Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Cytokinesis wikipedia , lookup
Hedgehog signaling pathway wikipedia , lookup
Organ-on-a-chip wikipedia , lookup
Signal transduction wikipedia , lookup
Histone acetylation and deacetylation wikipedia , lookup
Cellular differentiation wikipedia , lookup
List of types of proteins wikipedia , lookup
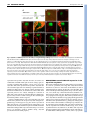
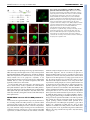
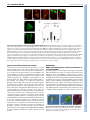
RESEARCH ARTICLE 1643 Development 136, 1643-1651 (2009) doi:10.1242/dev.032177 DORNRÖSCHEN is a direct target of the auxin response factor MONOPTEROS in the Arabidopsis embryo Melanie Cole1,*, John Chandler1,*, Dolf Weijers2, Bianca Jacobs1, Petra Comelli1 and Wolfgang Werr1,† DORNRÖSCHEN (DRN), which encodes a member of the AP2-type transcription factor family, contributes to auxin transport and perception in the Arabidopsis embryo. Live imaging performed with transcriptional or translational GFP fusions shows DRN to be activated in the apical cell after the first zygotic division, to act cell-autonomously and to be expressed in single cells extending laterally from the apical shoot stem-cell zone at the position of incipient leaf primordia. Here, we show that the Auxin response factor (ARF) MONOPTEROS (MP) directly controls DRN transcription in the tips of the embryonic cotyledons, which depends on the presence of canonical Auxin response elements (AuxREs), potential ARF-binding sites flanking the DRN transcription unit. Chromatin immunoprecipitation experiments show that MP binds in vivo to two AuxRE-spanning fragments in the DRN promoter, and that MP is required for expression of DRN in cotyledon tips. Hence, DRN represents a direct target of MP and functions downstream of MP in cotyledon development. INTRODUCTION Embryogenesis in plants establishes the basic body plan and stem cell populations for the generation of all post-embryonic organs. In Arabidopsis, embryo development is rapid and initially characterized by stereotypic cell divisions, which establish apicalbasal polarity and radial symmetry (Jürgens, 1992; Weijers and Jürgens, 2005). Based on seedling-lethal mutant analyses, the apical tier of four cells within the octant-stage embryo generates most of the cotyledon tissue and the shoot apical meristem (SAM) (Jürgens, 1995). The remaining cotyledon tissue, hypocotyl and root apical initials, the so-called central domain, derive from the lower tier of four cells, whereas the basal hypophysis region forms the quiescent centre and root cap initials (Scheres et al., 1994). Periclinal cell divisions superimpose a second axis of radial (inner-outer) symmetry towards the 16-cell stage. This radial symmetry is broken in the apical embryo domain by cotyledon specification, which marks the transition to bilateral symmetry, with a central-peripheral axis extending from the SAM outwards to the expanding cotyledons and, typical for dicots, a second, perpendicular plane of symmetry medially between both cotyledons (reviewed by Jenik et al., 2007; Chandler et al., 2008). Cotyledon defects in the seedling represent a deficiency in the acquisition of bilateral symmetry and are associated with mutations in genes encoding transcription factors, such as the CUP-SHAPED COTYLEDON (Aida et al., 1997; Aida et al., 1999), CLASS III HD-ZIP (Prigge et al., 2005) or DORNRÖSCHEN/DORNRÖSCHEN-LIKE (DRN/DRNL) (Chandler et al., 2007) genes or genes relating to auxin signalling such as PINOID (PID) (Bennett et al., 1995) or auxin transport (Friml et al., 2003). Auxin is also essential for specification of the 1 Institute of Developmental Biology, University of Cologne, Gyrhofstrasse 17, 50931 Cologne, Germany. 2Laboratory of Biochemistry, Wageningen University, Dreijenlaan 3, 6703 HA Wageningen, The Netherlands. *These authors contributed equally to this work Author for correspondence (e-mail: [email protected]) † Accepted 6 March 2009 basal embryo domain, where the MONOPTEROS (MP) or BODENLOS (BDL) gene products have a function in auxin perception and mutant mp or bdl embryos exhibit pronounced basal domain defects (Berleth and Jürgens, 1993; Hamann et al., 1999). Auxin response factors (ARFs) are transcription factors that regulate the expression of auxin response genes (Guilfoyle and Hagen, 2007; Guilfoyle et al., 1998). One of the best-studied ARF family members is MP (Hardtke and Berleth, 1998), mutant alleles of which show defects in vascular tissue and basal embryo development (Mayer et al., 1991; Berleth and Jürgens, 1993; Przemeck et al., 1996). ARFs bind to TGTCTC auxin response elements (AuxRE) in promoters of auxin-responsive genes (Ulmasov et al., 1997a) and function in combination with Aux/IAA (auxin/indole acetic acid) inhibitors, which dimerize with ARF activators in an auxin-regulated manner and hence modulate ARF activity (Ulmasov et al., 1997b). The Arabidopsis genome encodes 23 ARFs (Okushima et al., 2005), representing either transcriptional activators or repressors of auxin response genes, and 29 Aux/IAA proteins, which allows tremendous theoretical combinatorial possibilities affecting auxin response on the cellular level (Guilfoyle and Hagen, 2007). The Arabidopsis embryonic root meristem is initiated by the specification of a single cell, the hypophysis (Hamann et al., 1999), which crucially depends on the interaction of MP with its Aux/IAA inhibitor BDL (Hamann et al., 2002). Both MP and BDL genes function transiently in a small subdomain of the pro-embryo adjacent to the future hypophysis, where they promote transport of auxin and elicit a second non-cell-autonomous signal, which acts synergistically with auxin to specify hypophysis cell identity (Weijers et al., 2006). In the last few years, much research has addressed the regulation of ARF gene/protein expression, their function in plant growth or development, potential target genes, and the mechanisms by which they regulate target gene promoters. However, direct in vivo target genes served by individual ARFs remain unknown (Lau et al., 2008). Loss-of-function drn mutants have a bipartite phenotype affecting both the apical and the basal embryo domain. DRN was initially identified via a dominant drn-D allele isolated from an activationtagging screen in Arabidopsis, as having a shoot apical meristem function (Kirch et al., 2003). DRN encodes an AP2/ERF-type DEVELOPMENT KEY WORDS: Arabidopsis, Dornroeschen, Auxin response factor, Embryonic patterning, Target gene 1644 RESEARCH ARTICLE MATERIALS AND METHODS Construction of transcriptional/translational GFP fusions, mutagenesis of AuxREs and plant transformation DRN promoter activity depends on elements upstream and downstream of the protein-coding region (S. Niczyporuk, Diploma thesis, University of Cologne, 2003). The expression cassette combines 4864 bp upstream and 1378 bp downstream sequences flanking a unique SmaI/XmaI site. The GFP or chimeric DRN-GFP coding regions were inserted before transfer into the binary pGVTV-Asc-BAR vector (Überlacker and Werr, 1996). Mutagenesis of AuxREs was performed with the GeneEditor in vitro Site-Directed