Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Butyric acid wikipedia , lookup
Citric acid cycle wikipedia , lookup
Fatty acid metabolism wikipedia , lookup
Ribosomally synthesized and post-translationally modified peptides wikipedia , lookup
Fatty acid synthesis wikipedia , lookup
Nucleic acid analogue wikipedia , lookup
Point mutation wikipedia , lookup
Peptide synthesis wikipedia , lookup
Metalloprotein wikipedia , lookup
Catalytic triad wikipedia , lookup
Proteolysis wikipedia , lookup
Genetic code wikipedia , lookup
Biochemistry wikipedia , lookup
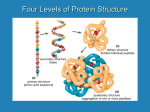
Amino Acids “When you understand the amino acids, you understand everything” ©CMBI 2001 Amino Acids Proteins are macromolecules made up from 20 different amino acids. The heart of the amino acid is the so-called C. To which are bound: an amino group, a carboxyl group, a hydrogen, and the side chain. O H2N C CH OH R The C, C, N and O atoms are called backbone atoms. R denotes any one of the 20 possible side chains. ©CMBI 2001 Amino Acids Backbones run from the amino to the carboxy end, or in other words, from N-terminus to C-terminus. ©CMBI 2001 Zwitterion At pH 7, the amino group and the carboxyl groups are both ionised. In this state the amino group is protonated and thus positive, and the carboxyl group is de-protonated, and thus negative. O H3N An amino acid in this polarised state is called a zwitterion. + C CH O R ©CMBI 2001 - Di-peptide Amino acids bind, to form a protein. Upon binding, two protons from the NH3 and one oxygen from the carboxyl join to form a water. So the peptide bond has at the one side a C=O and at the other side an N-H. Only the ends of the chain are NH3 or carboxylic. ©CMBI 2001 The Peptide Bond H2N f CH R1 phi () psi () omega () O R2 C CH OH N C H O = torsion angle around N-CA bond = torsion angle around CA-C bond = connection between two amino acids = peptide bond (always close to 180 degrees, sometimes close to zero) ©CMBI 2001 20 Amino Acids The side chains, R, determine the differences in the structural and chemical properties of the 20 ‘natural’ amino acids. The 20 amino acids can be classified as follows: Aliphatic/hydrophobic Polar Alcoholic Sulfur-containing Aromatic Charged Special Ala, Leu, Ile, Val Asn, Gln Ser, Thr, (Tyr) Met, Cys Phe, Tyr, Trp, (His) Arg, Lys, Asp, Glu, (His) Gly (no R), Pro (cyclic, imino-acid) Several amino acids belong in more than one category. ©CMBI 2001 Amino Acid Sequence The amino acid sequence (also called primary structure) of a protein is the order of the amino acids in the protein chain. The sequence is always read from the N-terminus to the Cterminus of the protein. For example: +H3N-Lys-Val-Phe-Ala-Met-Cys-Leu-Leu-Arg-Val-COO- Or (in one-lettercode): KVFAMCLLRV ©CMBI 2001 Direction of Protein Chain Lys-Val Val-Lys ©CMBI 2001 The 20 Amino Acids A C D E F G H I K L M N P Q R S T V W Y Ala Cys Asp Glu Phe Gly His Ile Lys Leu Met Asn Pro Gln Arg Ser Thr Val Trp Tyr Alanine Cysteine Aspartic acid (Aspartate) Glutamic acid (Glutamate) Phenylalanine Glycine Histidine Isoleucine Lysine Leucine Methionine Asparagine Proline Glutamine Arginine Serine Threonine Valine Tryptophan Tyrosine ©CMBI 2001 The 20 Amino Acids ©CMBI 2001 Amino Acid Characteristics There are many of ways to characterize the properties of amino acids. The ones most useful and most commonly used are: Hydrophobicity Size Charge Secondary structure preference Alcoholicity Aromaticity And on top of that there are some special characteristics like bridge forming by cysteines, rigidity of prolines, titrating at physiological pH of histidine, flexibility of glycines, etc. ©CMBI 2001 Hydrophobicity Hydrophobicity is the most important characteristic of amino acids. It is the hydrophobic effect that drives proteins towards folding. Actually, it is all done by water. Water does not like hydrophobic surfaces. When a protein folds, exposed hydrophobic side chains get buried, and release water of its sad duty to sit against the hydrophobic surfaces of these side chains. Water is very happy in bulk water because there it has on average 3.6 H-bonds and about six degrees of freedom. So, whenever we discuss protein structure, folding, and stability, it is all the entropy of water, and that is called the hydrophobic effect. ©CMBI 2001 Secondary Structure Preference Amino acids form chains, the sequence or primary structure. These