcription factor. Northern blots revealed the concomitant increase in HSP72 mRNA in stretched rat hearts. In a separate set of experiments, gadolinium, an inhibitor of SACs, was added to the perfusate. Gadolinium inhibited the activation of HSF and decreased HSP72 mRNA level. Because gadolinium can inhibit both SACs and L-type calcium channels, we perfused a group of hearts with diltiazem, a specific L-type calcium channel blocker, to eliminate the involvement of L-type calcium channels. Diltiazem failed to inhibit the activation of HSF. Conclusions—Stretch in the rat heart results in activation of HSF1 and an increase in HSP72 mRNA through SACs. This represents a novel mechanism of HSF activation and may be an important cardiac signaling pathway for hemodynamic stress. (Circulation. 2001;104:209-214.) Key Words: stretch 䡲 ion channels 䡲 calcium 䡲 proteins C ells from all organisms respond to stress by synthesizing heat-shock, or stress, proteins (HSPs). HSPs are a group of highly conserved proteins classified by their molecular weights. Many studies have demonstrated that HSPs play an important role in the protection of stressed organisms.1– 4 The regulation of heat-shock response is mediated by ⱖ1 cytosolic proteins known as heat-shock factors (HSFs) that interact with a specific regulatory element, the heat-shock element, in the promoter regions of the HSP gene.5 HSF activation is one of the earliest responses observed in many different cells exposed to stresses, such as ischemia, hypoxia, hyperthermia, and mechanical stretch.6 –9 Mechanical stretch and pressure overload have been shown to induce HSP72 expression in a variety of different cells and tissues.8,10 –12 Several studies reported that stretch resulted in HSF1 activation and HSP72 expression in rat aorta,8,13,14 but few data are available for the heart.9,15 Previously, we observed that even a single myocardial stretch can cause increased expression of HSP72 in the isolated erythrocyteperfused rabbit heart.15 The mechanism is unknown, but several research groups have postulated that stretch-activated ion channels (SACs) may act as mechanotransducers to mediate stretch-induced gene expression.16,17 As yet, however, there is no direct evidence that stretch-induced HSP72 expression is mediated by SACs. Therefore, the purpose of this study was 3-fold. First, we used 2 different standard isolated heart perfusion models (Langendorff and working) to demonstrate that stretch alone was sufficient to induce the heat-shock response in rat hearts. These models eliminated any influence of circulating neural and/or humoral factors. Second, we tested the hypothesis that the induction of HSP72 mRNA was via the activation of HSF, specifically HSF1. Third, we used gadolinium, an SAC blocker, to test the hypothesis that the stretch-induced heatshock response was mediated by SACs. Methods Langendorff Perfusion Male Sprague-Dawley rats (228⫾9 g, n⫽43; Harlan Inc, Indianapolis, Ind) were heparinized and anesthetized with sodium pentobarbital. The heart was removed and placed in ice-cold Ringer’s solution (mmol/L: NaCl 120.0, KCl 4.7, NaHCO3 18.0, glucose 11.0, MgSO4 1.5, and CaCl2 2.5, equilibrated with 95% O2/5% CO2). Retrograde perfusion (Langendorff) was quickly established with a perfusion pressure of 95 to 100 cm H2O. The pH and temperature of Received January 18, 2001; revision received March 14, 2001; accepted March 21, 2001. From Texas A&M University, Department of Veterinary Physiology and Pharmacology, College Station (J.C., J.S.W.), and Baylor College of Medicine and VA Medical Center, Houston (R.N.M.C., A.A.K.), Tex. Dr Chang is now at the Department of Cell Biology, Baylor College of Medicine, Houston, Tex. Dr Cornelussen is now at the Cardiovascular Research Institute Maastricht, Department of Physiology, Maastricht University, Maastricht, the Netherlands. Guest Editor for this article was Derek M. Yellon, PhD, DSc, Hon MRCP, University College London Hospitals and Medical School, London, UK. Correspondence to Dr A.A. Knowlton, Cardiology Research, Baylor College of Medicine, VA Medical Center 151C, 2002 Holcombe, Houston, TX 77030. E-mail [email protected] © 2001 American Heart Association, Inc. Circulation is available at http://www.circulationaha.org 209 210 Circulation July 