Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
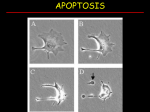
Toad Lilies Elephant Frog Cell- cell interaction • Cells may communicate by direct contact. Immune connection: Macrophage will use direct contact to alert T cells that invaders are present Copyright © 2002 Pearson Education, Inc., publishing as Benjamin Cummings Fig. 11.4 Epithelial Cell Junction Types Structures in the B7/CD28 family. Structures are modeled on the crystal determinations. Loops have been added to one end of the IgV domains to emphasize the orientation of the CDR-like loops and their interaction with ligand or lack thereof. General Schemes of Intercellular Signalling Extracellular signaling molecules released by cells occurs over distances from a few microns - autocrine (c) and paracrine (b) signaling to several meters in endocrine (a) signaling. In some instances, receptor proteins attached to the membrane of one cell interact directly with receptors on an adjacent cell (d). © 2000 by W. H. Freeman and Company. All rights reserved. Representative (typical) Neuron http://www.horton.ednet.n s.ca/staff/selig/Activities/n ervous/na1.htm 1. cell body or soma -single nucleus with prominent nucleolus -Nissl bodies -rough ER & free ribosomes for protein synthesis -proteins then replace neuronal cellular components for growth and repair of damaged axons in the PNS -neurofilaments or neurofibrils give cell shape and support bundles of intermediate filaments -microtubules move material inside cell -lipofuscin pigment clumps (harmless aging) - yellowish brown Anatomy of a Synapse Copyright © 2008 Pearson Education, Inc., publishing as Benjamin Cummings. Figure 8.2a Propagation of an Action Potential Why is the resting potential a negative number? What is the significance of the refractory period? Refractory Period Neurotransmitters • • • • • More than 100 identified produced by neurons and stored within the neuron secrete these NTs in response to generation of an electrical signal (action potential) by the neuron bind onto target neurons (synapse) or target muscle cells (neuromuscular junction) Some bind receptors on the target and cause channels to open in the target (sodium channels) – – • • e.g. binds to receptors on target neuron – causes generation of another action potential by target neuron e.g. binds to receptors on target muscle cells – causes contraction Others bind receptors on the target and result in a second messenger system Results in either excitation or inhibition of the target Neuromuscular junction Neuromuscular junction EPSPs: excitatory post-synaptic potentials IPSPs: inhibitory post-synaptic potentials What kinds of hormone are there? Known Hormonal Classes • Proteins & peptides chemcases.com/olestra/ images/insulin.jpg • Lipids (steroids, eicosanoids) • Amino acid derived (thyronines, neurotransmitters) chem.pdx.edu/~wamserc/ ChemWorkshops/ gifs/W25_1.gif • Gases (NO, CO) website.lineone.net/~dave.cushman/ epinephrine.gif What is a hormone receptor? Hormone Receptors are cellular proteins that bind with high affinity to hormones & are altered in shape & function by binding; they exist in limited numbers. Binding to hormone is noncovalent & reversible. Hormone binding will alter binding to other cellular proteins & may activate any receptor protein enzyme actions. What are the main types of receptors? Membrane Receptors Imbedded in target cell membrane; integral proteins/ glycoproteins; penetrate through membrane For protein & charged hormones (peptides or neurotransmitters) 3 major groups: Serpentine = 7 transmembrane domains, Growth factor/cytokine = 1 transmembrane domain, Ion channels • • • • • easily travels through the blood - hydrophilic but cannot diffuse through plasma membrane! therefore absolutely requires the expression of receptors on the cell surface – integral membrane proteins that act as first messenger the receptor protein activates a series of signaling events within the cells – e.g. epinephrine binds to receptor and activates an adjacent G-protein in membrane – G-protein activates adenylate cyclase to convert ATP to cyclic AMP (cAMP) in the cytosol – cAMP acts as a 2nd messenger – cAMP activates a series of proteins in the cytosol called kinases – kinases act to phosphorylate their targets – either activating them or inhibiting them – this speeds up/slows down physiological responses within the cell – phosphodiesterase inactivates cAMP quickly many second messengers are made in cells in response to specific hormones – • • e.g. calcium, IP3, DAG Cell response is turned off unless new hormone molecules arrive this mechanism allows for amplification – one