Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
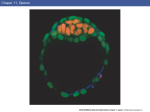
387 Development 109, 387-393 (1990) Printed in Great Britain © T h e Company of Biologists Limited 1990 Fibroblast growth factor during mesoderm induction in the early chick embryo E. MITRANI 1 , Y. GRUENBAUM 2 , H. SHOHAT 1 and T. Z I V 1 2 Departments of Zoology1 and Genetics2, The Institute of Life Sciences, The Hebrew University of Jerusalem, Jerusalem 91904, Israel Summary A chick genomic clone that reveals a high degree of homology to the mammalian and Xenopus bFGF gene has been isolated. The pattern of expression of bFGF has been examined during early chick embryogenesis. RNA blot analysis revealed that chick bFGF is already transcribed at pregastrula stages. Immunolabeling analysis indicated that bFGF protein is present at these early developmental stages and is distributed evenly in the epiblast, hypoblast and marginal zone of the chick blastula. Substances that can inhibit FGF action were applied to early chick blastoderms grown in vitro under defined culture conditions (DCM). Both heparin and suramin were capable of blocking the formation of mesodermal structures in a dose-dependent manner. Our results indicate that FGF-like substances may need to be present for axial structures to develop although they may be acting earlier during the induction of nonaxial mesoderm. Introduction the entire lower layer. Interaction between the epiblast and the hypoblast results in the formation of axial mesoderm through the process of primitive streak formation (Azar and Eyal-Giladi, 1981; Mitrani and Eyal-Giladi, 1981; Mitrani et al. 1983). In the present work, we have isolated a chick genomic clone that reveals a high degree of homology to the mammalian and Xenopus bFGF gene. We have studied the transcription pattern of the chick bFGF homologue and the localization of the bFGF protein during early stages of chick development. We have also performed experiments in which agents that can block FGF-like substances, were applied to early chick blastoderms grown in vitro under defined culture conditions (DCM). Application of both heparin and suramin blocked the formation of mesodermal structures in a dose-dependent manner. Basic fibroblast growth factor (bFGF) can induce the formation of non-axial mesoderm in ectodermal animal caps of Xenopus blastula (Slack et al. 1987). The limited capacity of bFGF to induce muscle actin, which is the earliest marker for muscle differentiation (Gurdon, 1987), can be increased by transforming growth factor-/? (TGF-/3, Kimelman and Kirschner, 1987). Rosa et al. (1988), however, have claimed that TGF-/32, but not TGF-/31 is active in o^actin induction while addition of bFGF has a small synergistic effect. The Xenopus cell line XTC secretes mesodermal-inducing capacity (Smith, 1987). Partial characterization of the secreted protein showed that its properties closely resemble those of TGF-/3 (Smith et al. 1988). FGF is probably involved in the induction of more ventral non-axial mesodermal structures (Godsave et al. 1988 and see Slack and Isaacs, 1989). 1NT-2 and kFGF, two oncogene-related factors related to FGF induce mesoderm formation in Xenopus animal caps as assayed by cardiac actin mRNA levels (Paterno et al. 1989). Recently, it has been shown that aFGF can induce mesoderm formation in colonies from single cells derived from animal caps of stage 8 Xenopus blastoderms (Godsave and Slack, 1989). The chick blastula [stage XIII of Eyal-Giladi and Kochav (E&K), 1976] is formed of two physically distinct layers. The upper layer comprises the primary ectoderm or epiblast, the marginal zone and the area opaca. The primary endoderm or hypoblast constitutes Key words: FGF, mesoderm induction, heparin, suramin, chick embryo. Materials and methods Library screening A chick genomic library (kindly provided by D. Engel) was screened with a 600 bp human bFGF probe containing the entire protein coding region (Abraham et al. 1986a, 1986b). The hybridization solution contained 37% formamide, 5xSSC, lxDenhardt's, O.lmgmP 1 salmon sperm DNA and ^ n i m i "i1 of labeled probe. Hybridization was performed at 37°C. Washes were 3 times, 20min each with 2xSSC, 