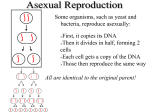
Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
Cancer epigenetics wikipedia , lookup
Point mutation wikipedia , lookup
Site-specific recombinase technology wikipedia , lookup
Vectors in gene therapy wikipedia , lookup
Genome (book) wikipedia , lookup
Polycomb Group Proteins and Cancer wikipedia , lookup
Mir-92 microRNA precursor family wikipedia , lookup
PowerPoint to accompany Genetics: From Genes to Genomes Fourth Edition Leland H. Hartwell, Leroy Hood, Michael L. Goldberg, Ann E. Reynolds, and Lee M. Silver Prepared by Mary A. Bedell University of Georgia Copyright © The McGraw-Hill Companies, Inc. Permission required to reproduce or display Hartwell et al., 4th edition 1 PART V How Genes Are Regulated CHAPTER Somatic Mutation and the Genetics of Cancer CHAPTER OUTLINE 17.1 Overview: Initiation of Division 17.2 Cancer: A Failure of Control over Cell Division 17.3 The Normal Control of Cell Division Copyright © The McGraw-Hill Companies, Inc. Permission required to reproduce or display Hartwell et al., 4th edition, Chapter 17 2 The relative percentages of new cancers in the United States that occur at different sites Fig. 17.1 Copyright © The McGraw-Hill Companies, Inc. Permission required to reproduce or display Hartwell et al., 4th edition, Chapter 17 3 Two unifying themes about cancer genetics Cancer is a disease of genes • Multiple cancer phenotypes arise from mutations in genes that regulate cell growth and division • Environmental chemicals increase mutation rates and increase chances of cancer Cancer has a different inheritance pattern than other genetic disorders • Inherited mutations can predispose to cancer, but the mutations causing cancer occur in somatic cells • Mutations accumulate in clonal descendants of a single cell Copyright © The McGraw-Hill Companies, Inc. Permission required to reproduce or display Hartwell et al., 4th edition, Chapter 17 4 Overview of the initiation of cell division Two basic types of signals that tell cells whether to divide, metabolize, or die Extracellular signals – act over long or short distances • Collectively known as hormones • Steroids, peptides, or proteins Cell-bound signals – require direct contact between cells Copyright © The McGraw-Hill Companies, Inc. Permission required to reproduce or display Hartwell et al., 4th edition, Chapter 17 5 An example of an extracellular signal that acts over large distances Thyroid-stimulating hormone (TSH) produced in pituitary gland Moves through blood to thyroid gland, which expresses thyroxine Fig. 17.2a Copyright © The McGraw-Hill Companies, Inc. Permission required to reproduce or display Hartwell et al., 4th edition, Chapter 17 6 An example of an extracellular signal that is mediated by cell-to-cell contact Fig. 17.2b Copyright © The McGraw-Hill Companies, Inc. Permission required to reproduce or display Hartwell et al., 4th edition, Chapter 17 7 Each signaling system has four components Growth factors • Extracellular hormones or cell-bound signals that stimulate or inhibit cell proliferation Receptors • Comprised of a signal-binding site outside the cell, a transmembrane segment, and an intracellular domain Signal transducers • Located in cytoplasm Transcription factors • Activate expression of specific genes to either promote or inhibit cell proliferation Copyright © The McGraw-Hill Companies, Inc. Permission required to reproduce or display Hartwell et al., 4th edition, Chapter 17 8 Hormones transmit signals into cells through receptors that span the cellular membrane Fig. 17.3a Copyright © The McGraw-Hill Companies, Inc. Permission required to reproduce or display Hartwell et al., 4th edition, Chapter 17 9 Signaling systems can stimulate or inhibit growth Signal transduction activation or inhibition of intracellular targets after binding of growth factor to its receptor Fig. 17.3b&c Copyright © The McGraw-Hill Companies, Inc. Permission required to reproduce or display Hartwell et al., 4th edition, Chapter 17 10 RAS is an intracellular signaling molecule Fig. 17.3d Copyright © The McGraw-Hill Companies, Inc. Permission required to reproduce or display Hartwell et al., 4th edition, Chapter 17 11 Cancer phenotypes result from the accumulation of mutations Mutations are in genes controlling proliferation as well as other processes • Result in a clone of cells that overgrows normal cells Cancer phenotypes include: • Uncontrolled cell growth • Genomic and karyotypic instability • Potential for immortality • Ability to invade and disrupt local and distant tissues Copyright © The McGraw-Hill Companies, Inc. Permission required to reproduce or display Hartwell et al., 4th edition, Chapter 17 12 Phenotypic changes that produce uncontrolled cell growth Most normal cells Autocrine stimulation: Cancer cells can make their own stimulatory signals a.1 Most normal cells Loss of contact inhibition: Many cancer cells Many cancer cells a.2 Growth of cancer cells doesn't stop when the cells contact each other Fig. 17.4 Copyright © The McGraw-Hill Companies, Inc. Permission required to reproduce or display Hartwell et al., 4th edition, Chapter 17 13 Phenotypic changes that produce uncontrolled cell growth (cont) Most normal cells Loss of cell death: Cancer cells are more resistant to programmed cell death (apoptosis) Loss of gap junctions: Many cancer cells a.3 Most normal cells Many cancer cells a.4 Cancer cells lose channels for communicating with adjacent cells Fig. 17.4 Copyright © The McGraw-Hill Companies, Inc. Permission required to reproduce or display Hartwell et al., 4th edition, Chapter 17 14 Phenotypic changes that produce genomic and karyotypic instability Defects in DNA replication machinery: b.1 Cancer cells have lost the ability to replicate their DNA accurately Increased mutation rates can occur because of defects in DNA replication machinery Fig. 17.4 Copyright © The McGraw-Hill Companies, Inc. Permission required to reproduce or display Hartwell et al., 4th edition, Chapter 17 15 Phenotypic changes that produce genomic and karyotypic instability (cont) Increased rate of chromosomal aberrations: Cancer cells often have chromosome rearrangements (translocations, deletions, aneuploidy, etc) Some rearrangements appear regularly in specific tumor types Fig. 17.4b.2 Fig. 17.4b.2 Copyright © The McGraw-Hill Companies, Inc. Permission required to reproduce or display Hartwell et al., 4th edition, Chapter 17 16 Phenotypic changes that produce a potential for immortality Loss of limitations on the number of cell divisions: Tumor cells can divide indefinitely in culture (below) and express telomerase (not shown) Most normal cells Many cancer cells c.1 Immortality c.2 Growth in soft agar Fig. 17.4 Copyright © The McGraw-Hill Companies, Inc. Permission required to reproduce or display Hartwell et al., 4th edition, Chapter 17 17 Phenotypic changes that enable a tumor to disrupt local tissue and invade distant tissues Ability to metastasize: Tumor cells can invade the surrounding tissue and travel through the bloodstream d.1 Angiogenesis: Tumor cells can secrete substances that promote growth of blood vessels d.2 Fig. 17.4 Copyright © The McGraw-Hill Companies, Inc. Permission required to reproduce or display Hartwell et al., 4th edition, Chapter 17 18 Evidence from mouse models that cancer is caused by several mutations Transgenic mice with dominant mutations in the myc gene and in the ras gene Mice with recessive mutations in the p53 gene (a) (b) Fig. 17.5a Fig. 17.5 Copyright © The McGraw-Hill Companies, Inc. Permission required to reproduce or display Hartwell et al., 4th edition, Chapter 17 19 Evidence that cancer cells are clonal descendants of a single somatic cell Analysis of polymorphic enzymes encoded by the X chromosome in females Sample from normal tissues has mixture of both alleles Clones of normal cells has only one allele Sample from tumor has only one allele Fig. 17.6 Copyright © The McGraw-Hill Companies, Inc. Permission required to reproduce or display Hartwell et al., 4th edition, Chapter 17 20 The incidence of some common cancers varies between countries Table 17.1 Copyright © The McGraw-Hill Companies, Inc. Permission required to reproduce or display Hartwell et al., 4th edition, Chapter 17 21 The role of environmental mutagens in cancer Concordance for the same type of cancer in first degree relatives (i.e. siblings) is low for most forms of cancer The incidence of some cancers varies between countries (see Table 17.2) • When a population migrates to a new location, the cancer profile becomes like that of the indigenous population Numerous environmental agents are mutagens and increase the likelihood of cancer • Some viruses, cigarette smoke Copyright © The McGraw-Hill Companies, Inc. Permission required to reproduce or display Hartwell et al., 4th edition, Chapter 17 22 Cancer development over time Lung cancer death rates and incidence of cancer with age Fig. 17.7 Copyright © The McGraw-Hill Companies, Inc. Permission required to reproduce or display Hartwell et al., 4th edition, Chapter 17 23 Some families have a genetic predisposition to certain types of cancer Example: retinoblastoma caused by mutations in RB gene Individuals who inherit one copy of the RB− allele are prone to cancer of the retina During proliferation of retinal cells, the RB+ allele is lost or mutated Tumors develop as a clone of RB−/RB− cells Fig. 17.8 Copyright © The McGraw-Hill Companies, Inc. Permission required to reproduce or display Hartwell et al., 4th edition, Chapter 17 24 Cancer is thought to arise by successive