Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Nucleic acid analogue wikipedia , lookup
Fatty acid synthesis wikipedia , lookup
Point mutation wikipedia , lookup
Ribosomally synthesized and post-translationally modified peptides wikipedia , lookup
Metalloprotein wikipedia , lookup
Genetic code wikipedia , lookup
Peptide synthesis wikipedia , lookup
Protein structure prediction wikipedia , lookup
Proteolysis wikipedia , lookup
Amino acid synthesis wikipedia , lookup
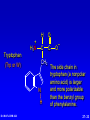
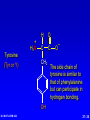
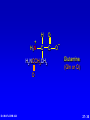
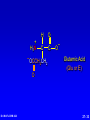
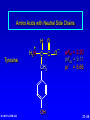
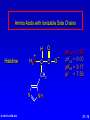
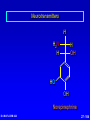
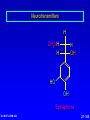
Amino Acids, Peptides, and Proteins Dr. Wolf's CHM 424 27- 1 Objectives Draw a general amino acid and identify the two functional groups common to all. Classify each amino acid according to the chemical nature of its R group. Define the meaning of an essential amino acid. Draw the reaction that joins two amino acids to form a peptide bond. Describe and differentiate primary, secondary, tertiary, and quaternary protein structures. Describe and differentiate co-enzymes and prosthetic groups. List and discuss four forces that stabilize globular protein structure. List important structural similarities and differences between myoglobin and hemoglobin. Describe the mutation present in hemoglobin giving rise to sickle cell disease. Dr. Wolf's CHM 424 27- 2 Classification of Amino Acids Dr. Wolf's CHM 424 27- 3 Fundamentals While their name implies that amino acids are compounds that contain an —NH2 group and a —CO2H group, these groups are actually present as —NH3+ and —CO2– respectively. They are classified as , , , etc. amino acids according the carbon that bears the nitrogen. Dr. Wolf's CHM 424 27- 4 Amino Acids + NH3 CO2– + – H3NCH2CH2CO2 + – H3NCH2CH2CH2CO2 Dr. Wolf's CHM 424 an -amino acid that is an intermediate in the biosynthesis of ethylene a -amino acid that is one of the structural units present in coenzyme A a -amino acid involved in the transmission of nerve impulses 27- 5 The 20 Key Amino Acids More than 700 amino acids occur naturally, but 20 of them are especially important. These 20 amino acids are the building blocks of proteins. All are -amino acids. They differ in respect to the group attached to the carbon. These 20 are listed in Table 27.1. Dr. Wolf's CHM 424 27- 6 H + H3N C O C O – R The amino acids obtained by hydrolysis of proteins differ in respect to R (the side chain). The properties of the amino acid vary as the structure of R varies. Dr. Wolf's CHM 424 27- 7 H + H3N C O C O – R The major differences among the side chains concern: Size and shape Electronic characteristics Dr. Wolf's CHM 424 27- 8 General categories of -amino acids nonpolar side chains polar but nonionized side chains acidic side chains basic side chains Dr. Wolf's CHM 424 27- 9 General categories of -amino acids nonpolar side chains polar but nonionized side chains acidic side chains basic side chains Dr. Wolf's CHM 424 27- 10 Amino Acid R-groups Non-Polar Hydrophobic Polar Charged Uncharged Arginine (+) Glutamic acid (-) Aspartic Acid (-) Lysine (+) Histidine (+) Cysteine Proline Serine Glutamine Asparagine Tryptophan Phenylalanine Isoleucine Tyrosine Leucine Valine Methionine Ambivalent Glycine Threonine Alanine Dr. Wolf's CHM 424 27- 11 Charged Amino Acids + + N N O N - O O O O N Arginine [Arg] Glutamate [Glu] O Aspartate [Asp] N N Lysine [Lys] O +N N O O - N N N Histidine [His] Dr. Wolf's CHM 424 27- 12 Hydrophobic Indexes Arginine Arg [R] -11.2 Glutamic Acid Glu [E]-9.9 Aspartic Acid Asp [D] -7.4 Lysine Lys [K] -4.2 Histidine His [H] -3.3 Cysteine Cys [C] -2.8 Proline Pro [P] -0.5 Serine Ser [S] -0.3 Glutamine Gln [Q] -0.3 Asparagine Asn [N] -0.2 Dr. Wolf's CHM 424 Glycine Gly [G] 0 Threonine Thr [T] 0.4 Alanine Ala [A] 0.5 Methionine Met [M] 1.3 Valine Val [V] 1.5 Leucine Leu [L] 1.8 Tyrosine Tyr [Y] 2.3 Isoleucine Ile [I] 2.5 Phenylalanine Phe [F] 2.5 Tryptophan Trp [W] 3.4 27- 13 Essential amino acids Definition - Those amino acids that cannot be synthesized in the body in sufficient quantities for anabolic needs. In humans, Isoleucine Leucine Valine Tryptophan Methionine Lysine Phenylalanine Threonine Histidine