10, 2001 the perfusate were 7.4 and 37°C, respectively. A PE90 tubing was inserted into the left ventricle (LV) and through the apex for thebesian drainage. We placed a fluid-filled latex balloon into the LV via the left atrium and connected it to a transducer to monitor pressures. Initial LV end-diastolic pressure (LVEDP) was adjusted to 10 mm Hg by adjustment of balloon volume. The heart was submerged in a water-jacketed constant-temperature chamber that contained Ringer’s solution equilibrated with 95% O2/5% CO2. The protocol was approved by the Animal Welfare Committee of Texas A&M University. Working Heart Perfusion The heart was perfused by the Langendorff method first without balloon or ventricular drain. The left atrium was connected to a second cannula and perfused at a preload pressure of 10 to 15 cm H2O. Afterload was set at 50 to 70 cm H2O by adjustment of the height of the aortic outflow line above the heart. A compliance chamber was added to the aortic outflow line. The heart was perfused in the working mode (ejecting LV). Downloaded from http://circ.ahajournals.org/ by guest on June 12, 2017 Maximum Ventricular Developed Pressure To monitor cardiac contraction, we collected cardiac maximum ventricular developed pressure (Pmax) throughout the experiments with a pressure transducer connected to a computerized dataacquisition system (Labview). Gel Mobility Shift Assay As described previously, a single-strand self-complementary oligonucleotide containing the 5⬘-nGAAn-3⬘ repeats was synthesized (5⬘-CTAGAAGCTTCTAGAAGCTTCTAG-3⬘), annealed, and endlabeled with [␥-32P]ATP.18,19 Because HSF is normally present in the cell in an inactive form, we were able to use whole-cell lysates for our studies. Samples were processed as previously described. Cold competition and supershift studies were done as previously described. Figure 2. A, GMSA in Langendorff-perfused hearts (B lanes) and nonperfused rat hearts (A lanes). B, Effects of varying stretch (LVEDP) on HSF activation in Langendorff-perfused hearts: 10 mm Hg LVEDP (lane B), 7 to 8 mm Hg LVEDP (lane C), and 5 mm Hg LVEDP (lane D). Heart in lane B showed strongest HSF activity shift band, followed by lanes C and D, indicating that different amount of stretch can influence HSF activity. Lane A, nonperfused heart. NS indicates nonspecific binding; HSF, activation with band shift. RNA Isolation and Northern Blot Analysis After each experiment, heart ventricles were rapidly dissected and freeze-clamped in liquid nitrogen. Total RNA isolation was performed with a STAT-60 kit (Tel-test, Inc). Northern blotting was performed as described previously.6,15 Experimental Protocols All hearts were perfused for a 20-minute stabilization period followed by 1 of 7, 60-minute experimental protocols (Figure 1). As a positive control, 2 working hearts were perfused at 43°C for 13 minutes. Statistical Analysis Data were analyzed by 1-way ANOVA, followed by the StudentNewman-Keuls test (SigmaStat software, SPSS Inc). A value of P⬍0.05 was considered significant. Data are presented as mean⫾SEM. Results HSF Activation Was Observed in Langendorff Stretched Hearts but Not in Working Hearts Figure 1. Experimental protocols for Langendorff and working perfusions. All hearts were perfused for 20-minute stabilization period. Hearts in Langendorff mode were perfused with gadolinium (SAC and L-type calcium channel blocker) or diltiazem (specific L-type calcium channel blocker) for 60 minutes. To impose mechanical stretch in working perfusion mode, aortic cannula with small diameter (⬍1 mm ID) was used to mimic aortic stenosis. Two hearts were perfused at 43°C for 13 minutes as positive controls. n⫽hearts/group. Previously, we observed that a mechanical stretch induced the expression of HSP7215 in the rabbit heart. In the present study, we again demonstrated that mechanical stretch, which included stretch from insertion of the apical drain and placement of the ventricular balloon, initiated the heat-shock response in the Langendorff perfusion. The gel mobility shift assays (GMSAs) showed that this stretch activated HSF (Figure 2A). In pilot experiments, the effects of different amounts of stretch on HSF activity were examined. This was done by varying the initial LVEDP in Langendorff perfusions. As shown in