Mutagenesis System (Promega, Madison). The primers are shown in Table 1. Mutated fragments were assembled in three possible combinations to restore a functional expression cassette before insertion of the GFP marker gene. All chimeric gene constructs were inserted into the AscI site of pGVTV-Asc-BAR for Agrobacterium tumefaciens GV3101-mediated transformation (Bechtold and Pelletier, 1998). Plants were cultivated on soil in the greenhouse under long-day (16 hours light/8 hours dark) conditions at 22°C, and transgenic progeny were selected using resistance against the herbicide Basta. Genetic materials and genotyping EMS mutant mp alleles arf5-1 (N24605) and mp-U55 (N8147) (Mayer et al., 1991) and the T-DNA insertion allele mp-S319 (N21319) from the SALK collection (Alonso et al., 2003) were obtained from the NASC European Arabidopsis Stock Centre. mp mutant alleles were propagated as heterozygotes and DRN::GFP expression was analysed in the fraction (25%) of homozygous mp embryos obtained after selfing. Heterozygosity of the arf5-1 allele was confirmed by PCR on seedlings, whereas transmission of the mp-U55 allele solely relied on embryonic phenotypes after germination of F2 seeds. To obtain mp-U55 drn double mutants, heterozygous mp-S319/+ plants were crossed with homozygous drn/drn plants, both in Columbia ecotype, and transmission of the mp-S319 allele was confirmed by genotyping F1 progeny. Independent F2 lines were selected that contained a recombination event between the linked mp and drn loci, and progeny of the resulting mpS319/+, drn/drn lines were subjected to detailed phenotypic and genetic analyses in the F3 generation. The primers used for genotyping are included in Table 1. Paired Student’s t-tests were performed on select phenotypic data to assess statistical significance. Details of the construction of the MP-GFP fusion will be described elsewhere (Alexandra Schlereth and Dolf Weijers, unpublished). In brief, eGFP was amplified by PCR and inserted into the MlsI restriction site in the central region of the MP gene within an 8.5 kb genomic fragment that has been shown to complement the mp mutant (Weijers et al., 2006). The resulting MP-GFP construct (in pGreenII BASTA) was transformed into Col-0 Arabidopsis plants, and transgenic plants were selected based on GFP fluorescence in roots and embryos. Subsequently, one line was selected for a cross with the mp mutant and analysis of F2 seedlings demonstrated full complementation of the mutant. Confocal imaging and histology GFP expression was monitored using a Leica TCS SP2 confocal laserscanning microscope (CLSM). Earliest embryo stages were analysed still within the ovule, whereas later stages were isolated. Whole-mount seedlings were stained with the lipophilic styryl dye FM4-64 (4 μM) in 1% Tween, 130 mM NaCl, 10 mM phosphate buffer (pH 7.5). GFP was excited at 488 nm and emission analysed between 502 and 525 nm. All images were processed using Adobe Photoshop CS2 software. Chromatin immunoprecipitation and real-time PCR For crosslinking, nuclei were prepared from immature siliques (5g), essentially as described in (Conley et al., 1994), isolated nuclei were resuspended in nuclear isolation buffer (NIB: 50 mM Hepes pH 7.4, 5 mM MgCl2, 25 mM NaCl, 5% sucrose, 30% glycerol, 0.1% β-mercaptoethanol) supplemented with 20 μg/ml plant protease inhibitor cocktail (PPIC; Sigma P9599), 1 mM Na-orthovanadate and 1 mM NaF (NIBA). Crosslinking was performed with 1% formaldehyde for 5 minutes at room temperature in NIBA and stopped by pelleting and resuspending nuclei in NIBA containing 0.1 M glycine. Chromatin was fragmented by micrococcal nuclease (MNase) digestion (Wagschal et al., 2007) and nuclei were lysed in 10 mM TRIS pH 7.5, 1 mM EDTA, 0.5% SDS, 20 μg/ml PPIC. An aliquot of fragmented chromatin served as an input control for the PCR analysis and the remainder was subjected to immunoprecipitation (Mulholland et al., 2002). Magnetic bead-coupled anti-GFP antibodies (Miltenyi Biotec) were used to enrich for MP-GFP-containing chromatin fragments and decrosslinking of ChIP eluate and input control was preceded by a proteinase K treatment (1 μl/50 μl; Sigma P4580). DNA was purified with phenol/chloroform, precipitated with ethanol, washed with 70% ethanol and resuspended in 50 μl 10 mM Tris pH 7.5, 1 mM EDTA. The input control DEVELOPMENT transcription factor and maps to chromosome 1 within a larger chromosomal duplication also containing a paralogue DRN-LIKE (DRNL) at about 20 cM distance from DRN (Kirch et al., 2003). A strong drnl allele, drnl-2 or bcm1 (B-Class Modifier 1), also affects stamen development (Nag et al., 2007). Analysis of drn single mutants or double mutants between drn and a weak drnl-1 allele revealed a functional contribution to hypophysis specification in the globular/early heart-stage embryo; abnormal longitudinal cell divisions in the suspensor cells subtending the hypophysis led to a suspensor comprised of multiple parallel cell files. A proportion of drn drnl-1 double mutants phenocopies mp, a phenotype not frequently observed in drn or drnl single mutants, with an absence of the primary root and hypocotyl in the seedling. However, patterning defects in the apical embryo domain are reflected in a cotyledon phenotype, which is observed in up to 10% of drn single mutant seedlings and in 30% of drn drnl double mutant seedlings, with an additional 20% of drn drnl seedlings showing postgermination basal domain defects. Genetically, DRN and DRNL are highly redundant during embryonic patterning (Chandler et al., 2007). This conclusion is validated by protein-interaction studies showing that both DRN and DRNL interact with Class III HD-ZIP proteins such as PHAVOLUTA (PHV), PHABULOSA, REVOLUTA or CORONA and a basic helix-loop-helix-type transcription factor BIM1 (BES1-interacting myc-like) protein (Chandler et al., 2007; Chandler et al., 2009). Consistent with the assumption that these three classes of transcription factors form a higher-order protein complex, the phenotype of the drn phv bim1 triple mutant implicates all three genes in a single genetic pathway. DRN acts upstream of auxin signalling/perception or transport, as drn mutant embryos lack the DR5::GFP maximum in the hypophysis and upper suspensor cell and the subcellular polar distribution of PIN1-GFP is randomized and not oriented towards the hypophyseal cell in the basal domain of the globular embryo as in wild type (Chandler et al., 2007). A similar deficiency in directed auxin transport is observed in triple and quadruple mutant combinations of class III HD-ZIP genes (Prigge et al., 2005), encoding known interaction partners of DRN. Different lines of evidence thus support a connection between DRN and auxin transport or perception although overexpression experiments functionally link DRN (ESR1) or its paralogue DRNL (ESR2) also to cytokinins (Banno et al., 2001; Ikeda et al., 2006). DRN expression, as shown by RNA in situ hybridization, is dynamic throughout embryogenesis, beginning in the apical cell of the 2-cell embryo and gradually becoming restricted to the apical