Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Site-specific recombinase technology wikipedia , lookup
Epigenetics of neurodegenerative diseases wikipedia , lookup
Microevolution wikipedia , lookup
Birth defect wikipedia , lookup
Saethre–Chotzen syndrome wikipedia , lookup
Oncogenomics wikipedia , lookup
Neuronal ceroid lipofuscinosis wikipedia , lookup
DiGeorge syndrome wikipedia , lookup
Frameshift mutation wikipedia , lookup
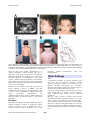
Atlas of Genetics and Cytogenetics in Oncology and Haematology OPEN ACCESS JOURNAL AT INIST-CNRS Cancer Prone Disease Section Review Noonan syndrome Marco Tartaglia, Bruce D Gelb Dipartimento di Biologia Cellulare e Neuroscienze, Istituto Superiore di Sanità, Rome, Italy (MT, BDG) Published in Atlas Database: June 2005 Online updated version: http://AtlasGeneticsOncology.org/Kprones/NoonanID10085.html DOI: 10.4267/2042/38259 This work is licensed under a Creative Commons Attribution-Noncommercial-No Derivative Works 2.0 France Licence. © 2005 Atlas of Genetics and Cytogenetics in Oncology and Haematology multiple skeletal defects (spine and chest), webbed neck, mental retardation, cryptorchidism and bleeding diathesis. Identity Alias Male Turner syndrome Pseudo-Turner syndrome Inheritance Noonan syndrome is an autosomal dominant disorder. Rare cases with parental consanguinity have been described, but it is not clear that these represent true instances of autosomal recessive inheritance. Like many autosomal dominant disorders, a significant, but not precisely determined, percentage of cases represent de novo mutagenesis. The prevalence of Noonan syndrome has not been determined accurately to date. Most authors cite the figure of 1 in 1,000-2,500 live births. However, that estimate was not based on a population study. Fetal loss occurs for Noonan syndrome so disease incidence is higher than prevalence, but no estimate of the magnitude of this discrepancy is available. Neoplastic risk Children with Noonan syndrome are predisposed to malignancies, juvenile myelomonocytic leukemia (JMML) most commonly. JMML, formerly termed juvenile chronic myeloid leukemia or chronic myelomonocytic leukemia, is a myeloproliferative/ myelodysplastic disorder of childhood characterized by excessive proliferation of immature and mature myelomonocytic cells that originate from a pluripotent stem cell. In childhood, JMML accounts for approximately 30% of cases of myelodysplastic and myeloproliferative syndromes and 2% of leukemias. It typically presents in infancy and early childhood, and is often lethal. Recent studies have provided strong evidence that hypersensitivity to granulocytemacrophage colony-stimulating factor (GM-CSF), due to a selective inability to down-regulate the RAS/MAPK cascade, plays a central role in the clonal cell growth characteristic of JMML. In approximately 15-30% of JMML cases, the pathological activation of the RAS/MAPK cascade results from oncogenic NRAS or KRAS2 mutations that specifically affect GTP hydrolysis, leading to the accumulation of RAS in the GTP-bound active conformation. JMML has been reported in children with neurofibromatosis type 1 (NF1), an autosomal dominant disorder resulting from germline loss-of-function mutations of the NF1 tumor suppressor gene. In children with NF1 and JMML, the proliferative advantage of the leukemic cells results from a second hit, the somatic loss or inactivation of the normal NF1 allele. Clinics Phenotype and clinics Noonan syndrome is a clinically variable developmental disorder defined by short stature, facial dysmorphism and a wide spectrum of congenital heart defects. The distinctive facial features consist of a broad forehead, hypertelorism, down-slanting palpebral fissures, ptosis, high-arched palate and low-set, posteriorly rotated ears. Cardiovascular abnormalities, primarily pulmonic stenosis and hypertrophic cardiomyopathy, are present in up to 85% of affected individuals. Additional relatively frequent features are Atlas Genet Cytogenet Oncol Haematol. 