Figure 2B, an LVEDP of 10 mm Hg correlated with the strongest HSF activation (lane B in Figure 2B), followed by an LVEDP of 7 to 8 mm Hg (lane C in Figure 2B). Only minimal HSF activation occurred in the Chang et al Heat-Shock Factor and Stretch-Activated Channels 211 Figure 3. A, GMSA in Langendorff (A lanes), working perfusion with aortic stenosis (B lanes), and working perfusion (C lanes). B, Cold competition and supershift assay for stretched rat hearts. A lanes, Langendorff perfusion; B lanes, addition of excess cold heat-shock element oligo; C lanes, antibody (1:5, vol/vol) to HSF1 showing positive supershift (SS); D lanes, antibody (1:5, vol/vol) to HSF2 showing no supershift bands. Abbreviations as in Figure 2. Downloaded from http://circ.ahajournals.org/ by guest on June 12, 2017 heart with an LVEDP of 5 mm Hg (lane D in Figure 2B). Thus, the increase in HSF activation induced by stretch is correlated with the amount of stretch imposed on the heart. In contrast, working hearts, which are closer to a physiological perfusion, showed little activation of HSF (C lanes in Figure 3A) compared with the Langendorff- perfused hearts (A lanes in Figure 3A). To further test that mechanical stretch can induce the activation of HSF, we perfused a separate set of hearts in the working mode using an aortic cannula with a small diameter (⬍1 mm in ID) to mimic aortic stenosis. LV afterload was thereby increased, and the ventricular wall stress was enhanced. This resulted in a degree of activation of HSF similar to that observed in Langendorff hearts (B lanes, Figure 3A). Specific activation of HSF was confirmed by the cold heat-shock element competition assay (B lanes, Figure 3B). The addition of antibodies to HSF1 and HSF2 showed a supershift only with anti-HSF1 (C lanes, Figure 3B), indicating that it is HSF1 that is activated by stretch. HSF2 was not activated (no supershift bands in D lanes, Figure 3B). mol/L), if anything, increased activation of HSF (lane G, Figure 6). Figure 6 summarizes HSF binding activities in hearts from all of the experimental protocols: no perfusion (A), working perfusion (B), Langendorff perfusion (C), 10 mol/L gadolinium (D), 50 mol/L gadolinium (E), 1 mol/L diltiazem (F), and 3 mol/L diltiazem (G). As a positive control, hearts with heat shock (43°C for 13 minutes) were analyzed and showed similar degrees of activation of HSF (H lanes, Figure 6). Analysis in HSP72 mRNA Expression After 60 minutes, we found that the HSP72 mRNA level in Langendorff-perfused rat hearts increased compared with nonperfused control rat hearts (Figure 7). Northern blotting showed that gadolinium attenuated this increase at both doses (Figure 7). No dose-dependent inhibition was observed. This observation is consistent with the GMSA, which showed that HSF activity was decreased by 50% by gadolinium. Gadolinium but Not Diltiazem Inhibits Mechanical Stretch–Induced Activation of HSF We perfused hearts in the Langendorff mode with gadolinium, an SAC blocker. As shown in Figure 4 (C and D lanes) and Figure 5 (B and C lanes), gadolinium 10 and 50 mol/L markedly decreased HSF activation. Densitometric measurement of the GMSA showed a 45% and a 50% decrease in HSF binding activity, respectively, compared with Langendorff perfusion without gadolinium (Figure 5A, P⬍0.05). Although the high dose of gadolinium tended to suppress HSF activation more than the low dose, this was not statistically significant. Because gadolinium can block both SACs and L-type calcium channels, it was important to identify whether HSF activation was mediated by L-type calcium channels. To test this, hearts were perfused with diltiazem, a specific L-type calcium channel blocker. Two different concentrations (1 and 3 mol/L) of diltiazem were used, and both failed to inhibit the activation of HSF (D lanes in Figure 5B, and F and G lanes in Figure 6), indicating that gadolinium attenuates HSF activity via SACs. The high concentration of diltiazem (3 Figure 4. Gadolinium effect on Langendorff-perfused (stretched) rat hearts. A, Nonperfusion; B, Langendorff perfusion; C, Langendorff perfusion with gadolinium (10 mol/L); and D, gadolinium (50 mol/L). Abbreviations as in previous figures. 