Dr. Wolf's CHM 424 27- 14 H Glycine (Gly or G) + H3N C O C O – H Glycine is the simplest amino acid. It is the only one in the table that is achiral. In all of the other amino acids in the table the carbon is a chirality center. Dr. Wolf's CHM 424 27- 15 H + H3N C O C O – CH3 Alanine (Ala or A) Alanine, valine, leucine, and isoleucine have alkyl groups as side chains, which are nonpolar and hydrophobic. Dr. Wolf's CHM 424 27- 16 H + H3N C O C O – CH(CH3)2 Valine (Val or V) Dr. Wolf's CHM 424 27- 17 H + H3N C O C O – CH2CH(CH3)2 Leucine (Leu or L) Dr. Wolf's CHM 424 27- 18 H + H3N C O C O – CH3CHCH2CH3 Isoleucine (Ile or I) Dr. Wolf's CHM 424 27- 19 H + H3N C O C O – CH3SCH2CH2 Methionine (Met or M) The side chain in methionine is nonpolar, but the presence of sulfur makes it somewhat polarizable. Dr. Wolf's CHM 424 27- 20 H + H2N C O C CH2 H2C C H2 O – Proline (Pro or P) Proline is the only amino acid that contains a secondary amine function. Its side chain is nonpolar and cyclic. Dr. Wolf's CHM 424 27- 21 H + H3N C CH2 O C O – Phenylalanine (Phe or F) The side chain in phenylalanine (a nonpolar amino acid) is a benzyl group. Dr. Wolf's CHM 424 27- 22 H + H3N C O C O – Tryptophan CH2 (Trp or W) N H Dr. Wolf's CHM 424 The side chain in tryptophan (a nonpolar amino acid) is larger and more polarizable than the benzyl group of phenylalanine. 27- 23 General categories of -amino acids nonpolar side chains polar but nonionized side chains acidic side chains basic side chains Dr. Wolf's CHM 424 27- 24 H + H3N C O C O – CH2OH Serine (Ser or S) The —CH2OH side chain in serine can be involved in hydrogen bonding. Dr. Wolf's CHM 424 27- 25 H + H3N C O C O – CH3CHOH Threonine (Thr or T) The side chain in threonine can be involved in hydrogen bonding, but is somewhat more crowded than in serine. Dr. Wolf's CHM 424 27- 26 H + H3N C O C O – CH2SH Cysteine (Cys or C) The side chains of two remote cysteines can be joined by forming a covalent S—S bond. Dr. Wolf's CHM 424 27- 27 H + H3N C O C O – Tyrosine (Tyr or Y) CH2 The side chain of tyrosine is similar to that of phenylalanine but can participate in hydrogen bonding. OH Dr. Wolf's CHM 424 27- 28 H + H3N C H2NCCH2 O C O – Asparagine (Asn or N) O The side chains of asparagine and glutamine (next slide) terminate in amide functions that are polar and can engage in hydrogen bonding. Dr. Wolf's CHM 424 27- 29 H + H3N C H2NCCH2CH2 O C O – Glutamine (Gln or Q) O Dr. Wolf's CHM 424 27- 30 General categories of -amino acids nonpolar side chains polar but nonionized side chains acidic side chains basic side chains Dr. Wolf's CHM 424 27- 31 H + H3N – C OCCH2 O C O – Aspartic Acid (Asp or D) O Aspartic acid and glutamic acid (next slide) exist as their conjugate bases at biological pH. They are negatively charged and can form ionic bonds with positively charged species. Dr. Wolf's CHM 424 27- 32 H + H3N – C OCCH2CH2 O C O – Glutamic Acid (Glu or E) O Dr. Wolf's CHM 424 27- 33 General categories of -amino acids nonpolar side chains polar but nonionized side chains acidic side chains basic side chains Dr. Wolf's CHM 424 27- 34 H Lysine (Lys or K) + H3N C O C O – + CH2CH2CH2CH2NH3 Lysine and arginine (next slide) exist as their conjugate acids at biological pH. They are positively charged and can form ionic bonds with negatively charged species. Dr. Wolf's CHM 424 27- 35 H Arginine (Arg or R) + H3N C O C O – CH2CH2CH2NHCNH2 + NH2 Dr. Wolf's CHM 424 27- 36 H Histidine + H3N (His or H) CH2 N Dr. Wolf's CHM 424 C NH O C O – Histidine is a basic amino acid, but less basic than lysine and arginine. Histidine can interact with metal ions and can help move protons from one site to another. 27- 37 Stereochemistry of Amino Acids Dr. Wolf's CHM 424 27- 38 Configuration of -Amino Acids Glycine is achiral. All of the other amino acids in proteins have the L-configuration at their carbon. – CO2 + H3N H R Dr. Wolf's CHM 424 27- 39 R H2 N CH CO2H All DNA encoded aa are CHO All are chiral, except Glycine R=H H OH HO CH2OH H CH2OH D- All DNA encoded aa are usually L- LR CHO = HO CH2OH H (S) - Glyceraldehyde (-) Dr. Wolf's CHM 424 CHO = H2N C CO2H H (L) - Amino Acids (-) 27- 40 Acid-Base Behavior of Amino Acids Dr. Wolf's CHM 424 27- 41 Recall While their name implies that amino acids are compounds that contain an —NH2 group and a —CO2H group, these groups are actually present as —NH3+ and —CO2– respectively. How do we know this? Dr. Wolf's CHM 424 27- 42 aa are high melting point solids! Why? Answer = aa are ionic compounds under normal conditions LOW pH NEUTRAL C O O O R HIGH pH R C OH NH3 ammonium Form R C O NH3 Zwitterion O NH2 Carboxylate Form Isoelectric Point = concentration of zwitterion is at a maximum and the concentration of cations and anions is equal r aa with basic R-groups, we require higher pHs, and for aa with acidic R-groups, we require lower pHs Dr. Wolf's CHM 424 to reach the Isoelectric Point 27- 43 NH 3 CO2 (CH2)2 pH 7 CH CH H3N (CH2)2 CO2 Glu H3N CO2 Lys Isoelectric Point is the pH at which an aa or peptide carries no net charge. i.e. [RCOO-] = [RNH3+] So, for basic R-groups, we require higher pHs, and for acidic R-groups we require lower pHs e.g. Isoelectric point for gly pH = 6.0 Asp pH = 3.0 Lys pH = 9.8 Arg pH = 10.8 Dr. Wolf's CHM 424 27- 44 Properties of Glycine The properties of glycine: high melting point: (when heated to 233°C it decomposes before it melts) solubility: soluble in water; not soluble in nonpolar solvent more consistent with this than this •• O •• + H3NCH2C Dr. Wolf's CHM 424 •• O •• •• •– O• •• •• H2NCH2C •• OH •• 27- 45 Properties of Glycine The properties of glycine: high melting point: (when heated to 233°C it decomposes before it melts) solubility: soluble in water; not soluble in nonpolar solvent more consistent with this •• O •• + H3NCH2C Dr. Wolf's CHM 424 •• •– O• •• called a zwitterion or dipolar ion 27- 46 Acid-Base Properties of Glycine The zwitterionic structure of glycine also follows from considering its acid-base properties. A good way to think about this is to start with the structure of glycine in strongly acidic solution, say pH = 1. At pH = 1, glycine exists in its protonated form (a monocation). •• O •• + H3NCH2C Dr. Wolf's CHM 424 •• OH •• 27- 47 Acid-Base Properties of Glycine Now ask yourself "As the pH is raised, which is the first proton to be removed? Is it the proton attached to the positively charged nitrogen, or is it the proton of the carboxyl group?" You can choose between them by estimating their respective pKas. typical ammonium ion: pKa ~9 Dr. Wolf's CHM 424 •• O •• + H3NCH2C •• typical carboxylic acid: pKa ~5 OH •• 27- 48 Acid-Base Properties of Glycine The more acidic proton belongs to the CO2H group. It is the first one removed as the pH is raised. •• O •• + H3NCH2C Dr. Wolf's CHM 424 •• typical carboxylic acid: pKa ~5 OH •• 27- 49 Acid-Base Properties of Glycine Therefore, the more stable neutral form of glycine is the zwitterion. •• O •• + H3NCH2C •• •– O• •• •• O •• + H3NCH2C Dr. Wolf's CHM 424 •• typical carboxylic acid: pKa ~5 OH •• 27- 50 Acid-Base Properties of Glycine The measured pKa of glycine is 2.34. Glycine is stronger than a typical carboxylic acid because the positively charged N acts as an electron-withdrawing, acid-strengthening substituent on the carbon. •• O •• + H3NCH2C Dr. Wolf's CHM 424 •• typical carboxylic acid: pKa ~5 OH •• 27- 51 Acid-Base Properties of Glycine A proton attached to N in the zwitterionic form of nitrogen can be removed as the pH is increased further. •• O •• + H3NCH2C •• •– O• •• HO – •• O •• •• H2NCH2C •• •– O• •• The pKa for removal of this proton is 9.60. This value is about the same as that for NH4+ (9.3). Dr. Wolf's CHM 424 27- 52 Isoelectric Point pI •• O •• + H3NCH2C •• OH •• pKa = 2.34 •• O •• + H3NCH2C •• •– O• •• pKa = 9.60 •• O •• •• Dr. Wolf's CHM 424 H2NCH2C •• •– O• •• The pH at which the concentration of the zwitterion is a maximum is called the isoelectric point. Its numerical value is the average of the two pKas. The pI of glycine is 5.97. 27- 53 Acid-Base Properties of Amino Acids One way in which amino acids differ is in respect to their acid-base properties. This is the basis for certain experimental methods for separating and identifying them. Just as important, the difference in acid-base properties among various side chains affects the properties of the proteins that contain them. Table 27.2 gives pKa and pI values for amino acids with neutral side chains. Dr. Wolf's CHM 424 27- 54 Amino Acids with Neutral Side Chains H Glycine + H3N C O C O – pKa1 = 2.34 pKa2 = 9.60 pI = 5.97 H Dr. Wolf's CHM 424 27- 55 Amino Acids with Neutral Side Chains H Alanine + H3N C O C O – pKa1 = 2.34 pKa2 = 9.69 pI = 6.00 CH3 Dr. Wolf's CHM 424 27- 56 Amino Acids with Neutral