2005; 9(4) 345 Noonan syndrome Tartaglia M, Gelb BD Clinical features in Noonan syndrome. (A) Prenatal echography showing nuchal cystic hygroma; (B) dysmorphic facial features; (C) pectus deformities and cubitus valgus; (D) webbed neck; (E) schematic representation of major cardiac defects: 1, pulmonic stenosis; 2, hypertrophic cardiomyopathy; 3, atrial septal defects; 4, ventricular septal defects; 5, patent ductus arteriosus. (Figures kindly provided by G. Zampino, MD, Università Cattolica del Sacro Cuore, Rome, Italy). Since the NF1 gene product, neurofibromin, is a negative modulator of RAS function, this loss is associated with RAS hyperactivity. Remarkably, deregulated RAS function appears to be restricted to GM-CSF signaling in hematopoietic cells. More recently, somatic activating mutations in the PTPN11 gene, which encodes the protein tyrosine phosphatase (PTP) SHP-2, have been documented in approximately 35% of cases with JMML. prognosis without transplantation. stem cell Other findings Note A significant percentage of Noonan syndrome cases arise from de novo PTPN11 mutations. Among fourteen informative families, each consisting of an affected individual heterozygous for a PTPN11 mutation and unaffected parents, the paternal origin of mutation has been demonstrated in all cases. Consistent with this finding, advanced paternal age was noted among fathers of sporadic Noonan syndrome cases, compared with fathers of the reference population. Moreover, when a parent is affected with Noonan syndrome and harbors a PTPN11 mutation, a sex-ratio bias is operative for offspring who inherit the defect. This bias favors males by a factor of 2:1. The available data point to this bias being attributable to sex-specific developmental effects of PTPN11 mutations that favor survival of affected male embryos compared to female ones. Mutations in the PTPN11, RAS and NF1 genes are largely mutually exclusive in JMML, and their combined prevalence accounts for approximately 85% of cases. Acute lymphoblastic leukemia and solid tumors, particularly neuroblastoma and rhabdomyosarcoma, have also been documented with a relatively higher prevalence respect to the general population. Evolution In children with Noonan syndrome JMML may regress without treatment or follow an aggressive clinical course. By contrast, cases of JMML that arise in children without Noonan syndrome have a poor Atlas Genet Cytogenet Oncol Haematol. 2005; 9(4) hematopoietic 346 Noonan syndrome Tartaglia M, Gelb BD domain also interacts with the PTP domain using a separate site. As these subdomains show negative cooperativity, the N-SH2 domain functions as an intramolecular switch controlling SHP-2 catalytic activation. Specifically, the N-SH2 domain interacts with the PTP domain basally, blocking the catalytic site. Binding of the N-SH2 phosphopeptide-binding site to the phosphotyrosyl ligand promotes a conformational change of the domain that weakens the auto-inhibiting intramolecular interaction, making the catalytic site available to substrate, thereby activating the phosphatase. Expression: Widely expressed in both embryonic and adult tissues. Localisation: Cytoplasmic. It binds to activated cell surface receptors, cell adhesion molecules and scaffolding adapters. Phosphorylation of two tyrosine residues at the C-terminus