212 Circulation July 10, 2001 Downloaded from http://circ.ahajournals.org/ by guest on June 12, 2017 Figure 5. A, Comparison of HSF activation in (bar A) Langendorff (n⫽4) and (bar B) Langendorff with 10 mol/L gadolinium (n⫽2), (bar C) with 50 mol/L gadolinium (n⫽3), and (bar D) with 1 mol/L diltiazem (n⫽3) rat hearts. *P⬍0.05 vs control Langendorff. B, Representative gel shift for gadolinium and diltiazem effects on Langendorff (stretched) rat hearts. Abbreviations as in previous figures. Maximum Ventricular Developed Pressure Pmax from all groups was stable during perfusions (Figure 8). There was no significant difference in Pmax between the start and completion of perfusion for any of the groups. Furthermore, there was no significant difference in Pmax among the gadolinium, diltiazem, and working-mode groups compared with Langendorff-perfused control hearts. Discussion In the present study, we demonstrated that stretch initiated the heat-shock response in rat hearts by activating HSF1, and this was followed by an increase in HSP72 mRNA. The intensity of HSF activation correlated with the amount of stretch, suggesting that the magnitude of stretch influences the degree of HSF1 activation. HSF1 activation was blocked by gadolinium, but not by diltiazem, which indicates that the stretchinduced heat-shock response was mediated by SACs. An alternative perfusion system, the working heart, was used to determine whether perfusion alone was sufficient to activate HSF. Perfusion in the working mode, which is closer to physiological perfusion and does not use an apical drain or ventricular balloon, did not activate HSF; however, HSF could be activated by increasing afterload (aortic stenosis), which effectively added an acute stretch. Mechanical stresses, such as hemodynamic overload or mechanical stretch, alter the protein expression pattern in cardiac and skeletal muscles. For instance, Peterson and Lesch20 first reported that stretch could accelerate protein synthesis and amino acid transport. A similar phenomenon was observed in rat hearts subjected to high aortic pressures.21 Subsequent studies showed that the HSP72 gene and the “immediate early” (IE) genes, such as c-fos, c-myc, c-jun, JE, and Egr-1, were involved in the early response to mechanical stresses, followed by reexpression of fetal contractile protein genes, such as skeletal ␣-actin, -myosin heavy chain (MHC), and atrial natriuretic peptide (ANP) genes.9,22–25 How cells sense mechanical stimuli, transmit, and how this translates into gene expression is not understood. Increased sympathetic nervous activity and increased catecholamine levels have been suggested as potential mediators for induction of cardiac hypertrophy and IE gene expression.26,27 Since Vandenburgh and Kaufman28 reported that mechanical stretch without confounding factors caused increased protein synthesis in cultured skeletal muscle cells, however, many studies have demonstrated that mechanical stimuli alone can directly induce gene expression, including HSPs.9,10,12,15,23,24 In our study, we found that mechanical stretch and isovolumic contraction are sufficient to initiate the heat-shock response without neural and humoral factors. These observations extend our previous observations in the erythrocyteperfused rabbit heart, in which other factors, such as cytokines, could have had an effect, and demonstrate the activation of HSF via SACs in response to stretch.15 It has been postulated that SACs may function as a mechanotransducer between mechanical load and alterations in protein synthesis in cardiac hypertrophy.16,29 Mechanosensitive ion channels have been found in a broad variety of cell types.30 Activation of stretch-dependent channels results in cation influx, which is associated with gene expression and protein synthesis.16,31 We found that gadolinium decreased HSF activity by half and had a similar effect on HSP72 mRNA levels. Although other studies have shown a dose response,31 the small sample size may account for the lack of dose-dependent inhibition in the present study, or alternatively, a second activating pathway may be involved. Gadolinium 10 mol/L may have completely inhibited the SACs, even though it did not abolish HSF1 activation. A 5-fold increase in concentration had no further effect; thus, a second pathway may be involved in activation of HSF1 with stretch. Because