Side Chains H Valine + H3N C O C O – pKa1 = 2.32 pKa2 = 9.62 pI = 5.96 CH(CH3)2 Dr. Wolf's CHM 424 27- 57 Amino Acids with Neutral Side Chains H Leucine + H3N C O C O – pKa1 = 2.36 pKa2 = 9.60 pI = 5.98 CH2CH(CH3)2 Dr. Wolf's CHM 424 27- 58 Amino Acids with Neutral Side Chains H Isoleucine + H3N C O C O – pKa1 = 2.36 pKa2 = 9.60 pI = 5.98 CH3CHCH2CH3 Dr. Wolf's CHM 424 27- 59 Amino Acids with Neutral Side Chains H Methionine + H3N C O C O – pKa1 = 2.28 pKa2 = 9.21 pI = 5.74 CH3SCH2CH2 Dr. Wolf's CHM 424 27- 60 Amino Acids with Neutral Side Chains H Proline + H2N C O C O – pKa1 = 1.99 pKa2 = 10.60 pI = 6.30 CH2 H2C C H2 Dr. Wolf's CHM 424 27- 61 Amino Acids with Neutral Side Chains H Phenylalanine + H3N C CH2 Dr. Wolf's CHM 424 O C O – pKa1 = 1.83 pKa2 = 9.13 pI = 5.48 27- 62 Amino Acids with Neutral Side Chains H Tryptophan + H3N C CH2 O C O – pKa1 = 2.83 pKa2 = 9.39 pI = 5.89 N Dr. Wolf's CHM 424 H 27- 63 Amino Acids with Neutral Side Chains H Asparagine + H3N C O C O – pKa1 = 2.02 pKa2 = 8.80 pI = 5.41 H2NCCH2 O Dr. Wolf's CHM 424 27- 64 Amino Acids with Neutral Side Chains H Glutamine + H3N C H2NCCH2CH2 O C O – pKa1 = 2.17 pKa2 = 9.13 pI = 5.65 O Dr. Wolf's CHM 424 27- 65 Amino Acids with Neutral Side Chains H Serine + H3N C O C O – pKa1 = 2.21 pKa2 = 9.15 pI = 5.68 CH2OH Dr. Wolf's CHM 424 27- 66 Amino Acids with Neutral Side Chains H Threonine + H3N C O C O – pKa1 = 2.09 pKa2 = 9.10 pI = 5.60 CH3CHOH Dr. Wolf's CHM 424 27- 67 Amino Acids with Neutral Side Chains H Tyrosine + H3N C CH2 Dr. Wolf's CHM 424 OH O C O – pKa1 = 2.20 pKa2 = 9.11 pI = 5.66 27- 68 Amino Acids with Neutral Side Chains H Cysteine + H3N C O C O – pKa1 = 1.96 pKa2 = 8.18 pI = 5.07 CH2SH Dr. Wolf's CHM 424 27- 69 Amino Acids with Ionizable Side Chains H Aspartic acid + H3N – C OCCH2 O C O – pKa1 = pKa2 = pKa3 = pI = 1.88 3.65 9.60 2.77 O For amino acids with acidic side chains, pI is the average of pKa1 and pKa2. Dr. Wolf's CHM 424 27- 70 Amino Acids with Ionizable Side Chains H + H3N Glutamic acid – C OCCH2CH2 O C O – pKa1 = pKa2 = pKa3 = pI = 2.19 4.25 9.67 3.22 O Dr. Wolf's CHM 424 27- 71 Amino Acids with Ionizable Side Chains H + H3N C O C O – + CH2CH2CH2CH2NH3 pKa1 = pKa2 = pKa3 = pI = 2.18 8.95 10.53 9.74 Lysine For amino acids with basic side chains, pI is the average of pKa2 and pKa3. Dr. Wolf's CHM 424 27- 72 Amino Acids with Ionizable Side Chains H + H3N C O C O – CH2CH2CH2NHCNH2 pKa1 = pKa2 = pKa3 = pI = 2.17 9.04 12.48 10.76 + NH2 Arginine Dr. Wolf's CHM 424 27- 73 Amino Acids with Ionizable Side Chains H Histidine + H3N C CH2 N Dr. Wolf's CHM 424 O C O – pKa1 = pKa2 = pKa3 = pI = 1.82 6.00 9.17 7.59 NH 27- 74 Synthesis of Amino Acids Dr. Wolf's CHM 424 27- 75 From -Halo Carboxylic Acids O CH3CHCOH + 2NH3 Br H2O O – CH3CHCO + NH4Br +NH3 (65-70%) Dr. Wolf's CHM 424 27- 76 Strecker Synthesis O CH3CH NH4Cl NaCN CH3CHC N NH2 1. H2O, HCl, heat 2. HO– O – CH3CHCO Dr. Wolf's CHM 424 +NH3 (52-60%) 27- 77 Using Diethyl Acetamidomalonate O O C C C CH3CH2O CH3CNH H OCH2CH3 O Can be used in the same manner as diethyl malonate (Section 21.7). Dr. Wolf's CHM 424 27- 78 Example O O CH3CH2OCCCOCH2CH3 H CH3CNH O 1. NaOCH2CH3 2. C6H5CH2Cl O O CH3CH2OCCCOCH2CH3 CH3CNH Dr. Wolf's CHM 424 O CH2C6H5 (90%) 27- 79 O O Example HOCCCOH –CO2 CH2C6H5 H3 N + O HBr, H2O, heat HCCOH H3N + CH2C6H5 O O (65%) CH3CH2OCCCOCH2CH3 CH3CNH Dr. Wolf's CHM 424 O CH2C6H5 27- 80 Reactions of Amino Acids Dr. Wolf's CHM 424 27- 81 Acylation of Amino Group The amino nitrogen of an amino acid can be converted to an amide with the customary acylating agents. O O O + – + H3NCH2CO CH3COCCH3 O O CH3CNHCH2COH Dr. Wolf's CHM 424 (89-92%) 27- 82 Esterification of Carboxyl Group The carboxyl group of an amino acid can be converted to an ester. The following illustrates Fischer esterification of alanine. O + – + H3NCHCO CH3CH2OH CH3 HCl O Cl Dr. Wolf's CHM 424 – + H3NCHCOCH2CH3 CH3 (90-95%) 27- 83 Ninhydrin Test Amino acids are detected by the formation of a purple color on treatment with ninhydrin. O O OH + + H3NCHCO– OH R O O O RCH + CO2 + H2O + Dr. Wolf's CHM 424 O– N O O 27- 84 Some Biochemical Reactions of Amino Acids Dr. Wolf's CHM 424 27- 85 Biosynthesis of L-Glutamic Acid O HO2CCH2CH2CCO2H + NH3 enzymes and reducing coenzymes – HO2CCH2CH2CHCO2 + NH3 This reaction is the biochemical analog of reductive amination (Section 22.10). Dr. Wolf's CHM 424 27- 86 Transamination