cell lineage (Kirch et al., 2003). Here, we describe a detailed spatial and temporal analysis of DRN expression in the Arabidopsis embryo using reporter constructs and show cell-autonomy of the DRN protein. Point mutations in canonical AuxREs show that upstream AuxREs are essential for DRN promoter activity in the emerging cotyledons. DRN expression in mp loss-of-function alleles and chromatin immunoprecipitation (ChIP) experiments both identify DRN as a direct target of MONOPTEROS in the embryo. Development 136 (10) was diluted 50-fold relative to the ChIP eluate and quantitative PCR analysis was performed using an Applied Biosystems 7300 Real-Time PCR System and Power SYBR Green PCR Master Mix (Applied Biosystems) and the relative amounts of different amplicons were determined by the ΔΔCT method and normalisation to the open reading frame (ORF) amplicon. All ChIP primers are included in Table 1 and the relative positions of the amplicons depicted in Fig. 3. RESULTS DRN::GFP expression pattern The DRN promoter construct comprises 4865 bp upstream from the DRN translation start and 1378 bp downstream of the stop codon (Kirch et al., 2003) and its activity depends on the combination of upstream and downstream elements (S. Niczyporuk, Diploma thesis, University of Cologne, 2003). In the chimeric DRN::GFP transcriptional reporter, the GFP-coding region exactly replaces the DRN ORF from the ATG to the stop codon whereas the GFP ORF was inserted in front of the DRN translation stop codon to obtain the DRN::DRN-GFP translational fusion (Fig. 1A); both constructs thus differ only by the presence or absence of the DRN coding region. Characteristic DRN::GFP expression patterns from successive embryonic stages are combined in Fig. 1B-N, and comparison of these patterns with those from the translational DRN-GFP fusion showed no substantial differences in expression on the cellular level, Table 1. Primers used in this study Primer Sequence (5⬘ to 3⬘) AuxRE mutagenesis Mut ARE-A check mut-a Mut ARE-B check mut-b Mut ARE-C check mut-c Mut ARE-D check mut-d Mut ARE-E check mut-e GAGAGATTAGGAGCCATTTGATCAGAGC AGAGTAATAGTAATAAAAGAGTAGTC TAATTGCTGCTTATTGGCTCATTCAGTCATTTGC CTATGGTTTCACATAGAGTTGTAACATTTC CATGAAATACATATATGGCTCATACATACGTTTAG GTAAGCATCGGGCGATATGGCATGATTATTTC CAAAAGGCAGTGTTTGGCTCTGTCTTCTCCAAC GGAAAATTCCTTTGTTCAGCAATGTTGGTATC GTAATTTAATCTACTGGCTCATATAAGTAGACAAC GTATGTTTCGTGGTTATTGCGTTGATGTAAC ChIP analysis RTnonAREf RTnonAREr RT-ARE-Af RT-ARE-Ar RT-ARE-Bf RT-ARE-Br RT-ARE-Cf RT-ARE-Cr RT-ORFf RT-ORFr RT-actin2f RT-actin2r CCATGGTGGTTTTGGTCTTTAC TGTATTCAAATTTGTTTGGTTGG TGCATACATATGAATCTGACCCTA GAAGAGGAGCAAAATGATCTGC CAGAATTAACGCAGGGGTTTTA AAACCATCTACCATGACCATGAC AACAAGCTTCTGATACATGAAATACA CGAAGAGGACATTTCAGGTTTG AACTCTTTCAACGGCTCATCAT ACGACTCGTTGTTTTCGTTTTC GTGTTGTTAGCAACTGGGATGA CTCTTCAGGAGCAATACGAAGC Genotyping ARF5f ARF5r (+LBb1) MP-S319f MP-S319r (+LBa1) DRNf (+SPM8) DRNr LBa1salk T-DNA border LBb1salk T-DNA border Spm8 GGCTATTGAGATATTTTGGC CATTGAGGATTGAAGAAGCT GATTTTCCAGATAATTCTGGA ATGAATATAGTTTCAGGTCTC ATGGAAAAAGCCTTGAGAAAC ATCATGATGAATATCTAGCTA TGGTTCACGTAGTGGGCCATCG GCGTGGACCGCTTGCTGCAACT GTTTTGGCCGACACTCCTTAC RESEARCH ARTICLE 1645 except that the GFP protein was cytosolic whereas the DRN-GFP fusion protein accumulated more locally and was mostly associated with the nuclear compartment. Importantly, no evidence was obtained for trafficking of the DRN-GFP protein into basal suspensor cells during embryogenesis. However, introduction of the DRN::DRN-GFP translational fusion transgene into the drn1 drnl double mutant background resulted in partial complementation of the basal domain embryo phenotype. Homozygous drn-1 drnl-1 mutant embryos have virtually complete (94%) penetrance of abnormal cell divisions in the basal embryo and subtending suspensor domain, which is reduced to 43% (38/89 embryos) in the presence of the DRN::DRN-GFP transgene; similarly, the penetrance of the cotyledon phenotype in the seedling is reduced from 50 to 7% in this complementation experiment. The DRN-GFP fusion protein therefore is at least partially functional, and the cellautonomous activity in the apical embryo domain is sufficient for significant rescue of basal domain defects. In consequence, aberrant cell divisions in the basal cell lineage of drn mutant embryos probably relate to a secondary non-cell-autonomous signal downstream of DRN. The DRN::GFP transcriptional reporter and the DRN::DRN-GFP translational fusion were expressed throughout the 2-, 4-, 8- or 16cell embryo proper (Fig. 1B-G). From the 32-cell stage onwards, DRN::GFP expression was no longer detected in the basal domain, which gives rise to hypocotyl and primary root (Fig. 1H,I), and two discrete expression maxima became evident in the apical domain of the late globular embryo, which mark the positions of the prospective cotyledons (Fig. 1J). During the late heart stage, both DRN::GFP and DRN::DRN-GFP expression was restricted to the tips of the cotyledons, with no expression observed in the sinus between the cotyledons or in the prospective SAM (Fig. 1K; data not shown). DRN expression was activated in the SAM during the torpedo stage (Fig. 1L,M). Subsequently, expression ceased in the cotyledon tips, whereas expression in the SAM continued during later embryonic stages. At the cellular level, DRN::GFP showed a dynamic expression pattern at the SAM periphery, where expression was occasionally observed to extend laterally into a single cell outside the central expression domain of the SAM. This expression was observed in single cells at either flank of the SAM, at positions where the first leaf primordium will be initiated (Fig. 1N), or marked positions of incipient leaf primordia in the germinating seedling (Fig. 1O). DRN::GFP expression persisted in the L1 layer of the developing leaf during early development (Fig. 1O). Point mutations in AuxREs alter DRN::GFP expression Differences in auxin transport or signalling/perception observed in drn mutant embryos compared to wild type (Chandler et al., 2007), suggested an analysis of functional contribution of five canonical TGTCTC AuxRE motifs, three of which are located in the DRN 4.8 kb upstream region, and two downstream of the coding sequence (Fig. 2A). Point mutations were introduced into each canonical AuxRE motif (TGTCTC to TGgCTC) and three classes of transgenic Arabidopsis plants were raised carrying either mutations in all five AuxRE motifs (abcde) or mutations in the three upstream (abc) or the two downstream motifs (de). This T-to-G mutation in the AuxRE motif has been shown to eliminate binding of ARFs (Ulmasov et al., 1999). Point mutations in AuxRE motifs had no effect on DRN expression during early stages of embryogenesis, based on data from at least three independent transgenic lines (Fig. 2C,D). However, at the torpedo (Fig. 2E) and early walking-stick stage (Fig. 2F), AuxRE mutations in the upstream motifs (abcde or abc) abolished DRN::GFP DEVELOPMENT DORNRÖSCHEN an in vivo target of MONOPTEROS 1646 RESEARCH ARTICLE Development 136 (10) expression in the tips of the cotyledons. Therefore, at least