0.1 % SDS at room temperature and twice with the 1 388 E. Mitrani and others same solution at 50°C. Radiolabeled probe was prepared according to the method of Feinberg and Vogelstein (1983). DNA sequencing The desired fragments were subcloned into the vector pUC118. The nucleotide sequence of the cloned DNA was determined for both strands using the dideoxy chain termination method of Sanger et al. (1977). Northern blot analysis Northern blot analysis of total RNA was performed as described previously (Mitrani et al. 1987). Briefly, embryonic tissue was digested in proteinase K (200[igml"1) at 42°C for 45min. The digest was extracted twice with phenol-chloroform and precipitated in 0.3 M sodium acetate and 2 volumes of ethanol. Total nucleic acids were resuspended in 10 mM Tris-HCl/lOmM MgCl2, and digested with DNAse I (10//gmr') for 30min at 37°C. The RNA was denatured and electrophoresed on a 1 % a"garose/formaldehyde gel, transferred to a Genescreen (Dnnlop) nylon membrane crosslinked with UV and hybridized to the chicken bFGF probe (pGF0.9). The hybridization solution contained 50% formamide, 5xSSC, lxDenhardt's, O.lmgmP 1 salmon sperm DNA and 5xlCr'disintsminimi" 1 of labeled probe. Hybridization was performed at 42°C. Washes were 3 times, 20min each with 0.3xSSC, 0.1% SDS at 50°C and twice with the same solution at 65 °C. Dissection of embryos Staging of the early blastoderms was done according to EyalGiladi and Kochav (E&K) (1976). Gastrulating blastoderms were staged according to Hamburger and Hamilton (H&H) (1951). When necessary, the marginal zone was dissected from stage X1I1 blastoderms (Kahner et al. 1985). Growing whole blastoderms in vitro We have recently developed a reliable bioassay that allows for the growth of early chick blastoderms, or explants derived from them, in the absence of the vitelline membrane, and under defined culture conditions (DCM). This system allows for the controlled application of chosen factors directly to the blastoderm. The blastoderm is layered onto 1.8% agarose in Roswell Park Memorial Institute (RPMI) medium (Mitrani and Shimoni, 1990). The advantage of this culture system is that factors can be directly applied to the embryo via the culture medium in a dose-controlled fashion. Heparin sodium-salt grade I was obtained from Sigma. Suramin was a gift from Dr I. Vlodavsky. Growing cells in agarose The culture medium used was RPMI 1640 containing 10% fetal calf serum and 1 % penicillin and streptomycin. Cells were grown in agarose as described previously (Mitrani, 1984). Briefly, 0.8 ml of 0.33 % agarose in culture medium was prepared as an underlayer in 35 mm Petri dishes. Stage X-XJI blastoderms (E&K) were incubated in trypsin-EDTA for lOmin and then transferred to culture medium and dissociated by gently forcing them through a fine Pasteur pipette. Cells were plated on the same medium at a final agarose concentration of 0.3% and layered onto the previously agarose-coated Petri dish at 5X104 cells ml"1 (Mitrani, 1984). The cells were incubated at 37°C in 5% CO2 and 100% humidity. Duplicate dishes were plated with varying concentrations of heparin or suramin ranging from 10 to 200 /jgmP 1 . Indirect immunofluorescence Chicken blastoderms were dissected into Ringer's solution and processed for indirect immunofluorescence as described previously (Mitrani, 1982). Briefly, the blastoderms were transferred to a fixative solution containing 9 parts of formalin, 3 parts ethanol and one part acetic acid. Following 30min fixation, the blastoderms were subjected to three rinses, lOmin each, with 70% ethanol and transferred to Ringer's solution. The fixed blastoderms were then carefully placed into a drop of Tissue-Tek II mounting medium (Lab-Tek products), kept at -40°C until frozen and sectioned in a standard cryostat. 