by activated tyrosine kinase receptors creates binding sites for other SH2 domaincontaining signal transducers. Function: SHP-2 functions as a intracellular signal transducer. It positively modulates signal flow in most circumstances, but can also function as negative regulator depending upon its binding partner and interactions with downstream signaling networks. SHP2 positively controls the activation of the RAS/MAPK cascade induced by several growth factors, and negatively regulates JAK/STAT signaling. In most cases, SHP-2's function in intracellular signaling appears to be immediately proximal to activated receptors and upstream to RAS. The mechanisms of SHP-2's action and its physiological substrates are still poorly defined. However, both membrane translocation and PTPase activity are required for SHP-2 function. SHP-2 is required during development. Embryos nullizygous for Shp-2 have defects in gastrulation and mesodermal patterning resulting in severe abnormalities in axial and paraxial mesodermal structures. Shp-2 function is also required for development of terminal and skeletal structures, semilunar valvulogenesis in the heart, and hematopoiesis. Genes involved and proteins PTPN11 (Protein tyrosine phosphatase, non-receptor type, 11) Location 12q24.1 Centromere - FLJ34154 - RPL6 - PTPN11 - RPH3A OAS1 – telomere. DNA/RNA Description: The PTPN11 gene counts 16 exons. Exon 1 contains the 5'-UTR and the translation initiation ATG, and a few additional codons. Exon 15 contains the stop codon and exon 16 contains a major portion of the 3'-UTR. Other features of the PTPN11 gene, such as the promoter region and enhancer elements have not been delineated. Transcription: A 7.0-kb transcript is detected in several tissues (heart, brain, lung, liver, skeletal muscle, kidney, and pancreas) with highest steady-state levels in heart and skeletal muscle. The predominant human PTPN11 mRNA contains an open reading frame of 1,779 bases, resulting in a predicted protein of 593 amino acid residues. Pseudogene: A number of speudogenes, sharing more than 92% nucleotide identity with PTPN11 cDNA (including the untranslated regions), have been documented in the human genome. All the pseudogenes harbour frameshift mutations and multiple stop codons. Three of the five pseudogenes are likely to be expressed but with distinct tissue distribution and expression level. Protein Description: SHP-2 is a member of a small subfamily of cytoplasmic Src homology 2 (SH2) domaincontaining protein tyrosine phosphatases. The aminoterminal SH2 (N-SH2 and C-SH2) domains selectively bind to short amino acid motifs containing a phosphotyrosyl residue and promote SHP-2 association with cell surface receptors, cell adhesion molecules and scaffolding adapters. Crystallographic data indicate that the N-SH2 The PTPN11 gene and SHP-2 domain characterization. The coding exons are shown as numbered filled boxes. The functional domains of the protein, comprising two amino-terminal, tandemly arranged SH2 domains (N-SH2 and C-SH2) followed by a protein tyrosine phosphatase (PTP) domain, are shown below. Numbers below the domain structure indicate the amino-acid boundaries of those domains. Atlas Genet Cytogenet Oncol Haematol. 2005; 9(4) 347 Noonan syndrome Tartaglia M, Gelb BD Figure 3: Three-dimensional structure of SHP-2 in its catalytically inactive conformation, as determined by Hof and co-workers. Residues involved in catalysis are shown (space fill). Figure 4: Location of SHP-2 mutated residues in human disease. (A) Noonan syndrome and LEOPARD syndrome (germ-line origin; N=224); (B) Noonan syndrome with juvenile myelomonocytic leukemia (germ-line origin; N=11); (C) hematologic malignancies, including juvenile myelomonocytic leukemia, acute myeloid leukemia, acute lymphoblastic leukemia, myelodysplastic syndromes and chronic myelomonocytic leukemia (somatic origin; N=97). The pictures show the Ca trace of SHP-2 in its catalytically inactive conformation. Affected residues are indicated with their side chains as black sticks. Atlas Genet Cytogenet Oncol Haematol. 