one, or a combination of AuxRE motifs upstream of the coding region, are essential to maintain DRN promoter activity in the tips of the cotyledons. DRN::GFP expression in the tips of the cotyledons also coincided with a high auxin concentration/perception maximum as monitored by DR5::GFP (Fig. 2B). By contrast, mutations in the downstream AuxREs alone (de) had little effect on the DRN::GFP expression pattern in the embryo, although the restriction of DRN::GFP activity into the apical domain during the globular stage appeared delayed relative to that in transgenic lines containing the non-mutated promoter construct (Fig. 2D). Canonical AuxRE motifs are not required for DRN expression maxima at the position of the incipient cotyledons during the late globular stage of embryogenesis (Fig. 2D) and the expression of DRN::GFP in single cells extending from the SAM periphery was similarly not affected by mutations in upstream AuxREs. Positional information relating to DRN expression is thus also perceived in the absence of functional canonical AuxREs during cotyledon specification and at the SAM periphery. MONOPTEROS controls DRN::GFP expression in the tips of the cotyledons Based on the AuxRE function in the DRN promoter, it was tempting to search for a potentially interacting ARF. Cotyledon and basal domain defects, which are characteristic for drn mutants, are shared with homozygous mp embryos. We therefore crossed the DRN::GFP marker into two mp mutant backgrounds; the mp-U55 allele (Mayer et al., 1991) and the arf5-1 T-DNA insertion allele (Okushima et al., 2005) from the SALK collection. In both mp backgrounds, which were propagated as heterozygotes, DRN::GFP expression in the tips of the cotyledons was abolished in segregating homozygous mutant embryos (25% of progeny) from the heart stage onwards, whereas other DRN::GFP expression domains e.g. during early embryogenesis or in the SAM, remained unaffected. Phenotypic mp embryos at the late torpedo stage showed no DRN::GFP expression in cotyledon tips, whereas expression in the SAM was unaffected (Fig. 3A-C). As the mp mutant often has cotyledon defects, the DRN::GFP expression pattern in mp mutant embryos was difficult to compare with wild type or AuxRE-mutated DRN promoter versions, where it pre-patterns DEVELOPMENT Fig. 1. Dynamics of DRN expression in the embryo and germinating seedlings. (A) Schematic of the DRN::GFP transcriptional and DRN::DRNGFP translational fusions. (B-D) DRN::GFP expression in the apical cell of the early embryo (B) and restriction to the apical cell lineage in 4-cell embryos (C) (subtending non-expressing suspensor cells are outlined in B and C) or 8-cell embryos (D). (E) Cell autonomy of the DRN-GFP fusion protein at the 8-cell stage. (F-I) Comparison of DRN::GFP and DRN::DRN-GFP expression patterns in early (F,G) and late (H,I) globular stage embryos. The focus of the DRN-GFP fusion protein into the apical hemisphere presumably relates to the stability of the chimeric protein relative to that of the GFP marker. (J,K) Focussing of DRN::GFP expression to the prospective cotyledons at the late globular (J) and early heart (K) stage. (L,M) Expression patterns of the transcriptional DRN::GFP (L) and translational DRN::DRN-GFP (M) fusion in torpedo-stage embryos. Note DRN promoter activity in the SAM, which is absent during the heart stage (compare with K). (N) Lateral expression of DRN::GFP in a single cell extending laterally from the L1 layer at the position of an incipient leaf primordium in the torpedo-stage embryo. Inset, close-up of the SAM region showing a single GFPexpressing cell at the lateral flank. (O) A similar occurrence in the vegetative SAM of a two-leaf stage seedling. The frontal cotyledon has been removed and the first two leaf primordia are emerging on both sides of the SAM. The large cells above the leaf primordia belong to the remaining cotyledon at the back. Note DRN::GFP activity in the L1 layer of the leaf primordia (O) or the cotyledons (N). Scale bars: 20 μm. DORNRÖSCHEN an in vivo target of MONOPTEROS RESEARCH ARTICLE 1647 cotyledon initiation. In phenotypic heart-stage mp mutant embryos, DRN::GFP expression was highest in the L1 layer, with expression also found throughout the SAM (compare Fig. 3D with 3E). Although DRN::GFP expression was confined to the L1 layer of the apical embryo domain and to the prospective cotyledons, it was downregulated in wild type irrespective of the presence of functional AuxREs; however, in the mp mutant background, DRN::GFP apparently remained active in the SAM. The lack of DRN::GFP expression in the cotyledon tips in the absence of MP activity exactly recapitulates the results obtained from point mutations in upstream AuxREs of the DRN promoter. Apparently, MP provides a unique ARF function essential for DRN transcription in the tips of the embryonic cotyledons, which is not masked by redundancy among other members of the ARF family of transcription factors. MONOPTEROS interacts with the DRN promoter in vivo To test whether the regulation of DRN expression by MP involves direct binding of MP to the DRN promoter, we performed ChIP experiments with a transgenic MP::MP-GFP line expressing a functional MP-GFP fusion protein driven from the MP promoter (Fig. 3F,G). Immature siliques of transgenic MP::MP-GFP lines containing heart-stage embryos were collected and used for ChIP experiments, as outlined in Materials and methods. Anti-GFP antibodies coupled to magnetic beads were used to precipitate MPbound DNA fragments, and the enrichment of upstream or downstream DRN promoter sequences via ChIP was analysed and quantified by real-time PCR. The position of the different PCR amplicons in the DRN upstream or coding region is indicated in Fig. 3H. The bar diagram (Fig. 3I) shows the relative levels of individual PCR amplicons obtained in two independent ChIP experiments according to triplicate real-time qPCR normalized to an amplicon located in the DRN ORF. A control fragment in the promoter upstream region lacking any AuxRE (nonARE) was amplified to a similar level to that of the most distal AuxRE A. By contrast, AuxRE B and AuxRE C, which are proximal to the DRN transcription start site were selectively enriched in independent ChIP experiments. Within individual experiments, the enrichment of AuxRE B (about 12-fold) exceeded that of AuxRE C (about 5fold), suggesting that binding of AuxRE B to MP may either be stronger or more frequent than to AuxRE C. The ChIP experiments thus confirm the physical interaction of MP to two promoter regions spanning canonical AuxRE motifs proximal to the transcription start in the DRN upstream promoter region. Taken together, the combination of ChIP with the effect of point mutations in AuxREs in the DRN promoter and the DRN::GFP expression pattern in the mp mutant background confirms that DRN is a direct target of MP in the tips of the cotyledons during embryogenesis. DEVELOPMENT Fig. 2. Functional contribution of AuxREs in the DRN upstream and downstream region. (A) Position of AuxREs upstream or downstream of the DRN coding region, which is replaced