it is known that gadolinium also blocks L-type calcium channels,31,32 we used diltiazem, a specific L-type calcium channel blocker, to exclude its effects, and it did not block the activation of HSF. Similar observations, ie, the noninvolvement of L-type calcium channels in the stretch response, were made by Laine et al33 and Komuro et al.23 We did not observe a negative inotropic effect with gadolinium in the concentrations used. Gadolinium does not affect the stretch-induced expression of IE genes in cultured neonatal cardiac myocytes.17 It has been shown that multiple second messenger factors, such as mitogen-activated protein kinases and protein kinase C, may be involved in the signal transductions for IE gene expression via the activation of serum response factor–P62TCF com- Chang et al Heat-Shock Factor and Stretch-Activated Channels 213 Figure 6. Comparison of HSF activity in nonperfused (A), working perfusion (B), Langendorff perfusion (C), Langendorff with gadolinium (D, 10 mol/L; E, 50 mol/L), Langendorff with diltiazem (F, 1 mol/L; G, 3 mol/L), and working perfusion (43°C for 13 minutes) (H). Abbreviations as in previous figures. Downloaded from http://circ.ahajournals.org/ by guest on June 12, 2017 plex.25 The activation of these pathways and induction of the IE genes lead to cardiac hypertrophy.30 Induction of HSPs is not per se associated with hypertrophy, nor does overexpression of the HSPs result in hypertrophy.34 Thus, the activation of the heat-shock response by stretch may involve pathways distinct from the hypertrophic response. A recent study showed that mitogen-activated protein kinase specific inhibitors did not affect stretch-induced HSF1 activation in vascular smooth muscle cells, and our previous work suggested that protein kinase C does not play an important role in stretch-induced HSP72 expression.12,15 There are several additional differences between our work and the studies of stretch-induced IE genes. Not only are we studying a different set of genes, but we also used the intact, perfused heart as a model. The use of isolated cardiac myocytes, although an excellent model, may eliminate or modulate extracellular signaling. Furthermore, we have focused on the adult heart, and responses in the neonatal heart, in which the induction of IE genes has primarily been studied, may be different. Thus, differences in induction of the heat-shock response and the IE genes may reflect differences in overall regulation and/or differences in models. Others have observed that volume overload could precondition the heart, and both this stretch-induced preconditioning and classic preconditioning were blocked by gadolinium.35 They suggested that volume overload, “stretch,” preconditioned the heart through the activation of SACs. Because brief cardiac ischemia/reperfusion results in the upregulation of HSP726 and short ischemia could lead to a transient cardiac dilation (stretching), we suggest that the activation of SACs may be part of the common pathway in the initiation of the cellular stress response in the heart. Certainly, other important factors are present in ischemia, such as denatured Figure 7. Northern blot of HSP72 mRNA present in Langendorff-perfused rat hearts with and without gadolinium. proteins, but stretch may be a factor, particularly in the nonischemic region of the ventricle. Previously, we reported that we did not detect a further increase in HSP72 mRNA after ischemia in erythrocyte-perfused hearts compared with control perfusions.6 We may have already maximally induced the heat-shock response. With the current understanding of cytokines, it may be that in this preparation, which used erythrocytes from cows in the slaughterhouse, cytokines may have had an additive effect; however, perfusion without stretch, but with the erythrocytes, did not induce HSP72.36 More recently, knowing the effect of stretch, we took measures, such as reduced balloon volume/LVEDP (6 mm Hg), to prevent stretch. This resulted in minimal activation of HSF in control hearts, and we were able to correlate changes in ATP during ischemia/reperfusion with the degree of activation of HSF.19 In conclusion, we demonstrate that stretch is sufficient to activate HSF1 and increase HSP72 mRNA expression and that SACs are involved