8/tm sections were obtained and stained with the anti-bFGF antibodies (kindly provided by Dr I. Vlodavsky and by Dr G. Neufeld), diluted 1:200 to 1:1000. Fluorescein-labeled anti-mouse, for monoclonal antibodies, or anti-rabbit, for polyclonal antibodies, were used as secondary antibodies. In control experiments, normal pool serum was used instead of anti-bFGF antibodies. Also, in a separate control, anti-bFGF antibodies, diluted 1:200, were preincubated with 3j/g of cloned human bFGF protein (kindly provided by Dr G. Neufeld) for 60min at 37°C prior to incubation with the tissue. Results Cloning of a chicken bFGF homologue A chicken genomic library (kindly provided by D. Engel) was screened under low-stringency conditions, using a human bFGF cDNA clone as probe (Abraham et al. 1986a,b). Three phages, which turned out to contain the same insert, 4.2kb in length, were picked. Fig. 1A shows a partial restriction map of this clone A EcoRI Bglll Pst I Bgl II Eco Rl B cgtatatagga 12 gaataacttaccttacccttgccaatttcgataataactttgtcctgtttttCicagTC 71 AAA CTG CAG CTT CAA GCA GAA GAA AGA GGA GTA GTA TCA ATC AAA lys leu gin leu gin ala glu glu arg gly val val ser ile Iy3 61 ' 70 116 GGC GTA AGT GCA AAC CGC TTT CTG GCT ATG AAG GAG GAT GGC AGA gly val «*r ala asn arg ph« leu ala met lys glu asp gly arg 80 90 161 TTG CTG GCA CTG otaascagcattJxttaatgtatctctctgtatcttactgcta leu leu ala l«u 94 216 ggtcctcttttcggtttgtgcgttgcataggggagaaattaagctcggaaaaaacttgt 275 300 gtcaattaagagagagatttctgttg Fig. 1. Restriction map and partial sequence of pGF4.2 clone. (A) The exon part is shown as an open box. (B) The putative protein translation of bFGF sequence is shown under the sequence. The three amino acids substituted in the chicken compared to the mammalian bFGF are shown in bold letters. Also shown underlined, the nucleotides that are conserved in the chicken and the human second intron. bFGF during chick mesoderm induction 389 -pGF4.2-. The pstl-Bglll fragment -pGF0.9-, derived from one of the clones, was used as probe in this study. The entire coding region of the putative chicken bFGF second exon contained in pGF4.2 and flanking regions at both ends are shown in Fig. IB. The homology between the chicken exon sequence and the corresponding bFGF exon sequence in other species is 85 % at the nucleotide level and 86-91 % at the amino acid level (Fig. IB). Residue Cys78 in human is not essential for bFGF biological activity (Seno et al. 1988) and is not conserved either in chicken or Xenopus [the chicken Ser78 is substituted in Xenopus by Thr (Kimmelman and Kirschner, 1987)]. The change of Ser94 to Leu is conserved both in chicken and Xenopus. Tyr82, which is conserved in all bFGF genes analyzed to date is substituted in the chicken by Phe. All bFGF genes analyzed so far contain a second exon of identical size. Also, the first seven nucleotides of the putative chicken second intron are identical to those of the human second intron (Fig. IB). After the first 14 nucleotides, the two intron sequences completely diverge. Transcription of bFGF during embryonic development FGF transcription was analyzed in early chicken embryos by the Northern blot technique, using pGF0.9 as probe. About one hundred stage XIII blastoderms (E&K) were dissected in order to obtain enough RNA from the marginal zone region. RNA samples from stages X-XII (E&K) and early primitive streak stages were also tested. The lane in Fig. 2 contains 5 ^ig of total RNA from the marginal zone region. Three weak transcripts 7.8, 1.9 and 1.5 kb in size, could be detected only after exposing the autoradiogram for two weeks. The hybridization signal with RNA prepared from whole embryos at the above stages of development was below our level of detection. When the filter was reprobed with a-actin probe, the hybridization signal was stronger in the lane containing RNA derived from whole embryos compared to that of the marginal zone (data not shown). kb 7.8- t 1.91.5- Fig. 2. bFGF RNA is present in the marginal zone of stage XIII blastoderms. Northern blot analysis of marginal zone of stage XIII blastoderms. 5j*g of total RNA were loaded on the lane and pGF0.9 fragment was used as a probe. The length of the hybridized transcripts in kb is marked on the left. Three faint bands (7.8, 1.9 and 1.5 kb) could be discerned after exposure of the autoradiogram for 14 days. Fig. 3. bFGF protein is expressed in the chick blastula. Indirect immunofluorescence analysis of bFGF in stage XII blastoderms. An antibody that recognizes the M-terminus of bFGF was used as the primary antibody (35). Transverse section through the central region (A) and the more peripheral region (B) of a stage XII blastoderm. bFGF immunostaining pattern seems to be evenly distributed between the epiblast (e), the emerging hypoblast (h) and the marginal zone (mz). Control section is shown in C Magnification x800. 