by GFP in the DRN::GFP construct. Canonical AuxREs are indicated by capital letters (A,B,C,D,E) in the wild type; mutated AuxREs are indicated by crosses in the individual constructs depicted beneath and denoted by lowercase letters. (B) DR5::GFP expression in late heart-stage embryos showing auxin concentration/perception maxima in the tips of the cotyledons and in the embryonic root. (C-F) Comparison of GFP expression obtained with the three AuxRE mutated constructs depicted in A relative to the non-mutated DRN::GFP transcriptional fusion at different stages of embryo development: (C) globular stage, (D) early heart stage, (E) late heart/early torpedo stage, (F) late torpedo stage. Note the absence of DRN::GFP expression in cotyledon tips later than heart stage (E or F) when upstream elements are mutated (constructs abc or abcde), which contrasts with the marking of the prospective cotyledons in D. 1648 RESEARCH ARTICLE Development 136 (10) Fig. 3. Molecular and genetic interactions between MP and DRN. (A-C) DRN::GFP expression pattern in wild-type embryos (A) as compared with in the mp-U55 (B) or arf5-1 (C) mutant backgrounds. Note GFP activity in the SAM in both wild-type and mutant embryos, but the absence of expression in the tips of cotyledons in both mp mutant backgrounds. (D,E) Downregulation of DRN::GFP in the SAM of early heart-stage wild-type embryos (D) as compared with a continuous stripe of DRN::GFP activity from the cotyledons through the SAM in mp-U55 mutant embryos (E). (F,G) MP::MP-GFP expression in early (F) or late (G) heart-stage embryos showing a broad expression domain in apical and basal regions, including the vasculature. (H) Schematic of the DRN gene showing the upstream region and transcriptional unit (box), AuxRE positions (ovals) and the position of PCR amplicons in the promoter or coding region. (I) Results of two independent ChIP experiments performed with nuclei from immature siliques containing heart-stage embryos. The relative levels of PCR amplicons are depicted, as estimated by real-time PCR and normalized to the DRN ORF amplicon. AuxRE-B and AuxRE-C, which lie proximal to the DRN transcription start site, are selectively amplified. By contrast, chromatin templates spanning the distal AuxRE-A or the noARE amplicons are present at levels similar to those for the DRN ORF. Bars indicate the variation in three independent real-time PCR replicates performed for each ChIP experiment. Scale bars: 20 μm. DISCUSSION DRN transcription pattern and the contribution of canonical AuxREs The DRN promoter is activated after the first zygotic division and DRN transcription in the apical cell coincides with acropetal auxin transport in the 2-cell Arabidopsis embryo (Friml et al., 2003), although the effect of point mutations in canonical AuxRE motifs in the DRN promoter does not suggest that transcription is controlled by auxin before the heart stage. Canonical AuxREs are not relevant for dynamic changes in DRN expression during the globular stage when DRN promoter activity is lost in the basal embryo domain or when expression maxima are established at the position of the prospective cotyledons. These results are consistent with the genetically defined role of DRN upstream of auxin transport or perception in the early Arabidopsis embryo (Chandler et al., 2007). However, they by no Fig. 4. Genetic interactions between mp and drn mutant alleles. (A) The mp basal domain defect, (B) the absence of the seedling root and presence of a single cotyledon and (C) the pin-like inflorescence phenotype. For frequencies see Table 2. DEVELOPMENT Genetic interaction between mp and drn To test for genetic interactions between mp and drn, we created double mutants using a weak mp-S319 allele from the SALK collection (Alonso et al., 2003). MP is located on chromosome 1 (27.6 cM) in the interval between DRN (19.2 cM) and DRNL (39.0 cM); this linkage necessitated a recombination event between drn and mp in order for mp to be propagated as a heterozygote in a drn mutant background. In a population segregating for mp but homozygous for drn, double mutant progeny should comprise a 25% fraction. The penetrance of seedling and inflorescence phenotypes was determined for progeny of six independent mp/MP drn/drn lines (Fig. 4; Table 2). The mp-S319 allele either showed a lowly penetrant embryonic pattern phenotype reflected in the absence of the basal seedling domain (Berleth and Jürgens, 1993) or a pin-like inflorescence phenotype (Przemeck et al., 1996), and together both phenotypes were fully penetrant, corresponding to the expected 25% fraction. In lines homozygous for drn and also segregating for mp, the penetrance of either the mp basal domain seedling (Fig. 4A,B) or inflorescence phenotype (Fig. 4C) was not statistically different to that of mp single mutants, and the frequency of the drn cotyledon phenotype was significantly reduced (P<0.001), thus confirming that the penetrance of characteristic drn cotyledon defects is dependent on MP function and that the mp phenotype is epistatic to that of drn. The absence of additivity of phenotypic penetrance or novel phenotypes together with the epistasis of mp over drn, supports the assumption that MP and DRN are in a linear pathway and that MP is a positive upstream activator of DRN. DORNRÖSCHEN an in vivo target of MONOPTEROS RESEARCH ARTICLE 1649 means exclude a contribution of non-canonical auxin response motifs to the regulation of early embryonic DRN expression, as it is still not clear how degenerate the consensus TGTCTC sequence can be to serve as a functional AuxRE in planta (Guilfoyle and Hagen, 2007). There is also no contribution of AuxREs to the activation of DRN transcription in the SAM at the late heart stage; DRN expression is still absent from the SAM when meristem markers such as WUSCHEL (WUS) (Mayer et al., 1998) and SHOOT MERISTEMLESS (STM) (Long and Barton, 1998) have already been activated. The late onset of DRN activity in the SAM only towards the end of the heart stage implies that DRN function may not directly contribute to SAM anlage. At the SAM periphery, the DRN::GFP marker reveals strikingly dynamic DRN promoter activity: from the group of 6-8 L1 layer cells permanently expressing DRN::GFP in the SAM centre, expression is often observed in single cells extending laterally from the flank of the stem cell zone at the position of the next incipient leaf primordium. Persisting DRN::GFP expression in the L1 layer of emerging leaf primordia makes it tempting to interpret these single DRN::GFP-expressing cells as evidence for anlagen of new leaf primordia, which in the close Arabidopsis relative Cardamine hirsuta trace back to as few as two cells (Barkoulas et al., 2008). The initiation of lateral organs is dependent on the polar transport of auxin into incipient primordia (Reinhardt et al., 2003), however, it remains to be determined whether this transport already occurs as early as the observed DRN::GFP expression in single cells lateral to the SAM in embryo or seedling. This lateral expansion of DRN::GFP transcription is also independent of