2005; 9(4) 348 Noonan syndrome Tartaglia M, Gelb BD EGF. Cell culture and whole embryo studies reveal that increased RAS/MAPK signaling is variably present, appearing to be cell-context specific. Somatic: Somatic activating mutations in PTPN11 have been documented in a heterogeneous group of hematologic malignancies and pre-leukemic disorders, and rarely in certain solid tumors. Selection: 181G>T (D61Y), 182A>T (D61V), 205G>A (E69K), 214G>A (A72T), 215C>T (A72V), 226G>A (E76K), 226G>C (E76Q), 227A>T (E76V), 227A>G (E76G), 227A>C (E76A), 1471C>T (P491S), 1472C>T (P491L), 1504T>C (S502P), 1504T>G (S502A), 1520C>A (T507K), 1528C>A (Q510K). Homology: PTPN6 (protein tyrosine phosphatase, nonreceptor type, 6) previously known as SHP1 or SHP-1 (Src homology 2 domain-containing protein tyrosine phosphatase, 1). Mutations Studies reported PTPN11 mutation detection rates ranging between 33% to 60%. Differences in inclusion criteria stringency and recruiting strategies are likely to responsible for such variable mutation detection rate. It should also be emphasized that the mutation prevalence detected in any Noonan syndrome cohort is sensitive to its composition in terms of relative abundance of sporadic and familial cases, as PTPN11 mutations appear to be more prevalent among families transmitting the trait compared to sporadic cases. With those issues stipulated, the contribution of PTPN11 mutations to the etiology of Noonan syndrome appears to be approximately 50%. A statistically significant association with pulmonary valve stenosis and lower incidence of hypertrophic cardiomyopathy was found among the group with PTPN11 mutations. Overall, a large percentage of PTPN11 mutation-negative individuals tended to exhibit fewer or mild clinical features of NS, even though approximately half of the Noonan syndrome patients without a PTPN11 mutation appeared clinically indistinguishable from typical PTPN11 mutation-positive patients. Germinal: The vast majority of mutations affect residues residing at or close to the interface between the N-SH2 and PTP domains. Increasing evidence supports that a number of Noonan syndrome-causative mutations promote SHP-2 gain-of-function by destabilizing the catalytically inactive conformation of the protein, and prolong signal flux through the RAS/MAPK pathway in a ligand-dependent manner. Selection: 124A>G (T42A), 182A>G (D61G), 184T>G (Y62D), 188A>G (Y63C), 214G>T (A72S), 215C>G (A72G), 218C>T (T73I), 228G>T, C (E76D), 236A>G (N79R), 317A>C (D106A), 922A>G (N308D). Two specific missense mutations (836A>G, Y279C; 1403C>T, T468M) have been identified to recur in LEOPARD syndrome, a developmental disorder closely related to Noonan syndrome. A mouse model bearing the NS-causative Asp61Gly mutation in the Ptpn11 gene has been recently generated and characterized. The Ptpn11D61G/ D61G genotype is embryonic lethal. At day E13.5, these embryos are grossly edematous and hemorrhagic, and have diffuse liver necrosis. A number of severe cardiac defects are also observed. Heterozygous embryos exhibit cardiac defects, proportionate growth failure and perturbed craniofacial development. Hematologic anomalies include a mild myeloproliferative disease. Ptpn11D61G/+ embryonic fibroblasts are characterized by a three-fold increased Shp-2 activity and increased association of Shp-2 with Gab1 after stimulation with Atlas Genet Cytogenet Oncol Haematol. 