390 E. Mitrani and others Expression of the protein bFGF during early embryonic stages Monoclonal antibodies or polyclonal antibodies directed against the bFGF protein were used for indirect immunofiorescence experiments. Various antibodies were tested. In particular, a polyclonal antibody raised against a synthetic peptide corresponding to the bovine bFGF amino terminal positions 1 to 12 (R4/R6 kindly provided by Dr I. Vlodavsky, Vlodavsky et al. 1987) and a polyclonal antibody also directed against the Nterminus of bovine bFGF (kindly provided by Dr G. Neufeld, (Schweigerer et al. 1987)), gave the more consistent results. We found positive labeling already by stage XI (E&K). The staining seemed to be distributed evenly throughout most of the blastodermic area with some scattered cells giving a higher signal but distributed evenly throughout the epiblast, the emerging hypoblast and the marginal zone (Fig. 3A,B). During primitive streak formation, with the onset of migration of mesenchymal cells through the primitive streak, no significant change in the labeling pattern could be observed (data not shown). Two sets of controls were also performed. The first control entailed replacing the anti-bFGF antibody with pooled rabbit serum. The second control entailed preincubating the anti-bFGF antibody with excess human bFGF, before applying it to the tissue section. No positive immunofluorescence signal could be detected in any control experiment. Instead a very weak uniform background was observed (Fig. 3C). Effect of FGF- inhibitors on early blastoderm development Blastoderms from preblastula to gastrula stages (stages X-XIII (E&K) and stages 1-3 (H&H)) were grown in DCM in the presence of heparin or suramin. Both substances are known to block bFGF interaction with its receptor (Neufeld and Gospodarowicz, 1985, 1987). Blastoderms treated with heparin or with suramin at doses ranging from 1 to 200 ^g ml" 1 responded in a dose-dependent manner (Table 1 and Fig. 4). Pregastrula blastoderms when grown in the presence of heparin at a dose of 50/.igm\~l were incapable of forming a primitive streak in 42 % of cases and when treated with heparin at 100/UgmF1 in 72% of cases (Table 1). Gastrulating blastoderms, when treated at SO^gml"1, continued through gastrulation, formed a full-length primitive streak and in 60 % of cases developed further. 10 % were arrested after forming a fulllength notochord but no somites, whilst the remainder 50% continued to develop into normal embryos. At 100 ^g ml" 1 , gastrulating blastoderms developed a fulllength primitive streak in 75 % of the cases but in the remaining 25 % formed a full-length notochord but were incapable of forming somites. Suramin-treated blastoderms behaved similarly except that the effects were observed at slightly lower doses (Table 1A and see Fig. 5). In contrast whole stage XI-XIII blastoderms when grown in DCM for 48 h developed into normal embryos in 77 % of the cases. The rate of development corresponded to those of normal blastoderms grown in ovo (Figs 4A,C). Table 1. (A) Maximum degree of axial mesoderm development of stage X-XII blastoderms expressed as a percentage Factor Oigml"1) no axis primitive streak chorda chorda+somites Stage n (%) (%) (%) (%) X-XII - 35 13 2 8 77 25 7 21 18 3 10 4 20 29 42 72 _ _ - 8 42 28 17 0 40 75 10 20 23 16 10 5 20 15 13 69 100 0 0 5 16 0 0 0 heparin X-XII Primitive streak 10 25 50 100 25 50 100 X-XII Primitive streak suramin 1 10 25 50 100 25-50 4 68 0 29 9 0 0 10 14 11 100 50 0 25 0 10 38 25 0 40 (B) Cloning efficiency of stage XII celb as percentage of untreated celb Factor added (,/gmr1) Goning efficiency (%) heparin 200 100 suramin 200 100 80 70 33 6 0 60 bFGF during chick mesoderm induction 391 Fig. 4. Suramin and heparin block the formation of axial structures. Control and treated stage XII blastoderms grown in DCM. (A) Untreated blastoderm grown for 24 h has reached the characteristic head process stage. (B) Blastoderm grown in the