canonical AuxREs, suggesting that positional information monitored by DRN promoter activity at the SAM periphery might not rely on auxin signalling. This situation is reminiscent of the late globular embryo when two DRN::GFP expression maxima are established at the positions of the prospective cotyledons despite mutations in upstream AuxREs (Fig. 2D, abc or abcde). However, functional AuxREs in the DRN promoter upstream region are indispensable for the maintenance of DRN::GFP expression in the tips of the cotyledons during the heart and torpedo stage of embryo development. The selectivity of individual DRN promoter element function on the cellular level is extremely striking; they regulate only a very specific aspect of DRN transcription in the embryo. Although DRN genetically acts upstream of auxin (Chandler et al., 2007) in the embryo, point mutations in AuxREs provide clear evidence for a feedback control via auxin signalling. The expression patterns of the promoter DRN::GFP and protein fusion DRN::DRN-GFP constructs were essentially comparable, thus providing no evidence for intercellular trafficking of the DRN protein, e.g. from the apical to the basal domain of the embryo. Therefore, DRN apparently acts cell-autonomously in the Arabidopsis embryo. The absence of a DR5::GFP maximum in the hypophyseal region and aberrant cell divisions in the root/suspensor associated with drn loss of function therefore either relate to defects in auxin signalling or must be explained by another non-cellautonomous signal downstream of DRN, as it is also discussed for mp mutants (Weijers et al., 2006). Molecular and genetic interactions Three complementary findings support the idea that DRN is a direct target of MP: the results of point mutations in AuxREs, the DRN::GFP expression pattern in an mp mutant background and ChIP experiments, which confirm the physical interaction of MP with two small promoter fragments containing AuxREs proximal to the DRN transcription start site. The strict dependence of DRN::GFP expression in the tips of the cotyledons on canonical AuxREs in the promoter upstream region and a functional MP allele is notable, considering that MP is a member of the Arabidopsis ARF gene family comprised of 23 members with inherent redundancy. A functional overlap between MP/ARF5 and NPH4/ARF7 during cotyledon development has been reported (Hardtke et al., 2004). However, this is incompatible with the absence of DRN::GFP activity in the tips of the cotyledons obtained in the mp-U55 (Mayer et al., 1991) and the arf5-1 (Okushima et al., 2005) alleles, which show that MP is unique in promoting transcriptional activity of the DRN promoter in the tips of the Total seedling phenotype penetrance (%) drn cotyledon phenotype penetrance (%) mp seedling phenotype penetrance (%) mp inflorescence phenotype penetrance (%) 1 2 3 4 5 6 7.04 13.27 9.55 12.63 13.91 10.40 11.23±2.62 1.53 2.81 2.59 2.55 3.08 2.17 2.46±0.54 5.51 10.46 6.96 10.08 10.83 8.23 8.68±1.96 21.59 15.32 18.52 14.29 13.46 17.62 16.80±3.04 1 2 3 4 5 6 13.68 8.60 8.93 11.06 6.31 14.39 10.50±3.14 0.50 0.27 0.19 0.00 0.08 0.26 0.22±0.17 13.18 8.33 8.74 11.06 6.23 14.13 10.35±3.15 9.89 14.17 17.00 11.13 15.40 13.30 13.48±2.65 9.86 8.80 11.53 9.95 9.54 9.44 9.85±0.92 0.12 0.10 0 0 0 0 0.04±0.06 Parental genotype Line drn drn/MP mp Mean±s.d. MP mp Mean±s.d. drn drn 1 2 3 4 5 6 Mean±s.d. The mp-S319 allele is propagated heterozygously. DEVELOPMENT Table 2. The penetrance of seedling and inflorescence phenotypes obtained in six independent mp/MP drn/drn lines in comparison to frequencies characteristic for mp and drn single mutants cotyledons, and therefore that other ARF family members, including NPH4/ARF7, cannot compensate for the loss of MP function with respect to DRN::GFP transcription in a few apical cells of the emerging cotyledons in heart- or torpedo-stage embryos. As striking as the strict dependence of DRN::GFP transcription on MP in the tips of the cotyledons is the fact that this is very local, comprising only a small subfraction of the DRN (Fig. 1) or MP (Hardtke and Berleth, 1998) expression domains in the embryo. Apparently, MP is not sufficient alone to activate DRN::GFP outside the apical tip of the cotyledons. It is known that ARFs are under control of Aux/IAA proteins (Ulmasov et al., 1997b); for example, MP binds to BDL and both enter the nuclear compartment (Weijers et al., 2006). MP might bind DNA targets (Lau et al., 2008) including AuxREs in the DRN promoter more widely as a heterodimer with BDL. However, to effect transcriptional activation, MP has to be released from BDL repression, which requires the nuclear SCFTIR1 complex and high auxin concentrations (Mockaitis and Estelle, 2008; Tan et al., 2007), as can be reflected in DR5::GFP activity in the tips of the cotyledons (see Fig. 2B). Apart from MP modulation by BDL and auxin, the TOPLESS co-repressor restricts MP activity during embryogenesis (Szemenyei et al., 2008) or heterodimerization with other ARFs might result in altered DNA target-site specificity (Kim et al., 1997; Ulmasov et al., 1997a). Although MP is capable of forming heterodimers with NPH4/ARF7 (Hardtke et al., 2004), the overlap of the MP or NPH4 expression domains throughout the embryonic vasculature cannot explain the local response of DRN::GFP in the tips of the cotyledons. Possibly, additional cis or trans elements have to converge for a positive transcriptional read-out from the DRN promoter with the differential response of AuxREs. However, the specific loss of DRN::GFP activity at the tips of the cotyledons in mp single mutant backgrounds unequivocally demonstrates that MP is unique among the ARF gene family in controlling DRN transcription. According to point mutations and ChIP experiments, at least two AuxREs proximal to the DRN transcriptions start site are potential DNA target sites for the MP protein. AuxREs have been repeatedly identified in available genome sequences and functionally supported by transcriptome analyses, but the descriptions of specific auxindependent events that depend on protein-DNA interactions between an individual member of the ARF family and specific target gene promoters have remained elusive in vivo (Guilfoyle and Hagen, 2007; Lau et al., 2008), although NPH4/ARF7 has been shown in vitro to interact with AuxRE-containing fragments of the LATERAL ORGAN BOUNDARIES DOMAIN16 (LBD) or LBD29 gene promoters (Okushima et al., 2007). The identification of DRN as a bona fide target of MP, the unique role of MP in controlling DRN expression at the tip of the embryonic cotyledons, possibly in response to auxin, and the suitability of the Arabidopsis embryo for live imaging should now facilitate studies on the cellular and molecular level. The genetic epistasis of the mp mutant phenotype over that of drn, together with the absence of any additive phenotypic effects in either penetrance or novel phenotypes in the double mutant supports a genetic hierarchy whereby MP is a positive regulator of DRN (Avery and Wasserman, 1992). This genetic interaction is somewhat difficult to explain relative to