in the activation of HSF1 and upregulation of HSP72 mRNA. This represents a novel pathway for the activation of HSF1. Although these results have implications for researchers using the isolated, perfused rat heart preparation, the overall implications are far greater. Stretch is a common phenomenon in both the normal and diseased cardiovascular system. Increased stretch occurs with hypertension, heart failure, and myocardial infarction, for Figure 8. Pmax at start and end of experiments. There were no significant differences in developed pressure for gadoliniumtreated, diltiazem-treated, and 2 working-mode hearts vs Langendorff-perfused control hearts. No difference in Pmax was found between start and end of experiment within each group. 214 Circulation July 10, 2001 example, and SACs may be an important signaling pathway for the induction of the stress response in these settings. Furthermore, the heart has higher levels of HSP72 than many other tissues (unpublished results), and this may be related to the repetitive stretch that occurs as part of normal cardiac function. Acknowledgments This study was supported by NHLBI grants HL-45257 (Dr Wasser) and HL-58515 (Dr Knowlton). We thank Dr Heinrich Taegtmeyer for advice on the working heart model and Feng Xu for technical assistance. References Downloaded from http://circ.ahajournals.org/ by guest on June 12, 2017 1. Knowlton AA. The role of heat shock proteins in the heart. J Mol Cell Cardiol. 1995;27:121–131. 2. Benjamin IJ, McMillan DR. Stress (heat shock) proteins: molecular chaperones in cardiovascular biology and disease. Circ Res. 1998;83: 117–132. 3. Beckmann RP, Mizzen LA, Welch WJ. Interaction of Hsp 70 with newly synthesized proteins. Science. 1990;248:850 – 854. 4. Cheng MY, Hartl FU, Martin J, et al. Mitochondrial heat-shock protein hsp60 is essential for assembly of proteins imported into yeast mitochondria. Nature. 1989;337:620 – 625. 5. Sorger PK. Heat shock factor and the heat shock response. Cell. 1991; 65:363–366. 6. Knowlton AA, Brecher P, Apstein CS. Rapid expression of heat shock protein in the rabbit after brief cardiac ischemia. J Clin Invest. 1991;87: 139 –147. 7. Iwaki K, Chi S-H, Dillmann WH, Mestril R. Induction of HSP70 in cultured rat neonatal cardiomyocytes by hypoxia and metabolic stress. Circulation. 1993;87:2023–2032. 8. Xu Q, Fawcett TW, Udelsman FR, et al. Activation of heat shock transcription factor 1 in rat aorta in response to high blood pressure. Hypertension. 1996;28:53–57. 9. Izumo S, Nadal-Ginard B, Mahdavi V. Protooncogene induction and reprogramming of cardiac gene expression produced by pressure overload. Proc Natl Acad Sci U S A. 1988;85:339 –343. 10. Delcayre C, Samuel J-L, Marotte F, et al. Synthesis of stress proteins in rat cardiac myocytes 2– 4 days after imposition of hemodynamic overload. J Clin Invest. 1988;82:460 – 468. 11. Neufer PD, Ordway GA, Hand GA, et al. Continuous contractile activity induces fiber type specific expression of HSP70 in skeletal muscle. Am J Physiol. 1996;271:C1828 –C1837. 12. Xu Q, Schett G, Li C, et al. Mechanical stress-induced heat shock protein 70 expression in vascular smooth muscle cells is regulated by Rac and Ras small G proteins but not mitogen-activated protein kinases. Circ Res. 2000;86:1122–1128. 13. Udelsman R, Blake MJ, Stagg CA, et al. Vascular heat shock protein expression in response to stress. J Clin Invest. 1993;91:465– 473. 14. Xu Q, Li D, Holbrook NJ, et al. Acute hypertension induces heat shock protein 70 gene expression in rat aorta. Circulation. 1995;92:1223–1229. 15. Knowlton AA, Eberli FR, Brecher P, et al. A single myocardial stretch or decreased systolic fiber shortening stimulates the expression of heat shock protein 70 in the isolated, erythrocyte-perfused rabbit heart. J Clin Invest. 1991;88:2018 –2025. 16. Kent RL, Hoober K, Cooper G. Load responsiveness of protein synthesis in adult mammalian myocardium: role of cardiac deformation linked to sodium influx. Circ Res. 1989;64:74 – 85. 17. Sadoshima J-I, Takahashi T, Jahn L, et al. Roles of mechano-sensitive ion channels, cytoskeleton, and contractile activity in stretch-induced immediate-early gene expression and hypertrophy of cardiac myocytes. Proc Natl Acad Sci U S A. 1992;89:9905–9909. 