presence of lO^gml"1 suramin for 24 h: no axis develops. (C) Untreated blastoderm grown for 48 h: a normal stage 13 (H&H) embryo develops. (D) Blastoderm grown in the presence of 10/igml"1 suramin for 48 h: a very rudimentary primitive streak develops. (E) Blastoderm grown in the presence of 25 jjg ml"1 heparin for 48 h: no axis develops. (F) Blastoderm grown in the presence of 25;/gml"1 suramin for 48 h: no axial structures develop. Magnification x!5. Single cells derived from either pregastrulation or gastrulation blastoderms were grown as colonies in soft agar in the presence of either heparin or suramin. Concentrations of each substance as high as 200 jig ml"1, did not inhibit agar growth of the early cells (Table IB). Discussion We have isolated a chick genomic clone that reveals a high degree of homology to the mammalian and Xeno- pus bFGF gene. A fragment of this clone, containing the full second exon, was used as probe in RNA hybridization experiments. We found that bFGF is already expressed at the blastula stage. We could only detect RNA messages in the marginal zone of stage XIII blastoderms. In contrast, we detected bFGF protein throughout the developing blastoderm. One possible explanation for that difference would be that bFGF transcripts are present in other embryonic cells, but in amounts that are below our level of detection. It is nevertheless interesting that bFGF transcripts were 392 E. Mitrani and others Fig. 5. Different levels of abnormalities caused by suramin. Whole stage XII blastoderms grown in DCM for 48 h in the presence of 25 fig ml"' suramin for48h. (A) No axial structures develop; (B) an embryonic axis develops but there are no somites; (C) both a notochord and somites developed from this treated blastoderm. Magnification x!5, detected in the marginal zone since this region has the capacity to regenerate a hypoblastic layer, which, when in contact with a stage XIII epiblast, can induce the formation of axial structures (Azar and Eyal-Giladi, 1979). Slack and Isaacs (1989) have recently reported that bFGF-like molecules are present in Xenopus blastulae. There have also been previous attempts at examining FGF levels in the early chick embryo (Seed et al. 1988). These workers, however, reported that FGF is present in the chick egg yolk and egg white, but did not address the question of whether the early embryo itself is synthesizing FGF. Direct experiments on induction of non-axial mesoderm cannot be performed in the chick since, by the time the epiblast and hypoblast tissues sort themselves out into two separate entities, non-axial mesoderm has already been predetermined (Azar and Eyal-Giladi, 1979; Mitrani and Eyal-Giladi, 1982). Since bFGF is already expressed in the chick gastrula, we had to resort to applying agents that block bFGF action in order to assay for the possible function of bFGF. Neither heparin nor suramin are specific for bFGF but both are known to block bFGF action (Neufeld and Gospodarowicz, 1985, 1987; Slack et al. 1987; Moscatelli and Quarto, 1989). The two substances, although rather different, had very similar effects on early blastoderms. Application of either heparin or suramin also resulted in mesoderm malformations when applied after the primitive streak was already induced, suggesting that FGF-like substances may be required throughout the process of axial mesoderm formation. It is unlikely that both substances acted in a nonspecific toxic manner since, when applied at higher concentrations to cells derived from the same blastoderms and grown in agarose, anchorage-independent growth (AIG) was not inhibited. This behavior should be contrasted with the behavior of the same cells when treated with retinoic acid (RA). RA can block AIG at doses as low as 10~ 9 M whilst effects on whole blastoderms can only be felt at concentrations which are two orders of magnitude higher (Mitrani and Shimoni, 1989). The doses of heparin that were shown to inhibit the formation of axial structures although relatively high were similar to those that were shown previously to inhibit FGF induction of mesoderm in Xenopus animal caps in transfilter experiments (Slack et al. 1987). Heparin at lmgml" 1 had no toxic effects in