H-R combination can activate two G proteins which activates 4 kinases which activate 16 more kinases etc……. What are the main types of receptors? Nuclear Receptors Nuclear proteins that usually act in pairs & bind to specific Hormone Recognition Elements (HREs) = sequences on the DNA in the promoter regions of target genes For small, hydrophobic molecules (steroids, thyroid hormones) Action of Lipid-Soluble Hormones: Endogenous signaling • • • • • • • • • Hormone must be carried by a transport protein that allows it to dissolve within the aqueous (watery) environment of the blood plasma Hormone diffuses through phospholipid bilayer & into cell the receptor is located within the cell (cytoplasm or the nucleus) binding of H to R results in its translocation into the nucleus the H then binds directly to specific sequences within the DNA = response elements this binding turns on/off specific genes – activates or inhibits gene transcription if turned on - new mRNA is formed & directs synthesis of new proteins new protein alters cell’s activity if turned off – no new protein results and the cell’s activity is altered Signal transduction: Adenyl cyclase system Gs/i/o/x – G proteins, CaM – calmodulin, GTP – guanosine triphosphate, ATP – adenosine triphosphate, ADP – adenosine diphosphate, cAMP – cyclic adenosine monophosphate, 5´-AMP - 5´-adenosine monophosphate, PKA – protein kinase A, PKC – protein kinase C Signal transduction: Phosphoinositide system PI-PLC - phospholipase C specific for phosphoinositides, PIP2 - phosphatidylinositol4,5-biphosphate, IP3 - inositol-1,4,5-triphosphate; DG - diacylglycerol; PKC – protein kinase C AR – adrenoceptor, G – G protein, PI-PLC – phosphoinositide specific phospholipase C, IP3 – inositoltriphosphate, DG – diacylglycerol, CaM – calmodulin, AC – adenylyl cyclase, PKC – protein kinase C Gs/i/o/x – G proteins, CaM – calmodulin, GTP – guanosine triphosphate, ATP – adenosine triphosphate, ADP – adenosine diphosphate, cAMP – cyclic adenosine monophosphate, 5´-AMP - 5´-adenosine monophosphate, PKA – protein kinase A, PKC – protein kinase C • Models of estrogen action. In the “classical” pathway of estrogen action (i), estrogen or other selective estrogen receptor modulators (SERMs) bind to the estrogen receptor (ER), a ligand-activated transcription factor that regulates transcription of target genes in the nucleus by binding to estrogen response element (ERE) regulatory sequences in target genes and recruiting coregulatory proteins (CoRegs) such as coactivators. Rapid or “nongenomic” effects of estrogen may also occur through the ER located in or adjacent to the plasma membrane (ii), which may require the presence of “adaptor” proteins, which target the ER to the membrane. Activation of the membrane ER leads to a rapid change in cellular signaling molecules and stimulation of kinase activity, which in turn may affect transcription. Lastly, other non-ER membrane-associated estrogenbinding proteins (EBPs) may also trigger an intracellular response (iii). • Risk factors can be distinguished in terms of their ability to cause breast cancer directly, through genetic damage, or by altering hormonal metabolism. Vulnerability factors prolong the duration of breast cell growth, while contributing factors can distort hormone levels. • Bifunctional pathways to breast cancer. Abbreviations: E2, 17ß-estradiol; E1, estrone; OHE1, hydroxyestrone; ER, estrogen receptor. In the bifunctional pathway, the E2 metabolites affect cell proliferation and breast cancer development either directly via receptorindependent mechanisms involving structural/functional alterations in DNA, or indirectly via receptordependent mechanisms involving phenotypic growth regulation. Both mechanisms eventually upregulate aberrant proliferation and development of breast cancer (37). • The response of a particular cell to a signal depends on its particular collection of receptor proteins, relay proteins, and proteins needed to carry out the response. Figure 19-34 Fluctuating Levels of Mitotic Cyclin and MPF During the Cell Cycle Figure 19-38 Role of the Rb Protein in Cell Cycle Control Role of the p53 Protein in Responding to DNA Damage Apoptosis • Apoptosis (1972) – Greek word “falling off” • Built-in (programmed) mechanism) • or self-destructionsuicide • Type of programmed cell death based upon morphological features Programmed cell death during development. Programmed cell death is involved in forming structures such as the digits of the hand (a), deleting structures such as nearly all of an insect's larval components (b), controlling cell numbers in, for example, the nervous system (c) and eliminating