chains fold in -helices, b-strands, b-turns, and loops (or for short, helix, strand, turn and loop), the secondary structure. These secondary structure elements fold further to make whole proteins, but more about that later. There are relations between the physico-chemical characteristics of the amino acids and their secondary structure preference. I.e., the b- branched residues (Ile, Thr, Val) like to sit in b-strands. We will now discuss the 20 ‘natural’ amino acids, and we will later return to the problem of secondary structure preferences. ©CMBI 2001 A Ala Alanine Alanine is a small, hydrophobic residue. Its side chain, R, is just a methyl group. Alanine likes to sit in an alpha helix,it doesn’t like beta strand very much, but it hates beta-turns. If you want to mutate a residue, but you don’t have a good plan about what to replace it with, take alanine. ©CMBI 2001 C Cys Cysteine Cysteine is a small till intermediately large hydrophobic residue. It doesn’t like the alpha helix, but doesn’t mind strand and turn. It can form Cys-Cys bridges with other cysteines. It can bind metals (especially Zn and Cu). The S-H group is very reactive. ©CMBI 2001 D Asp Aspartic acid Aspartic acid, or aspartate, is an intermediately large, hydrophilic, negatively charged residue. Its side chain normally titrates at pH 4.5. It likes to sit near the N-terminus of a helix, and in turns. It hates strands.It often occurs in active sites. It can bind ions (mainly Ca). ©CMBI 2001 E Glu Glutamic acid Glutamic acid, or glutamate, is a large, hydrophilic, negatively charged residue. Its side chain titrates at pH 4.6. It loves the helix, doesn’t mind being in a strand, but is not so good for a turn. ©CMBI 2001 F Phe Phenylalanine Phenylalanine is a large, hydrophobic, aromatic residue. It is good for a strand, it doesn’t mind sitting in a helix, but it hates the turn. ©CMBI 2001 G Gly Glycine Glycine is the smallest residue. It doesn’t have a side chain, so its hydrophobicity is a bit undetermined, but you can call it hydrophobic or assign it an intermediate hydrophobicity. The fact that it doesn’t have a side chain means that its backbone is very flexible so that it can make backbone turns that other residues cannot make. It is very bad for helix, bad for strand, but it is the star of the turns. ©CMBI 2001 H His Histidine Histidine is very special. It is hydrophilic due to the two nitrogens in its side chain. Both nitrogens can titrate (the first one at pH 6.2). It is a little bit aromatic. It is often seen in active sites.It is neutral at physiological pH, but it can easily become positive, and occasionally even negative. It can bind metal ions (mainly Zn, Ni, Cu). It is not particularly picky about its secondary structure. ©CMBI 2001 I Ile Isoleucine Isoleucine is an intermediately large, hydrophobic residue. It is beta branched which means that it likes to sit in a strand. It doesn’t mind sitting in a helix either, but it cries its eyes out in a turn. ©CMBI 2001 K Lys Lysine Lysine is a large, hydrophilic, positively charged residue. It is not a good strand residue, but it doesn’t mind sitting in a helix or in a turn. Its side chain is very long and flexible. ©CMBI 2001 L Leu Leucine Leucine is an intermediately large, hydrophobic residue that really loves to sit in a helix. It is also good for a strand, but it hates turns. ©CMBI 2001 M Met Methionine Methionine is a large, sulphur containing, hydrophobic residue. It loves helices, doesn’t mind sitting in a strand, but it hates turns. Methionine can bind metals with its sulphur, but this sulphur is much less reactive than the sulphur in cysteine. It is often the first residue of a molecule. The N-terminus is mostly positive and thus mostly at the surface. Therefore, methionine, despite being hydrophobic, is often at the surface. We call this a forced marriage. ©CMBI 2001 N Asn Asparagine Asparagine is an intermediately large, polar residue. It hates the helix, is mildly un-amused in a strand, but it loves the turn. It can bind metal ions (Ca), but doesn’t do that as well as its isosteric partner aspartic acid. ©CMBI 2001 P Pro Proline In proline, the side chain is connected to the backbone at two places: the C and the N. Therefore, it is not an amino acid, but an imino acid. Unless it is the N-terminal residue, proline does not have a backbone proton, and thus is not good for helices and strands. Due to the