Xenopus isolated animal caps, and did not block the formation of mesoderm in animal-vegetal combinations (Slack et al. 1987). If an animal cap of a Xenopus blastula is grown in isolation, differentiation of non-axial mesoderm does not take place. In the chick, however, isolated epiblasts can give rise to non-axial mesoderm (blood islands) without the addition of exogenous factors (Azar and Eyal-Giladi, 1979; Mitrani and Eyal-Giladi, 1982; Stern and Canning, 1990). Presumably by stage XI-XII the signals responsible for the induction of non-axial mesoderm are already present in the chick blastoderm. The fact that bFGF transcripts and protein are already present in pregastrula blastoderms suggests that in the chick bFGF may be involved, as in Xenopus, in the induction of non-axial mesoderm. We have recently shown that bFGF cannot induce axial mesoderm (notochord and somites) in isolated chick epiblasts (Mitrani and Shimoni, 1990). In contrast, we showed that conditioned medium derived from the Xenopus XTC cell line (XTC CM) (Smith, 1987), induced the formation of mesoderm which became organized into structures such as a full-length notochord and rows of bilaterally symmetric somites (Mitrani and Shimoni, 1990). These experiments suggested that bFGF is not sufficient to induce axial structures in the chick. The blocking experiments bFGF during chick mesoderm induction 393 reported here, however, indicate that FGF-like substances may still be necessary for that process to take place. FGF-like substances may thus be acting throughout the whole process of mesoderm formation: earlier in development in the induction of non-axial mesoderm and later, in combination with inductive factors provided by the hypoblast (Mitrani and Eyal-Giladi, 1981; Mitrani and Shimoni, 1990), during the generation of axial mesodermal structures. We wish to thank Dr I. Vlodavski for bFGF antibodies, bovine bFGF protein and suramin, Dr G. Neufeld for bFGF antibodies and for providing the human bFGF cDNA clone, Y. Grinfeld and I. Raibstein for help during the cloning process and Dr A. Fainsod for his comments. This work was partly supported by the Israel Academy of Sciences (E.M and Y. G) and by a grant from the American-Israel Binational Science foundation No. 85-00296/3 (E.M). References ABRAHAM, J. A., MERCIA, A., WHANG J. L., TUMOLO, A., FRIEDMAN, J., HJERRILD, K. A., GOSPODAROWICZ, D. AND FIDDES, J. C. (1986a). Nucleotide sequence of a bovine clone encoding the angiogenic protein, basic fibroblast growth factor. Science 233, 545-548. ABRAHAM, J. A., WHANG, J. L., TUMOLO, A., MERGIA, A., FRIEDMAN, J., GOSPODAROWICZ, D. AND FIDDES, J. C. (19866). Human basic fibroblast growth factor: nucleotide sequence and genomic organization. EMBO J. 5, 2523-2528. AZAR, Y. AND EYAL-GILADI, H. (J979). Marginal zone cells-the primitive streak inducing component of the primary hypoblast in the chick. J. Embryol. exp. Morph. 52, 79-88. AZAR, Y. AND EYAL-GILADI, H. (1981). Interaction of epiblast and hypoblast in the formation of the primitive streak and the embryonic axis in chick, as revealed by hypoblast-rotation experiments. J. Embryol. exp. Morph. 61, 133-144. EYAL-GILADI, H. AND KOCHAV, S. (1976). From cleavage to primitive streak formation. A complementary normal table and a new look at the first stages of the development of the chick. Devi Biol. 49, 321-337. FEINBERG, A. P. AND VOCELSTEIN, B. (1983). A technique for radiolabeling DNA restriction endonuclease fragments to high specific activity. Anal. Biochem. 132, 6—13. GODSAVE, S. F., ISAACS, H. V. AND SLACK, J. M. W. (1988). Mesoderm-inducing factors: a small class of molecules. Development 102, 555-566. GODSAVE, S. F. AND SLACK, J. M. (1989). Clonal analysis of mesoderm induction in Xenopus laevis. Devi Biol 134, 486-490. GURDON, J. B. (1987). Embryonic induction-molecular prospects. Development 99, 285-306. HAMBURGER, V. AND HAMILTON, H. L. (1951). A series of normal stages in the development of the chick embryo. J. Morph. 88, 49-92. KAHNER, O., MITRANI, E. AND EYAL-GILADI, H. (1985). Developmental potencies of area opaca and marginal zone areas of early chick blastoderms. J. Embryol. exp. Morph. 