abnormal cells such as those that harbour mutations (d). Studies on the development of the nervous system showed that in the process of assembling sensory fields, neurons are eliminated by orderly cell death in order to tailor sensory input to environmental stimuli (elimination or transplantation of limbs as key examples). Apoptosis plays in an important role in normal developmental processes Jacobson et al (1997) Cell, Vol. 88, 347– A cancer cell (mauve) undergoing apoptosis Comparison of cell death by necrosis and apoptosis Known Apoptotic Stimuli • • • • • • • Withdrawal of NGF Etoposide Actinomycin D UV radiation Staurosporin Enforced m-Myc expression Glucocorticoids Physiological Relevance of Apoptosis • • • • • • • • • • Embryonic Development Regulation of/by Immune System Negative Selection CTL Killing (eg. Immune surveillance, viral infections) Terminating Active Immune Response Tight Regulation of Cell Number (eg. BM, GI, Uterus, Skin) Compensatory Response to Cell Stress Intrinsic Pathway (e.g. GF removal, XRT, Chemo.) Extrinsic Pathway (e.g. FAS.Ag, TNF-R Activation) Senescence (ageing) Morphology of Apoptosis – – – – – – cell shrinkage extracellular exposure of phosphatidylserine shows boiling and blebbing chromatin condenses*-most characteristic feature DNA is degraded into oligonucleosomal fragments disassembly into apoptotic bodies • membrane bound, contains portions of the nucleus and various organelles – phagocytosis by neighboring cells Does not elicit inflammation-hallmark of apoptosis MORPHOLOGICAL FEATURES OF APOPTOSIS • Cell shrinkage • Chromatin condensation and fragmentation. • Formation of cytoplasmic blebs and apoptotic bodies. • Phagocytosis of apoptotic bodies by adjacent healthy cells or macrophages. • Lack of inflammation. Morphological and biochemical characteristics of apoptosis Morphologi changes: Early : Chromosome condensation, cell body shrink Later : Blebbing and Nucleus and cytoplasm fragment— Apoptotic bodies At last: Phagocytosed Biochemical features of apoptosis 1.PROTEIN CLEAVAGE: Caspases (cysteine protease) Nuclear scaffold Cytoskeletal protein 2.PROTEIN CROSS-LINKING: Transglutaminase Cytoplasmic proteinshrunken shalls apoptotic bodies Biochemical features of apoptosis 3. DNA breakdown: 50-300 kb pieces Ca2+, Mg2+ dependent endonucleases DNA oligonucleosomes DNA ladders (also seen in necrosis) 4. PHAGOCYTIC RECOGNITION Receptors on macrophages for the surface components (phosphatidylserine, thrombospondin) on apoptotic bodies. DNA fragmentation: biochemical hallmark • DNA cleaved into non-random fragments • Targets of endonuclease attack: linker regions between nucleosomes • 180-200 bp fragments & multiples of this unit Agarose gel electrophoresis Chromatin Fragmented Chromatin mitochondria Caspase-3 activation via tumor necrosis factor (TNF) family receptors (for example, Fas), FADD (Fas-activated death domain protein) and caspase-8 represents the extrinsic pathway (blue), whereas caspase-3 activation via the mitochondrial release of cytochrome c and Apaf-1–mediated processing of caspase9 represents the intrinsic pathway (red)3. For clarity, not all of the players are shown. Procaspase-3 is shown as a PAC-1– sensitive dormant single-chain precursor with an N-terminal prodomain (Pro). During apoptosis, caspase-3 assembles as an active p17-p12 heterotetramer after proteolytic processing between the p17 and p12 subunits (at Asp175) and removal of the prodomain2. PAC-1 is proposed to regulate the Asp-Asp-Asp (DDD) safety catch at amino acids 179–181 in procaspase-3, consequently inducing a conformational change that leads to proteolytic processing into the active p17 and p12 subunits1. Cys163 is the catalytic cysteine in the active site of caspase-3; the sequence shown illustrates its proximity to the DDD safety catch and DDM motif. Although caspase-7 (not shown) is believed to be a downstream caspase, its position relative to caspase-3 in apoptosis pathways is unclear. Functional homologues of caspases and caspase regulators across species are indicated by the same colour. Caspase-9 in mammals and Dronc in the fruitfly Drosophila melanogaster are initiator caspases, whereas caspase-3 and -7 in mammals and Drice in fruitflies belong to the class of effector caspases. CED-3 (cell-death abnormality-3) in the nematode worm Caenorhabditis elegans functions both as an initiator and effector caspase. The inhibitor of apoptosis (IAP) proteins suppress apoptosis by negatively regulating the caspases, whereas SMAC (second mitochondria-derived activator of caspases)/DIABLO (direct IAP-binding protein with low pI) in mammals and the RHG proteins Reaper, Hid, Grim