extra covalent bond, proline is already pre-bend, and thus good for turns. Even though it is very hydrophobic, it often sits at the surface ©CMBI 2001 Q Gln Glutamine Glutamine is a large, polar residue. It is not very picky about its secondary structure. It is isosteric with glutamic acid. ©CMBI 2001 R Arg Arginine Arginine is a big, hydrophilic, positively charged residue. Its side chain contains a so-called guadinium group that is rigid. It is not picky about its secondary structure. ©CMBI 2001 S Ser Serine Serine is a small, alcoholic residue of intermediate hydrophobicity. It is not too happy in helices and strands, but it loves to sit in turns. It often forms the active site of an enzyme together with histidine and aspartic acid. It is occasionally involved in metal (Ca) binding. ©CMBI 2001 T Thr Threonine Threonine is a small, alcoholic residue of intermediate hydrophobicity. It is betabranched and thus good for beta strands. It doesn’t care about helices or turns. It is occasionally involved in metal (Ca) binding. ©CMBI 2001 V Val Valine Valine is a small hydrophobic residue. It is beta-branched and thus good for beta strands. It is isosteric with threonine. Valine doesn’t care about helices, but it hates turns. ©CMBI 2001 W Trp Tryptophan Tryptophan is the biggest residue. It is aromatic. Despite that the nitrogen in the five-ring is donor for hydrogen bonds, it is very hydrophobic. It doesn’t care about helices or turns, but it loves strands. ©CMBI 2001 Y Tyr Tyrosine Tyrosine is a large, aromatic, alcoholic residue of intermediate hydrophobicity. It is not so happy in a helix, indifferent about turns, and it loves a strand. ©CMBI 2001 Secondary Structure Preferences Alanine Arginine Aspartic Acid Asparagine Cysteine Glutamic Acid Glutamine Glycine Histidine Isoleucine Leucine Lysine Methionine Phenylalanine Proline Serine Threonine Tryptophan Tyrosine Valine helix 1.42 0.98 1.01 0.67 0.70 1.39 1.11 0.57 1.00 1.08 1.41 1.14 1.45 1.13 0.57 0.77 0.83 1.08 0.69 1.06 strand 0.83 0.93 0.54 0.89 1.19 1.17 1.10 0.75 0.87 1.60 1.30 0.74 1.05 1.38 0.55 0.75 1.19 1.37 1.47 1.70 turn 0.66 0.95 1.46 1.56 1.19 0.74 0.98 1.56 0.95 0.47 0.59 1.01 0.60 0.60 1.52 1.43 0.96 0.96 1.14 0.50 ©CMBI 2001 Secondary Structure Preferences Alanine Glutamic Acid Glutamine Leucine Lysine Methionine Phenylalanine helix 1.42 1.39 1.11 1.41 1.14 1.45 1.13 strand 0.83 1.17 1.10 1.30 0.74 1.05 1.38 turn 0.66 0.74 0.98 0.59 1.01 0.60 0.60 Subset of helix-lovers. If we forget alanine (I don’t understand that things affair with the helix at all), they share the presence of a (hydrophobic) C-b, C-g and C-d (S-d in Met). These hydrophobic atoms pack on top of each other in the helix. That creates a hydrophobic effect. ©CMBI 2001 Secondary Structure Preferences Isoleucine Leucine Phenylalanine Threonine Tryptophan Tyrosine Valine helix 1.08 1.41 1.13 0.83 1.08 0.69 1.06 strand 1.60 1.30 1.38 1.19 1.37 1.47 1.70 turn 0.47 0.59 0.60 0.96 0.96 1.14 0.50 Subset of strand-lovers. These residues either have in common their b-branched nature (Ile, Thr, Val) or their hydrophobicity (rest). ©CMBI 2001 Secondary Structure Preferences Aspartic Acid Asparagine Glycine Proline Serine helix 1.01 0.67 0.57 0.57 0.77 strand 0.54 0.89 0.75 0.55 0.75 turn 1.46 1.56 1.56 1.52 1.43 Subset of turn-lovers. Glycine is special because it is so flexible, so it can easily make the sharp turns and bends needed in a b-turn. Proline is special because it is so rigid; you could say that it is prebend for the b-turn. Aspartic acid, asparagine, and serine have in common that they have short side chains that can form hydrogen bonds with the own backbone. These hydrogen bonds compensate the energy loss caused by bending the chain into a b-turn. ©CMBI 2001 Sequence Alignment Now that we know everything about the amino acids, we can look at the real goal of this whole excercise: sequence alignments. Which of the following alignments is better, left or right: CWPSAAFPWC CWPT---PWC CWPSAAFPWC CWP---TPWC CWPYAAWPWC CWP---FPWC CWPYAAWPWC CWPF---PWC ©CMBI 2001 Sequence Alignment Don’t forget that we still want to gather information about an unknown protein for which we determined the sequence. To gather that information, we will need databases and sequence alignments. To do these sequence alignments, we need to know everything about the amino acids. And that is what we are working on right now. ©CMBI 2001