mutations in a clone of proliferating cells Fig. 17.9 Copyright © The McGraw-Hill Companies, Inc. Permission required to reproduce or display Hartwell et al., 4th edition, Chapter 17 25 Cancer-producing mutations are of two general types Fig. 17.10 Copyright © The McGraw-Hill Companies, Inc. Permission required to reproduce or display Hartwell et al., 4th edition, Chapter 17 26 Oncogenes act dominantly and cause increased proliferation Oncogenes are produced when mutations cause improper activation a gene Two approaches to identifying oncogenes: • Tumor-causing viruses (Fig 17.11a) Many tumor viruses in animals are retroviruses Some DNA viruses carry oncogenes [e.g. Human papillomavirus (HPV)] • Tumor DNA (Fig. 17.11b) Transform normal mouse cells in culture with human tumor DNA Copyright © The McGraw-Hill Companies, Inc. Permission required to reproduce or display Hartwell et al., 4th edition, Chapter 17 27 Cancer-causing retroviruses carry a mutant or overexpressed copy of a cellular gene After infection, retroviral genome integrates into host genome If the retrovirus integrates near a proto-oncogene, the protooncogene can be packaged with the viral genome Fig. 17.11a Copyright © The McGraw-Hill Companies, Inc. Permission required to reproduce or display Hartwell et al., 4th edition, Chapter 17 28 Retroviruses and their associated oncogenes Table 17.2 Copyright © The McGraw-Hill Companies, Inc. Permission required to reproduce or display Hartwell et al., 4th edition, Chapter 17 29 DNA from human tumor cells is able to transform normal mouse cells into tumor cells Human gene that is oncogenic can be identified and cloned from transformed mouse cells Fig. 17.11b Copyright © The McGraw-Hill Companies, Inc. Permission required to reproduce or display Hartwell et al., 4th edition, Chapter 17 30 The RAS oncogene is the mutant form of the RAS proto-oncogene Normal RAS is inactive until it becomes activated by binding of growth factors to their receptors Oncogenic forms of RAS are constitutively activated Fig. 17.11c Copyright © The McGraw-Hill Companies, Inc. Permission required to reproduce or display Hartwell et al., 4th edition, Chapter 17 31 Oncogenes are members of signal transduction systems Table 17.3 Copyright © The McGraw-Hill Companies, Inc. Permission required to reproduce or display Hartwell et al., 4th edition, Chapter 17 32 Cancer can be caused by mutations that improperly inactivate tumor suppressor genes Function of normal allele of tumor suppressor genes is to control cell proliferation Mutant tumor suppressor alleles act recessively and cause increased cell proliferation Tumor suppressor genes identified through genetic analysis of families with inherited predisposition to cancer • Inheritance of a mutant tumor suppressor allele • One normal allele sufficient for normal cell proliferation in heterozygotes • Wild-type allele in somatic cells of heterozygote can be lost or mutated abnormal cell proliferation Copyright © The McGraw-Hill Companies, Inc. Permission required to reproduce or display Hartwell et al., 4th edition, Chapter 17 33 The retinoblastoma tumor-suppressor gene Fig. 17.12 Copyright © The McGraw-Hill Companies, Inc. Permission required to reproduce or display Hartwell et al., 4th edition, Chapter 17 34 Mutant alleles of these tumor-suppressor genes decrease the accuracy of cell reproduction Table 17.4 Copyright © The McGraw-Hill Companies, Inc. Permission required to reproduce or display Hartwell et al., 4th edition, Chapter 17 35 The normal control of cell division Four phases of the cell cycle: G1, S, G2, and M Fig. 17.13 Copyright © The McGraw-Hill Companies, Inc. Permission required to reproduce or display Hartwell et al., 4th edition, Chapter 17 36 Experiments with yeast helped identify genes that control cell division Two kinds of used: Saccharomyces cerevisiae (budding yeast) and Schizosaccharomyces pombe (fission yeast) Usefulness of yeast for studies of the cell cycle • Both grow as haploids or diploids Can identify recessive mutations in haploids Can do complementation analysis in diploids • S. cerevisiae – size of buds serves as a marker of progress through the cell cycle Daughter cells arise as small buds on mother cell at end of G1 and grow during mitosis Stage of cell cycle can be determined by relative appearance of buds (see Fig 17.14) Copyright © The McGraw-Hill Companies, Inc. Permission required to reproduce or display Hartwell et al., 4th edition, Chapter 17 37 The isolation of temperature-sensitive mutants of yeast Mutants grow normally at permissive temperature (22°) At restrictive temperature (36°), mutants lose gene function After replica plating, colonies that grow at 22° but not at 36° have temperature-sensitive mutation Fig. 17.15 Copyright © The McGraw-Hill