transcriptional control exerted by MP at the DRN promoter, which affects only a single and rather late aspect of DRN expression involving only a minority of MP-expressing cells. More penetrant phenotypes in drn and mp single mutants are basal domain defects; similarly to mp or bdl mutants, drn mutant embryos lack the DR5::GFP auxin concentration/perception maximum in the hypophysis (Chandler et al., 2007). Concerning basal domain defects, Development 136 (10) MP and DRN act in different functional hierarchies; whereas mp mutant embryos exhibit reduced PIN1::PIN-GFP expression levels (Weijers et al., 2006), drn embryos show alterations in the intracellular compartmentation of the PIN1-GFP fusion protein in terms of membrane polarity compared with wild type (Chandler et al., 2007). By contrast, mono- or polycotyledony or cotyledon fusion, which are characteristic for drn mutant seedlings and which are reduced in drn mp double mutants, reflect aberrant specification of cells in the apical embryo domain. The dependence of DRN on MP in terms of embryonic patterning might be functionally addressed in more depth by ectopic expression of DRN in the mp mutant background. This, however, is difficult to perform and of questionable relevance for several reasons: firstly, 35S::DRN transgenic plants are inhibited in shoot formation (Banno et al., 2001) and the dominant drn-1D allele results in a phenotype only late in seedling development (Kirch et al., 2003) and only extremely rarely exhibits cotyledon defects (Chandler et al., 2007). Additionally, redundancy between DRN and DRNL might mask part of the interaction between MP and DRN; drn drnl double mutants exhibit cotyledon defects at 50% penetrance, but it is still unknown whether DRNL can substitute for DRN function or whether the DRNL promoter is also under control of MP. Finally, MP maps on chromosome 1 within the 20 cM interval between DRN and DRNL and so is tightly linked to both, and we have so far failed to establish a triple mutant. In the mp background, the DRN::GFP expression pattern is altered such that, in contrast to wild type, transcription in the future SAM region is not downregulated between the prospective cotyledons (compare Fig. 3D with 3E). Similar ectopic DRN promoter activity is not observed with point mutations in canonical AuxREs in the DRN promoter; by contrast, DRN::GFP activity initially marks the prospective cotyledons but is lost in both mp mutant embryos and following mutation of upstream DRN promoter ARFs. This unique role of MP at the DRN promoter and at the tips of the cotyledons coincides with a local maximum of YUCCA4 monooxygenase (Cheng et al., 2007), and thus local auxin biosynthesis, together with auxin maxima or response peak as monitored via DR5::GFP (see Fig. 2B). Exactly how the AP2-type transcription factor DRN functionally integrates into the embryonic patterning programme remains elusive; however, the deficiencies in PIN1-GFP polarization in drn mutants strongly implicate auxin transport, which might be initially direct auxin into cotyledon anlagen similarly to into incipient leaf primordia (Reinhardt et al., 2003) and contribute to an MP-dependent auxin maximum at the apical tip of the developing cotyledons. Conceivably, an auxin feedback loop here could control cotyledon outgrowth but also cause cellular ambiguities in the apical embryo domain, which is manifest in cotyledon defects in drn seedlings. By contrast, the epistasis of the mp phenotype over that of drn could reside in an altered cellular competence for the functional separation of cotyledons or their subsequent morphogenesis, which is implied by the altered DRN::GFP expression in mp mutants. Additional contributions to cotyledon development are suggested by cotyledon defects in pid (Bennett et al., 1995), polar transport double mutants such as pin4 pin7 (Friml et al., 2003) or the cuc mutants (Aida et al., 1997; Aida et al., 1999), most of these being lowly penetrant. In summary, genetic and molecular evidence identify DRN as a direct target gene of MONOPTEROS in the tips of the embryonic cotyledons in the Arabidopsis embryo. All data merge on the conclusion that two canonical AuxREs proximal to the DRN transcription start site serve as in vivo targets for MP, which now facilitates molecular studies on the specificity of auxin signalling on DEVELOPMENT 1650 RESEARCH ARTICLE the cellular, individual ARF and target gene level. DRN, encoding an AP2 family transcription factor, is a particularly intriguing target as it apparently acts upstream and downstream of auxin, is associated with the apical cell fate after the first zygotic division and perceives positional information at the flank of the stem-cell population, which relates to the initiation of new lateral organs. This work was funded by the Deutsche Forschungsgemeinschaft through SFB 572 (M.C., J.C., B.J., P.C. and W.W.) and by the Dutch Organisation for Scientific Research (NWO; ALW-VIDI grant 864.06.012 to D.W.). References Aida, M., Ishida, T., Fukaki, H., Fujisawa, H. and Tasaka, M. (1997). Genes involved in organ separation in Arabidopsis: an analysis of the cup-shaped cotyledon mutant. Plant Cell 9, 841-857. Aida, M., Ishida, T. and Tasaka, M. (1999). Shoot apical meristem and cotyledon formation during Arabidopsis embryogenesis: interaction among the CUPSHAPED COTYLEDON and SHOOT MERISTEMLESS genes. Development 126, 1563-1570. Alonso, J. M., Stepanova, A. N., Leisse, T. J., Kim, C. J., Chen, H., Shinn, P., Stevenson, D. K., Zimmerman, J., Barajas, P., Cheuk, R. et al. (2003). Genome-wide insertional mutagenesis of Arabidopsis thaliana. Science 301, 653-657. Avery, L. and Wasserman, S. (1992). Ordering gene function: the interpretation of epistasis in regulatory hierarchies. Trends Genet. 8, 312-316. Banno, H., Ikeda, Y., Niu, Q. W. and Chua, N. H. (2001). Overexpression of Arabidopsis ESR1 induces initiation of shoot regeneration. Plant Cell 13, 26092618. Barkoulas, M., Hay, A., Kougioumoutzi, E. and Tsiantis, M. (2008). A developmental framework for dissected leaf formation in the Arabidopsis relative Cardamine hirsuta. Nat. Genet. 