via L-Glutamic Acid O – HO2CCH2CH2CHCO2 + CH3CCO2H + NH3 L-Glutamic acid acts as a source of the amine group in the biochemical conversion of -keto acids to other amino acids. In the example to be shown, pyruvic acid is converted to L-alanine. Dr. Wolf's CHM 424 27- 87 Transamination via L-Glutamic Acid O – HO2CCH2CH2CHCO2 + CH3CCO2H + NH3 enzymes O HO2CCH2CH2CCO2H – + CH3CHCO2 + NH3 Dr. Wolf's CHM 424 27- 88 Mechanism O – HO2CCH2CH2CHCO2 + CH3CCO2H + NH3 The first step is imine formation between the amino group of L-glutamic acid and pyruvic acid. Dr. Wolf's CHM 424 27- 89 Mechanism O – HO2CCH2CH2CHCO2 + CH3CCO2H + NH3 – HO2CCH2CH2CHCO2 N CH3CCO2– Dr. Wolf's CHM 424 27- 90 Formation of the imine is followed by proton removal at one carbon and protonation of another carbon. H – HO2CCH2CH2CCO2 N CH3CCO2– Dr. Wolf's CHM 424 27- 91 – HO2CCH2CH2CCO2 N – CH3CCO2 H H – HO2CCH2CH2CCO2 N CH3CCO2– Dr. Wolf's CHM 424 27- 92 – HO2CCH2CH2CCO2 N – CH3CCO2 H Hydrolysis of the imine function gives -keto glutarate and L-alanine. Dr. Wolf's CHM 424 27- 93 – HO2CCH2CH2CCO2 N – CH3CCO2 H H2O +NH3 – HO2CCH2CH2CCO2 O Dr. Wolf's CHM 424 + CH3CCO2– H 27- 94 Biosynthesis of L-Tyrosine L-Tyrosine is biosynthesized from L-phenylalanine. A key step is epoxidation of the aromatic ring to give an arene oxide intermediate. – CH2CHCO2 + NH3 Dr. Wolf's CHM 424 27- 95 Biosynthesis of L-Tyrosine – CH2CHCO2 O + NH3 O2, enzyme – CH2CHCO2 + NH3 Dr. Wolf's CHM 424 27- 96 Biosynthesis of L-Tyrosine – CH2CHCO2 O + NH3 enzyme HO – CH2CHCO2 + NH3 Dr. Wolf's CHM 424 27- 97 Biosynthesis of L-Tyrosine Conversion to L-tyrosine is one of the major metabolic pathways of L-phenylalanine. Individuals who lack the enzymes necessary to convert L-phenylalanine to L-tyrosine can suffer from PKU disease. In PKU disease, Lphenylalanine is diverted to a pathway leading to phenylpyruvic acid, which is toxic. Newborns are routinely tested for PKU disease. Treatment consists of reducing their dietary intake of phenylalanine-rich proteins. Dr. Wolf's CHM 424 27- 98 Decarboxylation Decarboxylation is a common reaction of amino acids. An example is the conversion of L-histidine to histamine. Antihistamines act by blocking the action of histamine. N – CH2CHCO2 N H Dr. Wolf's CHM 424 + NH3 27- 99 Decarboxylation N CH2CH2 NH2 N H –CO2, enzymes N – CH2CHCO2 N H Dr. Wolf's CHM 424 + NH3 27- 100 Neurotransmitters – The chemistry of the brain and central nervous system is affected by neurotransmitters. Several important neurotransmitters are biosynthesized from L-tyrosine. + H3N H CO2 H H OH L-Tyrosine Dr. Wolf's CHM 424 27- 101 Neurotransmitters – The common name of this compound is L-DOPA. It occurs naturally in the brain. It is widely prescribed to reduce the symptoms of Parkinsonism. + H3N H CO2 H H HO OH L-3,4-Dihydroxyphenylalanine Dr. Wolf's CHM 424 27- 102 Neurotransmitters Dopamine is formed by decarboxylation of L-DOPA. H H2N H H H HO OH Dopamine Dr. Wolf's CHM 424 27- 103 Neurotransmitters H H2N H H OH HO OH Norepinephrine Dr. Wolf's CHM 424 27- 104 Neurotransmitters H CH3NH H H OH HO OH Epinephrine Dr. Wolf's CHM 424 27- 105 Peptides Dr. Wolf's CHM 424 27- 106 Peptides Peptides are compounds in which an amide bond links the amino group of one -amino acid and the carboxyl group of another. An amide bond of this type is often referred to as a peptide bond. Dr. Wolf's CHM 424 27- 107 Peptide bond formation O Aspartate C H H C O H Alanine H H C C H C H + - H N H N H C O H C O H H O O + condensation Dr. Wolf's CHM 424 H2O 27- 108 Peptide bond formation O O C H H + C H C H C H N H C H O H H H C N H C O O Peptide bond Primary Structure Dr. Wolf's CHM 424 27- 109 Alanine and Glycine H + H3N C CH3 Dr. Wolf's CHM 424 H O C – O + H3N C O C – O H 27- 110 Alanylglycine H + H3N C CH3 H O C N C H H O C – O Two -amino acids are joined by a peptide bond in alanylglycine. It is a dipeptide. Dr. Wolf's CHM 424 27- 111 Alanylglycine H + H3N N-terminus C CH3 H O C N C H H O C – O C-terminus Ala—Gly AG Dr. Wolf's CHM 424 27- 112 Alanylglycine and glycylalanine are constitutional isomers H + H3N C C CH3 H + H3N C H Dr. Wolf's CHM 424 H O N C H H H O C N C H CH3 O C – O Alanylglycine Ala—Gly AG – O Glycylalanine Gly—Ala GA O C 27- 113 Alanylglycine H + H3N C CH3 H O C N C H H O C – O The peptide bond is characterized by a planar geometry. Dr. Wolf's CHM 424 27- 114 Higher Peptides