and Sickle in fruitflies can remove the IAP-mediated negative regulation of caspases. AIF, apoptosis-inducing factor; APAF1, apoptotic-protease-activating factor-1; Cyt c, cytochrome c; EndoG, endonuclease G; HTRA2, high-temperature-requirement protein A2. Role of Caspases • Effectors (cell disassembly) (caspases 2,3,6,7) and initiators (caspases 8,9) • 14 identified mammalian caspases- 12 in humans • Cysteine protease that has an absolute requirement requirement for cleavage after aspartic acid • High specificity for which proteins are digested – PARP (116 kDa) nuclear polymerase that repairs DNA is cleaved by caspase 7. Types of Caspases • • • • • • • • • • Nedd2/Ich-1/caspase2 YAMA/CPP32/apopain/caspase-3- most characterized TX/Ich-2/ICErelII/caspase-4 TY/ICErelIII/caspase-5 Mch2/caspase-6 ICE-LAP-3/Mch-3/CMH1/caspase-7 FLICE/MACH/caspase-8-most proximally activated caspase ICE-LAP-6/caspase-9 Mch-4/FLICE 2/caspase-10 Ich3/caspase-11 Caspases (cysteine aspartic acid-specific proteases) are highly specific proteases that cleave their substrates after specific tetrapeptide motifs (P4-P3-P2-P1) where P1 is an Asp residue. The caspase family can be subdivided into initiators, which are able to auto-activate and initiate the proteolytic processing of other caspases, and effectors, which are activated by other caspase molecules. The effector caspases cleave the vast majority of substrates during apoptosis. All caspases have a similar domain structure comprising a propeptide followed by a large and a small subunit (see figure). The pro-peptide can be of variable length and, in the case of initiator caspases, can be used to recruit the enzyme to activation scaffolds such as the APAF1 apoptosome. Two distinct, but structurally related, pro-peptides have been identified; the caspase recruitment domain (CARD) and the death effector domain (DED), and these domains typically facilitate interaction with proteins that contain the same motifs. Caspase activation is usually initiated through proteolytic processing of the caspase between the large and small subunits to form a heterodimer. This processing event rearranges the caspase active site into the active conformation. Caspases typically function as heterotetramers, which are formed through dimerization of two caspase heterodimers. Initiator caspases exist as monomers in healthy cells, whereas effector caspases are present as pre-formed dimers. Not all mammalian caspases participate in apoptosis. For example, caspase-1, caspase-4, caspase-5 and caspase-12 are activated during innate immune responses and are involved in the regulation of inflammatory cytokine processing (for example, IL1 and IL18). Interestingly, caspase-12 is expressed as a truncated, catalytically inactive protein in most humans (caspase-12S*). However, a subset of individuals of African descent express full-length caspase-12 (caspase-12L*) and these individuals appear to be more susceptible to inflammatory diseases. To date, 400 substrates for the mammalian caspases have been identified, but the significance of many of these cleavage events remains obscure. Caspase 3 • Caspase 3 (CPP32/apopain/YAMA) – shares similarity to CED 3. – Protein substrates include: • PARP (poly ADP ribose) polymerase • PKC • sterol-regulatory element-binding protein • DNA dependent protein kinase (DNA repair) • U1-associated 70 kDa protein (mRNA splicing) • MEKK • DNA fragmentation factor- cytosolic factor that induces nuclear fragmentation • DNA damage-apoptosis model structure. The diagram depicts the general structure of the model. The model contains mathematical equations describing the individual protein-protein interactions and catalytic reactions for over 80 species that fall within the broad outlines shown. The equations are solved in specialised software including MatLab, and are run multiple times to simulate multi-cell environments. p53 Dependent Apoptotic Pathways Schematic representation of the p53-dependent apoptotic pathways by transcriptional activation of BAX, PUMA and APAF-1. Fertilization in sea urchin. 1. The acrosome releases enzymes to digest the jelly coat. 2. Actin filaments bind to receptors in the vitelline layer. 3. The sperm and egg plasma membranes fuse and become depolarized, preventing polyspermy the ability of other sperm to fertilize the egg. 4. The sperm nucleus enters and fuses with the egg's nucleus. 5. The vitelline layer swells to form a fertilization envelope that also blocks polyspermy. Releases ZPdegrading enzymes (from Gilbert, Developmental Biology) The Formation of Primary Germ Layers Figure Stages of prenatal development Frill-necked Lizard Squid lizard