Companies, Inc. Permission required to reproduce or display Hartwell et al., 4th edition, Chapter 17 38 A temperature-sensitive cell-cycle mutant in S. cerevesiae Cells grown at permissive temperature display buds of all sizes (asynchronous division) Growth of the same cells at restrictive temperature – all have large buds Fig. 17.14a Copyright © The McGraw-Hill Companies, Inc. Permission required to reproduce or display Hartwell et al., 4th edition, Chapter 17 Fig. 17.14b 39 Some important cell-cycle and DNA repair genes Table 17.5 Copyright © The McGraw-Hill Companies, Inc. Permission required to reproduce or display Hartwell et al., 4th edition, Chapter 17 40 CDKs interact with cyclins and control the cell cycle by phosphorylating other proteins Cyclin-dependent kinases (CDKs) – family of kinases that regulate the transition from G1 to S and from G2 to M • Cyclin specifies the protein targets for CDK Phosphorylation by CDKs can activate or inactive a protein Fig. 17.16a Copyright © The McGraw-Hill Companies, Inc. Permission required to reproduce or display Hartwell et al., 4th edition, Chapter 17 41 CDKs control the dissolution of the nuclear membrane at mitosis Lamins – provide structural support to the nucleus • Form an insoluble matrix during most of the cell cycle At mitosis, lamins are phosphorylated by CDKs and become soluble Fig. 17.16 Copyright © The McGraw-Hill Companies, Inc. Permission required to reproduce or display Hartwell et al., 4th edition, Chapter 17 42 Mutant yeast permit the cloning of a human CDK gene Human CDKs and cyclins can function in yeast and replace the corresponding yeast proteins Fig. 17.17 Copyright © The McGraw-Hill Companies, Inc. Permission required to reproduce or display Hartwell et al., 4th edition, Chapter 17 43 CDKs mediate the transition from the G1 to the S phase of the cell cycle Fig. 17.18 Copyright © The McGraw-Hill Companies, Inc. Permission required to reproduce or display Hartwell et al., 4th edition, Chapter 17 44 CDK activity in yeast is controlled by phosphorylation and dephosphorylation Fig. 17.19 Copyright © The McGraw-Hill Companies, Inc. Permission required to reproduce or display Hartwell et al., 4th edition, Chapter 17 45 Cell-cycle checkpoints ensure genomic stability Checkpoints monitor the genome and cell-cycle machinery before allowing progression to the next stage of cell cycle G1-to-S checkpoint • DNA synthesis can be delayed to allow time for repair of DNA that was damaged during G1 The G2-to-M checkpoint • Mitosis can be delayed to allow time for repair of DNA that was damaged during G2 Spindle checkpoint • Monitors formation of mitotic spindle and engagement of all pairs of sister chromatids Copyright © The McGraw-Hill Companies, Inc. Permission required to reproduce or display Hartwell et al., 4th edition, Chapter 17 46 The G1-to-S checkpoint is activated by DNA damage Fig. 17.20a Copyright © The McGraw-Hill Companies, Inc. Permission required to reproduce or display Hartwell et al., 4th edition, Chapter 17 47 Disruption of the G1-to-S checkpoint in p53-deficient cells can lead to amplified DNA Tumor cells often have homogenously staining regions (HSRs) or small, extrachromosomal pieces of DNA (double minutes) Fig. 17.20b Copyright © The McGraw-Hill Companies, Inc. Permission required to reproduce or display Hartwell et al., 4th edition, Chapter 17 48 Disruption of the G1-to-S checkpoint in p53-deficient cells can lead to many types of chromosome rearrangements Fig. 17.20c Copyright © The McGraw-Hill Companies, Inc. Permission required to reproduce or display Hartwell et al., 4th edition, Chapter 17 49 Checkpoints acting at the G2-to-M cell-cycle transition or during M phase Fig. 17.21 Copyright © The McGraw-Hill Companies, Inc. Permission required to reproduce or display Hartwell et al., 4th edition, Chapter 17 50 The necessity of checkpoints Checkpoints are not essential for cell division Cells with defective checkpoints are viable and divide at normal rates • But, they are much more vulnerable to DNA damage than normal cells Checkpoints help prevent transmission of three kinds of genomic instability (Fig 17.22) • Chromosome aberrations • Changes in ploidy • Aneuploidy Copyright © The McGraw-Hill Companies, Inc. Permission required to reproduce or display Hartwell et al., 4th edition, Chapter 17 51 Three classes of error lead to aneuploidy in tumor cells Fig. 17.22a Copyright © The McGraw-Hill Companies, Inc. Permission required to reproduce or display Hartwell et al., 4th edition, Chapter 17 52 Chromosome painting can be used to detect chromosome rearrangements Chromosomes from normal cells Chromosomes from tumor cells Fig. 17.22 Copyright © The McGraw-Hill Companies, Inc. Permission required to reproduce or display Hartwell et al., 4th edition, Chapter 17 53