Peptides are classified according to the number of amino acids linked together. dipeptides, tripeptides, tetrapeptides, etc. Leucine enkephalin is an example of a pentapeptide. Dr. Wolf's CHM 424 27- 115 Leucine Enkephalin Tyr—Gly—Gly—Phe—Leu YGGFL Dr. Wolf's CHM 424 27- 116 Oxytocin 3 2 4 5 Ile—Gln—Asn Tyr 1 Cys N-terminus C-terminus Cys—Pro—Leu—GlyNH2 6 S 7 8 9 S Oxytocin is a cyclic nonapeptide. Instead of having its amino acids linked in an extended chain, two cysteine residues are joined by an S—S bond. Dr. Wolf's CHM 424 27- 117 Oxytocin S—S bond An S—S bond between two cysteines is often referred to as a disulfide bridge. Dr. Wolf's CHM 424 27- 118 Introduction to Peptide Structure Determination Dr. Wolf's CHM 424 27- 119 Primary Structure The primary structure is the amino acid sequence plus any disulfide links. Dr. Wolf's CHM 424 27- 120 Classical Strategy (Sanger) 1. Determine what amino acids are present and their molar ratios. 2. Cleave the peptide into smaller fragments, and determine the amino acid composition of these smaller fragments. 3. Identify the N-terminus and C-terminus in the parent peptide and in each fragment. 4. Organize the information so that the sequences of small fragments can be overlapped to reveal the full sequence. Dr. Wolf's CHM 424 27- 121 Amino Acid Analysis Dr. Wolf's CHM 424 27- 122 Amino Acid Analysis Acid-hydrolysis of the peptide (6 M HCl, 24 hr) gives a mixture of amino acids. The mixture is separated by ion-exchange chromatography, which depends on the differences in pI among the various amino acids. Amino acids are detected using ninhydrin. Automated method; requires only 10-5 to 10-7 g of peptide. Dr. Wolf's CHM 424 27- 123 Partial Hydrolysis of Proteins Dr. Wolf's CHM 424 27- 124 Partial Hydrolysis of Peptides and Proteins Acid-hydrolysis of the peptide cleaves all of the peptide bonds. Cleaving some, but not all, of the peptide bonds gives smaller fragments. These smaller fragments are then separated and the amino acids present in each fragment determined. Enzyme-catalyzed cleavage is the preferred method for partial hydrolysis. Dr. Wolf's CHM 424 27- 125 Partial Hydrolysis of Peptides and Proteins The enzymes that catalyze the hydrolysis of peptide bonds are called peptidases, proteases, or proteolytic enzymes. Dr. Wolf's CHM 424 27- 126 Trypsin Trypsin is selective for cleaving the peptide bond to the carboxyl group of lysine or arginine. O O O NHCHC NHCHC NHCHC R R' R" lysine or arginine Dr. Wolf's CHM 424 27- 127 Chymotrypsin Chymotrypsin is selective for cleaving the peptide bond to the carboxyl group of amino acids with an aromatic side chain. O O O NHCHC NHCHC NHCHC R R' R" phenylalanine, tyrosine, tryptophan Dr. Wolf's CHM 424 27- 128 Carboxypeptidase Carboxypeptidase is selective for cleaving the peptide bond to the C-terminal amino acid. O O + H3NCHC R Dr. Wolf's CHM 424 protein C O – NHCHCO R 27- 129 End Group Analysis Dr. Wolf's CHM 424 27- 130 End Group Analysis Amino sequence is ambiguous unless we know whether to read it left-to-right or right-to-left. We need to know what the N-terminal and Cterminal amino acids are. The C-terminal amino acid can be determined by carboxypeptidase-catalyzed hydrolysis. Several chemical methods have been developed for identifying the N-terminus. They depend on the fact that the amino N at the terminus is more nucleophilic than any of the amide nitrogens. Dr. Wolf's CHM 424 27- 131 Sanger's Method The key reagent in Sanger's method for identifying the N-terminus is 1-fluoro-2,4dinitrobenzene. 1-Fluoro-2,4-dinitrobenzene is very reactive toward nucleophilic aromatic substitution (Section 23.5). NO2 O2N Dr. Wolf's CHM 424 F 27- 132 Sanger's Method 1-Fluoro-2,4-dinitrobenzene reacts with the amino nitrogen of the N-terminal amino acid. NO2 O2N O O F + H2NCHC NHCHC NHCH2C O2N Dr. Wolf's CHM 424 O NHCHC O NHCHC O NHCH2C CH(CH3)2 CH2C6H5 – NHCHCO CH3 CH(CH3)2 CH2C6H5 NO2 O O O – NHCHCO CH3 27- 133 Sanger's Method Acid hydrolysis cleaves all of the peptide bonds leaving a mixture of amino acids, only one of which (the N-terminus) bears a 2,4-DNP group. NO2 O O O O + + + NHCHCOH + H3NCHCO– + H3NCH2CO– + H3NCHCO– O2N CH(CH3)2 CH3 CH2C6H5 H3O+ NO2 O2N Dr. Wolf's CHM 424 O NHCHC O NHCHC O NHCH2C CH(CH3)2 CH2C6H5 O – NHCHCO CH3 27- 134 Insulin Dr. Wolf's CHM 424 27- 135 Insulin Insulin is a polypeptide with 51 amino