2005; 9(4) References Noonan JA. Hypertelorism with Turner phenotype. A new syndrome with associated congenital heart disease. Am J Dis Child. 1968 Oct;116(4):373-80 Nora JJ, Nora AH, Sinha AK, Spangler RD, Lubs HA. The Ullrich-Noonan syndrome (Turner phenotype). Am J Dis Child. 1974 Jan;127(1):48-55 Allanson JE. Noonan syndrome. J Med Genet. 1987 Jan;24(1):9-13 Burch M, Sharland M, Shinebourne E, Smith G, Patton M, McKenna W. Cardiologic abnormalities in Noonan syndrome: phenotypic diagnosis and echocardiographic assessment of 118 patients. J Am Coll Cardiol. 1993 Oct;22(4):1189-92 Jamieson CR, van der Burgt I, Brady AF, van Reen M, Elsawi MM, Hol F, Jeffery S, Patton MA, Mariman E. Mapping a gene for Noonan syndrome to the long arm of chromosome 12. Nat Genet. 1994 Dec;8(4):357-60 Saxton TM, Henkemeyer M, Gasca S, Shen R, Rossi DJ, Shalaby F, Feng GS, Pawson T. Abnormal mesoderm patterning in mouse embryos mutant for the SH2 tyrosine phosphatase Shp-2. EMBO J. 1997 May 1;16(9):2352-64 Hof P, Pluskey S, Dhe-Paganon S, Eck MJ, Shoelson SE. Crystal structure of the tyrosine phosphatase SHP-2. Cell. 1998 Feb 20;92(4):441-50 Legius E, Schollen E, Matthijs G, Fryns JP. Fine mapping of Noonan/cardio-facio cutaneous syndrome in a large family. Eur J Hum Genet. 1998 Jan;6(1):32-7 Qu CK, Yu WM, Azzarelli B, Cooper S, Broxmeyer HE, Feng GS. Biased suppression of hematopoiesis and multiple developmental defects in chimeric mice containing Shp-2 mutant cells. Mol Cell Biol. 1998 Oct;18(10):6075-82 Marino B, Digilio MC, Toscano A, Giannotti A, Dallapiccola B. Congenital heart diseases in children with Noonan syndrome: An expanded cardiac spectrum with high prevalence of atrioventricular canal. J Pediatr. 1999 Dec;135(6):703-6 Chen B, Bronson RT, Klaman LD, Hampton TG, Wang JF, Green PJ, Magnuson T, Douglas PS, Morgan JP, Neel BG. Mice mutant for Egfr and Shp2 have defective cardiac semilunar valvulogenesis. Nat Genet. 2000 Mar;24(3):296-9 Saxton TM, Ciruna BG, Holmyard D, Kulkarni S, Harpal K, Rossant J, Pawson T. The SH2 tyrosine phosphatase shp2 is required for mammalian limb development. Nat Genet. 2000 Apr;24(4):420-3 van Der Burgt I, Brunner H. Genetic heterogeneity in Noonan syndrome: evidence for an autosomal recessive form. Am J Med Genet. 2000 Sep 4;94(1):46-51 349 Noonan syndrome Tartaglia M, Gelb BD Tartaglia M, Mehler EL, Goldberg R, Zampino G, Brunner HG, Kremer H, van der Burgt I, Crosby AH, Ion A, Jeffery S, Kalidas K, Patton MA, Kucherlapati RS, Gelb BD. Mutations in PTPN11, encoding the protein tyrosine phosphatase SHP-2, cause Noonan syndrome. Nat Genet. 2001 Dec;29(4):465-8 Loh ML, Vattikuti S, Schubbert S, Reynolds MG, Carlson E, Lieuw KH, Cheng JW, Lee CM, Stokoe D, Bonifas JM, Curtiss NP, Gotlib J, Meshinchi S, Le Beau MM, Emanuel PD, Shannon KM. Mutations in PTPN11 implicate the SHP-2 phosphatase in leukemogenesis. Blood. 2004 Mar 15;103(6):2325-31 Digilio MC, Conti E, Sarkozy A, Mingarelli R, Dottorini T, Marino B, Pizzuti A, Dallapiccola B. Grouping of multiplelentigines/LEOPARD and Noonan syndromes on the PTPN11 gene. Am J Hum Genet. 2002 Aug;71(2):389-94 Tartaglia M, Cordeddu V, Chang H, Shaw A, Kalidas K, Crosby A, Patton MA, Sorcini M, van der Burgt I, Jeffery S, Gelb BD. Paternal germline origin and sex-ratio distortion in transmission of PTPN11 mutations in Noonan syndrome. Am J Hum Genet. 2004 Sep;75(3):492-7 Legius E, Schrander-Stumpel C, Schollen E, PullesHeintzberger C, Gewillig M, Fryns JP. PTPN11 mutations in LEOPARD syndrome. J Med Genet. 