40, 1136-1141. Bechtold, N. and Pelletier, G. (1998). In planta Agrobacterium-mediated transformation of adult Arabidopsis thaliana plants by vacuum infiltration. Methods Mol. Biol. 82, 259-266. Bennett, S. R. M., Alvarez, J., Bossinger, G. and Smyth, D. R. (1995). Morphogenesis in pinoid mutants of Arabidopsis thaliana. Plant J. 8, 505-520. Berleth, T. and Jürgens, G. (1993). The role of the monopteros gene in organising the basal body region of the Arabidopsis embryo. Development 118, 575-587. Chandler, J. W., Cole, M., Flier, A., Grewe, B. and Werr, W. (2007). The AP2 transcription factors DORNRÖSCHEN and DORNRÖSCHEN-LIKE redundantly control Arabidopsis embryo patterning via interaction with PHAVOLUTA. Development 134, 1653-1662. Chandler, J. W., Nardmann, J. and Werr, W. (2008). Plant development revolves around axes. Trends Plant Sci. 13, 78-84. Chandler, J. W., Cole, M., Flier, A. and W. W. (2009). BIM1, a bHLH protein involved in brassinosteroid signalling, controls Arabidopsis embryonic patterning via interaction with DORNROESCHEN and DORNROESCHEN-LIKE. Plant Mol. Biol. 69, 57-68. Cheng, Y., Dai, X. and Zhao, Y. (2007). Auxin synthesized by the YUCCA flavin monooxygenases is essential for embryogenesis and leaf formation in Arabidopsis. Plant Cell 19, 2430-2439. Conley, T. R., Park, S. C., Kwon, H. B., Peng, H. P. and Shih, M. C. (1994). Characterization of cis-acting elements in light regulation of the nuclear gene encoding the A subunit of chloroplast isozymes of glyceraldehyde-3-phosphate dehydrogenase from Arabidopsis thaliana. Mol. Cell. Biol. 14, 2525-2533. Friml, J., Vieten, A., Sauer, M., Weijers, D., Schwarz, H., Hamann, T., Offringa, R. and Jürgens, G. (2003). Efflux-dependent auxin gradients establish the apical-basal axis of Arabidopsis. Nature 426, 147-153. Guilfoyle, T. J. and Hagen, G. (2007). Auxin response factors. Curr. Opin. Plant Biol. 10, 453-460. Guilfoyle, T. J., Ulmasov, T. and Hagen, G. (1998). The ARF family of transcription factors and their role in plant hormone-responsive transcription. Cell Mol. Life Sci. 54, 619-627. Hamann, T., Mayer, U. and Jürgens, G. (1999). The auxin-insensitive bodenlos mutation affects primary root formation and apical-basal patterning in the Arabidopsis embryo. Development 126, 1387-1395. Hamann, T., Benkova, E., Baurle, I., Kientz, M. and Jürgens, G. (2002). The Arabidopsis BODENLOS gene encodes an auxin response protein inhibiting MONOPTEROS-mediated embryo patterning. Genes Dev. 16, 1610-1615. Hardtke, C. S. and Berleth, T. (1998). The Arabidopsis gene MONOPTEROS encodes a transcription factor mediating embryo axis formation and vascular development. EMBO J. 17, 1405-1411. Hardtke, C. S., Ckurshumova, W., Vidaurre, D. P., Singh, S. A., Stamatiou, G., Tiwari, S. B., Hagen, G., Guilfoyle, T. J. and Berleth, T. (2004). Overlapping and non-redundant functions of the Arabidopsis auxin response factors MONOPTEROS and NONPHOTOTROPIC HYPOCOTYL 4. Development 131, 1089-1100. RESEARCH ARTICLE 1651 Ikeda, Y., Banno, H., Niu, Q. W., Howell, S. H. and Chua, N. H. (2006). The ENHANCER OF SHOOT REGENERATION 2 gene in Arabidopsis regulates CUPSHAPED COTYLEDON 1 at the transcriptional level and controls cotyledon development. Plant Cell Physiol. 47, 1443-1456. Jenik, P. D., Gillmor, S. and Lukowitz, W. (2007). Embryonic patterning in Arabidopsis thaliana. Annu. Rev. Cell Dev. Biol. 23, 207-236. Jürgens, G. (1992). Pattern formation in the flowering plant embryo. Curr. Opin. Genet. Dev. 2, 567-570. Jürgens, G. (1995). Axis formation in plant embryogenesis: cues and clues. Cell 81, 467-470. Kim, J., Harter, K. and Theologis, A. (1997). Protein-protein interactions among the Aux/IAA proteins. Proc. Natl. Acad. Sci. USA 94, 11786-11791. Kirch, T., Simon, R., Grunewald, M. and Werr, W. (2003). The DORNRÖSCHEN/ENHANCER OF SHOOT REGENERATION1 Gene of Arabidopsis acts in the control of meristem cell fate and lateral organ development. Plant Cell 15, 694-705. Lau, S., Jürgens, G. and De Smet, I. (2008). The evolving complexity of the auxin pathway. Plant Cell 20, 1738-1746. Long, J. A. and Barton, M. K. (1998). The development of apical embryonic pattern in Arabidopsis. Development 125, 3027-3035. Mayer, K. F., Schoof, H., Haecker, A., Lenhard, M., Jürgens, G. and Laux, T. (1998). Role of WUSCHEL in regulating stem cell fate in the Arabidopsis shoot meristem. Cell 95, 805-815. Mayer, U., Torres Ruiz, R. A., Berleth, T., Miséra, S. and Jürgens, G. (1991). Mutations affecting body organization in the Arabidopsis embryo. Nature 353, 402-407. Mockaitis, K. and Estelle, M. (2008). Auxin receptors and plant development: a new signaling paradigm. Annu. Rev. Cell Dev. Biol. 24, 55-80. Mulholland, D. J., Cheng, H., Reid, K., Rennie, P. S. and Nelson, C. C. (2002). The androgen receptor can promote β-catenin nuclear translocation independently of adenomatous polyposis coli. J. Biol. Chem. 277, 17933-17943. Nag, A., Yang, Y. and Jack, T. (2007). DORNRÖSCHEN-LIKE, an AP2 gene, is necessary for stamen emergence in Arabidopsis. Plant Mol. Biol. 65, 219-232. Okushima, Y., Overvoorde, P. J., Arima, K., Alonso, J. M., Chan, A., Chang, C., Ecker, J. R., Hughes, B., Lui, A., Nguyen, D. et al. (2005). Functional genomic analysis of the AUXIN RESPONSE FACTOR gene family members in Arabidopsis thaliana: unique and overlapping functions of ARF7 and ARF19. Plant Cell 17, 444-463. Okushima, Y., Fukaki, H., Onoda, M., Theologis, A. and Tasaka, M. (2007). ARF7 and ARF19 regulate lateral root formation via direct activation of LBD/ASL genes in Arabidopsis. Plant Cell 19, 118-130. Prigge, M. J., Otsuga, D., Alonso, J. M., Ecker, J. R., Drews, G. N. and Clark, S. E. (2005). Class III homeodomain-leucine zipper gene family members have overlapping, antagonistic, and distinct roles in Arabidopsis development. Plant Cell 17, 61-76. Przemeck, G. K., Mattsson, J., Hardtke, C. S., Sung, Z. R. and Berleth, T. (1996). Studies on the role of the Arabidopsis gene MONOPTEROS in vascular development and plant cell axialization. Planta 200, 229-237. Reinhardt, D., Pesce, E. R., Stieger, P., Mandel, T., Baltensperger, K., Bennett, M., Traas, J., Friml, J. and Kuhlemeier, C. (2003). Regulation of phyllotaxis by polar auxin transport. Nature 426, 255-260. Scheres, B., Wolkenfeldt, H., Willemsen, V., Terlouw, M., Lawson, E., Dean, C. and Weisbeek, P. (1994). Embryonic origin of the Arabidopsis primary root meristem initials. Development 120 2475-2487. Szemenyei, H., Hannon, M. and Long, J. A. (2008). TOPLESS mediates auxindependent transcriptional repression during Arabidopsis embryogenesis. Science 319, 1384-1386. Tan, X., Calderon-Villalobos, L. I., Sharon, M., Zheng, C., Robinson, C. V., Estelle, M. and Zheng, N. (2007). Mechanism of auxin perception by the TIR1 ubiquitin ligase. Nature 446, 640-645. Überlacker, B. and Werr, W. (1996). Vectore with rare-cutter restriction enzyme sites for expression of open reading frames in transgenic plants. Mol. Breed. 2, 293-295. Ulmasov, T., Hagen, G. and Guilfoyle, T. J. (1997a). ARF1: a transcription factor that binds to auxin response elements. Science 276, 1865-1868. Ulmasov, T., Murfett, J., Hagen, G. and Guilfoyle, T. J. (1997b). Aux/IAA proteins repress expression of reporter genes containing catural and highly active synthetic auxin response elements. Plant Cell 9, 1963-1971. Ulmasov, T., Hagen, G. and Guilfoyle, T. J. (1999). Activation and repression of transcription by auxin-response factors. Proc. Natl. Acad. Sci. USA 96, 58445849. Wagschal, A., Delaval, K., Pannetier, M., Arnaud, P. and Feil, R. (2007). Chromatin immunoprecipitation (ChIP) on unfixed chromatin from cells and tissues to analyze histone modifications. CSH Protoc. 3, doi:10.1101/pdb.prot4767. Weijers, D. and Jurgens, G. (2005). Auxin and embryo axis formation: the ends in sight? Curr. Opin. Plant Biol. 8, 32-37. Weijers, D., Schlereth, A., Ehrismann, J. S., Schwank, G., Kientz, M. and Jürgens, G. (2006). Auxin triggers transient local signaling for cell specification in Arabidopsis embryogenesis. Dev. Cell 10, 265-270. DEVELOPMENT DORNRÖSCHEN an in vivo target of MONOPTEROS