acids. It has two chains, called the A chain (21 amino acids) and the B chain (30 amino acids). The following describes how the amino acid sequence of the B chain was determined. Dr. Wolf's CHM 424 27- 136 The B Chain of Bovine Insulin Phenylalanine (F) is the N terminus. Pepsin-catalyzed hydrolysis gave the four peptides: FVNQHLCGSHL VGAL VCGERGF YTPKA Dr. Wolf's CHM 424 27- 137 The B Chain of Bovine Insulin FVNQHLCGSHL VGAL VCGERGF YTPKA Dr. Wolf's CHM 424 27- 138 The B Chain of Bovine Insulin Phenylalanine (F) is the N terminus. Pepsin-catalyzed hydrolysis gave the four peptides: FVNQHLCGSHL VGAL VCGERGF YTPKA Overlaps between the above peptide sequences were found in four additional peptides: SHLV LVGA ALT TLVC Dr. Wolf's CHM 424 27- 139 The B Chain of Bovine Insulin FVNQHLCGSHL SHLV LVGA VGAL ALT TLVC VCGERGF YTPKA Dr. Wolf's CHM 424 27- 140 The B Chain of Bovine Insulin Phenylalanine (F) is the N terminus. Pepsin-catalyzed hydrolysis gave the four peptides: FVNQHLCGSHL VGAL VCGERGF YTPKA Overlaps between the above peptide sequences were found in four additional peptides: SHLV LVGA ALT TLVC Trypsin-catalyzed hydrolysis gave GFFYTPK which completes the sequence. Dr. Wolf's CHM 424 27- 141 The B Chain of Bovine Insulin FVNQHLCGSHL SHLV LVGA VGAL ALT TLVC VCGERGF GFFYTPK YTPKA Dr. Wolf's CHM 424 27- 142 The B Chain of Bovine Insulin FVNQHLCGSHL SHLV LVGA VGAL ALT TLVC VCGERGF GFFYTPK YTPKA FVNQHLCGSHLVGALTLVCGERGFFYTPKA Dr. Wolf's CHM 424 27- 143 Insulin The sequence of the A chain was determined using the same strategy. Establishing the disulfide links between cysteine residues completed the primary structure. Dr. Wolf's CHM 424 27- 144 Primary Structure of Bovine Insulin N terminus of A chain S S C terminus of A chain 15 5 E Q C V C S L Y Q L I F E N 20 C V YC A S S 10 N S S H L N Q V S C F G S H L V G A L Y L V 5 C 15 G 20 10 N terminus E of B chain G R F F Y K P T A C terminus 25 30 of B chain Dr. Wolf's CHM 424 27- 145 The Edman Degradation and Automated Sequencing of Peptides Dr. Wolf's CHM 424 27- 146 Edman Degradation 1. Method for determining N-terminal amino acid. 2. Can be done sequentially one residue at a time on the same sample. Usually one can determine the first 20 or so amino acids from the N-terminus by this method. 3. 10-10 g of sample is sufficient. 4. Has been automated. Dr. Wolf's CHM 424 27- 147 Edman Degradation The key reagent in the Edman degradation is phenyl isothiocyanate. N Dr. Wolf's CHM 424 C S 27- 148 Edman Degradation Phenyl isothiocyanate reacts with the amino nitrogen of the N-terminal amino acid. O C6H5N Dr. Wolf's CHM 424 C S + + H3NCHC R NH peptide 27- 149 Edman Degradation S O C6H5NHCNHCHC peptide NH R O C6H5N Dr. Wolf's CHM 424 C S + + H3NCHC R NH peptide 27- 150 Edman Degradation S O C6H5NHCNHCHC NH peptide R The product is a phenylthiocarbamoyl (PTC) derivative. The PTC derivative is then treated with HCl in an anhydrous solvent. The N-terminal amino acid is cleaved from the remainder of the peptide. Dr. Wolf's CHM 424 27- 151 Edman Degradation S O C6H5NHCNHCHC peptide NH R HCl S C6H5NH C C N Dr. Wolf's CHM 424 CH R O + + H3N peptide 27- 152 Edman Degradation The product is a thiazolone. Under the conditions of its formation, the thiazolone rearranges to a phenylthiohydantoin (PTH) derivative. S C6H5NH C C N Dr. Wolf's CHM 424 CH R O + + H3N peptide 27- 153 Edman Degradation C6H5 S N C C The PTH derivative is isolated and identified. The remainder of the peptide is subjected to a second Edman degradation. O CH HN R S C6H5NH C C N Dr. Wolf's CHM 424 CH R O + + H3N peptide 27- 154 The Strategy of Peptide Synthesis Dr. Wolf's CHM 424 27- 155 General Considerations Making peptide bonds between amino acids is not difficult. The challenge is connecting amino acids in the correct sequence. Random peptide bond formation in a mixture of phenylalanine and glycine, for example, will give four dipeptides. Phe—Phe Gly—Gly Phe—Gly Gly—Phe Dr. Wolf's CHM 424 27- 156 General Strategy 1. Limit the number of possibilities by "protecting" the nitrogen of one amino acid and the carboxyl group of the other. N-Protected phenylalanine O X NHCHCOH Dr. Wolf's CHM 424 CH2C6H5 C-Protected glycine O H2NCH2C Y 27- 157 General Strategy 2. Couple the two protected amino acids. X O O NHCHC NHCH2C Y CH2C6H5 O X NHCHCOH Dr. Wolf's CHM 424 CH2C6H5 O H2NCH2C Y 27- 158 General Strategy 3. Deprotect