89, 235-241. KIMELMAN, D. AND KIRSCHNER, M. (1987). Synergistic induction of mesoderm by FGF and TGF-/3 and the identification of an mRNA coding for FGF in the early Xenopus embryo. Cell 51, 869-877. MITRANI, E. (1982). Primitive streak-forming cells of the chick invaginate through a basement membrane. Wilhelm Roux' Arch. devl Biol. 191, 320-324. MITRANI, E. (1984). Early chick embryonic cells can form clones in agarose. Expl Cell Res. 152, 148-153. MITRANI, E. AND EYAL-GILADI, H. (1981). Hypoblastic cells can form a disk inducing an embryonic axis in chick epiblast. Nature 289,800-802. MITRANI, E. AND EYAL-GILADI, H. (1982). Early chick embryonic cells in culture. Differentiation 21, 56-61. MITRANI, E. AND SHIMONI, Y. (1989). Retinoic acid inhibits growth in agarose of early chick embryonic cells and may be involved in regulation of axis formation. Development 107, 275-281. MITRANI, E. AND SHIMONI, Y. (1990). Induction by soluble factors of organized axial structures in chick epiblasts. Science 247, 1092-1094. MITRANI, E., SHIMONI, Y. AND EYAL-GILADI, H. (1983). Nature of the hypoblastic influence on the chick embryo epiblast. J. Embryol. exp. Morph. 75, 21-30. MOSCATELLI, D. AND QUARTO, N. (1989). Transformation of NIH 3T3 cells with basic fibroblast growth factor or the hst/k-fgf oncogene causes downregulation of the fibroblast growth factor receptor: reversal of morphological transformation and restoration of receptor number by suramin. J. Cell Biol. 109, 2519-2527. NEUFELD, G. AND GOSPODAROWICZ, D. (1985). The identification and partial characterization of the fibroblast growth factor receptor of baby hamster kidney cells. J. biol. Chem. 260, 13860-13868. NEUFELD, G. AND GOSPODAROWICZ, D. (1987). Protamine sulfate inhibits mitogenic activities of the extracellular matrix and fibroblast growth factor, but potentiates that of epidermal growth factor. J. cell. Physiol. 132, 287-294. PATERNO, G. D . , GILLESPIE, L. L., DIXON, M. S., SLACK, J. W. M. AND HEATH, J. K. (1989). Mesoderm-inducing properties of INT2 and kFGF: two oncogene-encoded growth factors related to FGF. Development 106, 79-83. ROSA, F., ROBERTS, A. B., DANIELPOUR, D., DART, L. L. M., SPORN, B. AND DAWID, I. G. (1988). Mesoderm induction in amphibians: The role of TGF-/?2-like factors. Science 239, 783-785. SANGER, F., NICKLEN, S. AND COULSON, A. R. (1977). DNA sequencing with chain-terminating inhibitors. Proc. natn. Acad. Sci. U.S.A. 74, 5463-5467. SCHWEIGERER, L . , NEUFELD, G . , FRIEDMAN, J . , ABRAHAM, J. A . , FIDDES, J. C. AND GOSPODAROWICZ, D. (1987). Capillary endothelial cells express basic fibroblast growth factor, a mitogen that promotes their growth. Nature 325, 257-259. SEED, J., OLWIN, B. B. AND HAUSCHKA, S. D. (1988). Fibroblast growth factor levels in the whole embryo and limb bud during chick development. Devi Biol. 128, 50-57. SENO, M., SASADA, R., IWANE, M., SUDO, K., KUROKAWA, T. AND ITO, K. (1988). Stabilizing basic fibroblast growth factor using protein engineering. Biochem. Biophys. Res. Commun. 151, 701-708. SLACK, J. M. W., DARLINGTON, B. G., HEATH, J. K. AND GODSAVE, S. F. (1987). Mesoderm induction in early Xenopus embryos by heparin-binding growth factors. Nature 326, 197-200. SLACK, J. M. W. AND ISAACS, H. V. (1989). Presence of basic fibroblast growth factor in the early Xenopus embryo. Development 105, 147-153. SMITH, J. C. (1987). A mesoderm-inducing factor is produced by a Xenopus cell line. Development 99, 3-14. SMITH, J. C , YAQOOB, M. AND SYMES, K. (1988). Purification, partial characterization and biological effects of the XTC mesoderm-inducing factor. Development 103, 591-600. STERN, C. D. AND CANNING, D. R. (1990). Origin of cells giving rise to mesoderm and endoderm in chick embryo. Nature 343, 273-275. VLODAVSKY, I . , FOLKMAN, J . , SULLIVAN, R . , F R I D M A N , R . , ISHAIMICHAEU, R., SASSE, J. AND KLAGSBURN, M. (1987). Endothelial cell-derived basic fibroblast growth factor: Synthesis and deposition into subendothelial extracellular matrix. Proc. natn. Acad. Sci. U.S.A. 84, 2292-2296. MITRANI, E., COFFIN, J., BOEDTKER, H. AND DOTY, P. (1987). Rous sarcoma virus is integrated but not expressed in chicken early embryonic cells. Proc. natn. Acad. Sci. U.S.A. 84, 2781-2784. (Accepted 2 March 1990)