18. Sun L, Chang J, Kirchhoff SR, et al. Activation of HSF and selective increase in heat-shock proteins by acute dexamethasone treatment. Am J Physiol. 2000;278:H1091–H1097. 19. Chang J, Knowlton AA, Xu F, et al. Activation of heat shock response: relationship to energy metabolites: a 31P NMR study in isolated perfused rat hearts. Am J Physiol. 2001;280:H426 –H433. 20. Peterson MB, Lesch M. Protein synthesis and amino acid transport in the isolated rabbit right ventricular papillary muscle. Circ Res. 1972;31: 317–327. 21. Takala T. Protein synthesis in the isolated perfused rat heart. Basic Res Cardiol. 1981;76:44 – 61. 22. Komuro I, Kaida T, Shibazaki Y, et al. Stretching cardiac myocytes stimulates protooncogene expression. J Biol Chem. 1990;265:3595–3598. 23. Komuro I, Katoh Y, Kaida T, et al. Mechanical loading stimulates cell hypertrophy and specific gene expression in cultured rat cardiac myocytes. J Biol Chem. 1991;266:1265–1268. 24. Sadoshima J-I, Jahn L, Takahashi T, et al. Molecular characterization of the stretch-induced adaptation of cultured cardiac cells. J Biol Chem. 1992;267:10551–10560. 25. Sadoshima J, Izumo S. The cellular and molecular response of cardiac myocytes to mechanical stress. Annu Rev Physiol. 1997;59:551–571. 26. Moalic JM, Bauters C, Himbert D, et al. Phenylephrine, vasopressin and angiotensin II as determinants of proto-oncogene and heat-shock protein gene expression in adult rat heart and aorta. J Hypertens. 1989;7: 195–201. 27. Iwaki K, Sukhatme VP, Shubeita HE, et al. ␣- and -Adrenergic stimulation induces distinct patterns of immediate early gene expression in neonatal rat myocardial cells. J Biol Chem. 1990;265:13809 –13817. 28. Vandenburgh H, Kaufman S. In vitro model for stretch-induced hypertrophy of skeletal muscle. Science. 1979;203:265–268. 29. Bustamante JO, Ruknudin A, Sachs F. Stretch-activated channels in heart cells: relevance to cardiac hypertrophy. J Cardiovasc Pharmacol. 1991; 17(suppl 2):S110 –S113. 30. Morris CE. Mechanosensitive ion channels. Membr Biol. 1990;113: 93–107. 31. Yang X-C, Sachs F. Block of stretch-activated ion channels in Xenopus oocytes by gadolinium and calcium ions. Science. 1989;243:1068 –1071. 32. Hu H, Sachs F. Stretch-activated ion channels in the heart. J Mol Cell Cardiol. 1997;29:1511–1523. 33. Laine M, Id L, Vuolteenaho O, et al. Role of calcium in stretch-induced release and mRNA synthesis of natriuretic peptides in isolated rat atrium. Pflugers Arch. 1996;432:953–960. 34. Radford NB, Fina M, Benjamin IJ, et al. Cardioprotective effects of 70-kDa heat shock protein in transgenic mice. Proc Natl Acad Sci U S A. 1996;93:2339 –2342. 35. Ovize M, Kloner RA, Przyklenk K. Stretch preconditions canine myocardium. Am J Physiol. 1994;266:H137–H146. 36. Nakano M, Knowlton AA, Yokoyama T, et al. Tumor necrosis factor␣–induced expression of heat shock protein in adult feline cardiac myocytes. Am J Physiol. 1996;270:H1231–H1239. Activation of Heat-Shock Factor by Stretch-Activated Channels in Rat Hearts Jiang Chang, Jeremy S. Wasser, Richard N.M. Cornelussen and A.A. Knowlton Circulation. 2001;104:209-214 doi: 10.1161/01.CIR.104.2.209 Downloaded from http://circ.ahajournals.org/ by guest on June 12, 2017 Circulation is published by the American Heart Association, 7272 Greenville Avenue, Dallas, TX 75231 Copyright © 2001 American Heart Association, Inc. All rights reserved. Print ISSN: 0009-7322. Online ISSN: 1524-4539 The online version of this article, along with updated information and services, is located on the World Wide Web at: http://circ.ahajournals.org/content/104/2/209 Permissions: Requests for permissions to reproduce figures, tables, or portions of articles originally published in Circulation can be obtained via RightsLink, a service of the Copyright Clearance Center, not the Editorial Office. Once the online version of the published article for which permission is being requested is located, click Request Permissions in the middle column of the Web page under Services. Further information about this process is available in thePermissions and Rights Question and Answer document. Reprints: Information about reprints can be found online at: http://www.lww.com/reprints Subscriptions: Information about subscribing to Circulation is online at: http://circ.ahajournals.org//subscriptions/