2002 Aug;39(8):571-4 Tartaglia M, Martinelli S, Cazzaniga G, Cordeddu V, Iavarone I, Spinelli M, Palmi C, Carta C, Pession A, Aricò M, Masera G, Basso G, Sorcini M, Gelb BD, Biondi A. Genetic evidence for lineage-related and differentiation stage-related contribution of somatic PTPN11 mutations to leukemogenesis in childhood acute leukemia. Blood. 2004 Jul 15;104(2):307-13 Tartaglia M, Kalidas K, Shaw A, Song X, Musat DL, van der Burgt I, Brunner HG, Bertola DR, Crosby A, Ion A, Kucherlapati RS, Jeffery S, Patton MA, Gelb BD. PTPN11 mutations in Noonan syndrome: molecular spectrum, genotype-phenotype correlation, and phenotypic heterogeneity. Am J Hum Genet. 2002 Jun;70(6):1555-63 Tartaglia M, Niemeyer CM, Shannon KM, Loh ML. SHP-2 and myeloid malignancies. Curr Opin Hematol. 2004 Jan;11(1):4450 Neel BG, Gu H, Pao L. The 'Shp'ing news: SH2 domaincontaining tyrosine phosphatases in cell signaling. Trends Biochem Sci. 2003 Jun;28(6):284-93 Zenker M, Buheitel G, Rauch R, Koenig R, Bosse K, Kress W, Tietze HU, Doerr HG, Hofbeck M, Singer H, Reis A, Rauch A. Genotype-phenotype correlations in Noonan syndrome. J Pediatr. 2004 Mar;144(3):368-74 Tartaglia M, Niemeyer CM, Fragale A, Song X, Buechner J, Jung A, Hählen K, Hasle H, Licht JD, Gelb BD. Somatic mutations in PTPN11 in juvenile myelomonocytic leukemia, myelodysplastic syndromes and acute myeloid leukemia. Nat Genet. 2003 Jun;34(2):148-50 Jongmans M, Sistermans EA, Rikken A, Nillesen WM, Tamminga R, Patton M, Maier EM, Tartaglia M, Noordam K, van der Burgt I. Genotypic and phenotypic characterization of Noonan syndrome: new data and review of the literature. Am J Med Genet A. 2005 Apr 15;134A(2):165-70 Araki T, Mohi MG, Ismat FA, Bronson RT, Williams IR, Kutok JL, Yang W, Pao LI, Gilliland DG, Epstein JA, Neel BG. Mouse model of Noonan syndrome reveals cell type- and gene dosage-dependent effects of Ptpn11 mutation. Nat Med. 2004 Aug;10(8):849-57 Lee JS, Tartaglia M, Gelb BD, Fridrich K, Sachs S, Stratakis CA, Muenke M, Robey PG, Collins MT, Slavotinek A. Phenotypic and genotypic characterisation of Noonanlike/multiple giant cell lesion syndrome. J Med Genet. 2005 Feb;42(2):e11 Bentires-Alj M, Paez JG, David FS, Keilhack H, Halmos B, Naoki K, Maris JM, Richardson A, Bardelli A, Sugarbaker DJ, Richards WG, Du J, Girard L, Minna JD, Loh ML, Fisher DE, Velculescu VE, Vogelstein B, Meyerson M, Sellers WR, Neel BG. Activating mutations of the noonan syndrome-associated SHP2/PTPN11 gene in human solid tumors and adult acute myelogenous leukemia. Cancer Res. 2004 Dec 15;64(24):8816-20 Tartaglia M, Gelb BD. Germ-line and somatic PTPN11 mutations in human disease. Eur J Med Genet. 2005 AprJun;48(2):81-96 Tartaglia M, Gelb BD. Noonan syndrome and related disorders: genetics and pathogenesis. Annu Rev Genomics Hum Genet. 2005;6:45-68 Fragale A, Tartaglia M, Wu J, Gelb BD. Noonan syndromeassociated SHP2/PTPN11 mutants cause EGF-dependent prolonged GAB1 binding and sustained ERK2/MAPK1 activation. Hum Mutat. 2004 Mar;23(3):267-77 Tartaglia M, Martinelli S, Iavarone I, Cazzaniga G, Spinelli M, Giarin E, Petrangeli V, Carta C, Masetti R, Aricò M, Locatelli F, Basso G, Sorcini M, Pession A, Biondi A. Somatic PTPN11 mutations in childhood acute myeloid leukaemia. Br J Haematol. 2005 May;129(3):333-9 Loh ML, Reynolds MG, Vattikuti S, Gerbing RB, Alonzo TA, Carlson E, Cheng JW, Lee CM, Lange BJ, Meshinchi S. PTPN11 mutations in pediatric patients with acute myeloid leukemia: results from the Children's Cancer Group. Leukemia. 2004 Nov;18(11):1831-4 Atlas Genet Cytogenet Oncol Haematol. 2005; 9(4) This article should be referenced as such: Tartaglia M, Gelb BD. Noonan syndrome. Atlas Genet Cytogenet Oncol Haematol. 2005; 9(4):345-350. 350