the amino group at the N-terminus and the carboxyl group at the C-terminus. X O O NHCHC NHCH2C Y CH2C6H5 O + H3NCHC Dr. Wolf's CHM 424 O – NHCH2CO CH2C6H5 Phe-Gly 27- 159 Amino Group Protection Dr. Wolf's CHM 424 27- 160 Protect Amino Groups as Amides Amino groups are normally protected by converting them to amides. Benzyloxycarbonyl (C6H5CH2O—) is a common protecting group. It is abbreviated as Z. Z-protection is carried out by treating an amino acid with benzyloxycarbonyl chloride. Dr. Wolf's CHM 424 27- 161 Protect Amino Groups as Amides O O CH2OCCl + + – H3NCHCO CH2C6H5 1. NaOH, H2O 2. H+ O CH2OC O NHCHCOH CH2C6H5 Dr. Wolf's CHM 424 (82-87%) 27- 162 Protect Amino Groups as Amides O CH2OC O NHCHCOH CH2C6H5 is abbreviated as: O ZNHCHCOH or Z-Phe CH2C6H5 Dr. Wolf's CHM 424 27- 163 Removing Z-Protection An advantage of the benzyloxycarbonyl protecting group is that it is easily removed by: a) hydrogenolysis b) cleavage with HBr in acetic acid Dr. Wolf's CHM 424 27- 164 Hydrogenolysis of Z-Protecting Group O CH2OC O NHCHCNHCH2CO2CH2CH3 CH2C6H5 H2, Pd O CH3 CO2 H2NCHCNHCH2CO2CH2CH3 CH2C6H5 Dr. Wolf's CHM 424 (100%) 27- 165 HBr Cleavage of Z-Protecting Group O CH2OC O NHCHCNHCH2CO2CH2CH3 CH2C6H5 HBr O CH2Br Dr. Wolf's CHM 424 CO2 + H3NCHCNHCH2CO2CH2CH3 – CH2C6H5 Br (82%) 27- 166 The tert-Butoxycarbonyl Protecting Group O (CH3)3COC O NHCHCOH CH2C6H5 is abbreviated as: O BocNHCHCOH or Boc-Phe CH2C6H5 Dr. Wolf's CHM 424 27- 167 HBr Cleavage of Boc-Protecting Group O (CH3)3COC O NHCHCNHCH2CO2CH2CH3 CH2C6H5 HBr O H3C C H3C Dr. Wolf's CHM 424 CH2 CO2 + H3NCHCNHCH2CO2CH2CH3 – CH2C6H5 Br (86%) 27- 168 Carboxyl Group Protection Dr. Wolf's CHM 424 27- 169 Protect Carboxyl Groups as Esters Carboxyl groups are normally protected as esters. Deprotection of methyl and ethyl esters is by hydrolysis in base. Benzyl esters can be cleaved by hydrogenolysis. Dr. Wolf's CHM 424 27- 170 Hydrogenolysis of Benzyl Esters O O C6H5CH2OC O NHCHCNHCH2COCH2C6H5 CH2C6H5 H2, Pd O C6H5CH3 Dr. Wolf's CHM 424 CO2 + – H3NCHCNHCH2CO CH2C6H5 (87%) CH3C6H5 27- 171 Peptide Bond Formation Dr. Wolf's CHM 424 27- 172 aa are covalently linked by amide bonds (Peptide Bonds) The resulting molecules are called Peptides & Proteins R' R' N C O R N C R O Features of a Peptide Bond; 1. Usually inert 2. Planar to allow delocalisation 3. Restricted Rotation about the amide bond 4. Rotation of Groups (R and R’) attached to the amide bond is relatively free Dr. Wolf's CHM 424 27- 173 aa that are part of a peptide or protein are referred to as residues. Peptides are made up of about 50 residues, and do not possess a well-defined 3D-structure Proteins are larger molecules that usually contain at least 50 residues, and sometimes 1000. The most important feature of proteins is that they possess well-defined 3D-structure. Primary Structure is the order (or sequence) of amino acid residues Peptides are always written and named with the amino terminus on the left and the carboxy terminus on the right Dr. Wolf's CHM 424 27- 174 CH2OH CH3 O O H3 N CH H3 N C O C H3 N C O O O Serine Alanine Valine - 2 H2 O CH3 O H N H3 N C C O CH2OH O N H C O Tripeptide : Ala . Ser. Val Strong Acid Required to hydrolyse peptide bonds Dr. Wolf's CHM 424 27- 175 Lys. Cys. Phe 1. RSH Phe. Ser. Cys 2. 6 M HCl hydrolysis Lys + 2 Cys + 2 Phe + Ser Ph Cysteine residues create Disulfide Bridges between chains (CH2)4NH2 O H N H2 N C C OH N H O O This does not reveal Primary Structure C S S Ph O H N H2 N C OH N H O C O HO Dr. Wolf's CHM 424 27- 176 Forming Peptide Bonds The two major methods are: 1. coupling of suitably protected amino acids using N,N'-dicyclohexylcarbodiimide (DCCI) 2. via an active ester of the N-terminal amino acid. Dr. Wolf's CHM 424 27- 177 DCCI-Promoted Coupling O O ZNHCHCOH + H2NCH2COCH2CH3 CH2C6H5 DCCI, chloroform O ZNHCHC O NHCH2COCH2CH3 CH2C6H5 Dr. Wolf's CHM 424 (83%) 27- 178 Mechanism of DCCI-Promoted Coupling O + ZNHCHCOH C6H11N C NC6H11 CH2C6H5 H C6H11N O C C6H11N Dr. Wolf's CHM 424 OCCHNHZ CH2C6H5 27- 179 Mechanism of DCCI-Promoted Coupling The species formed by addition of the Zprotected amino acid to DCCI is similar in structure to an acid anhydride and acts as an acylating agent. Attack by the amine function of the carboxylprotected amino acid on the carbonyl group leads to nucleophilic acyl substitution. H C6H11N O C C6H11N Dr. Wolf's CHM 424 OCCHNHZ CH2C6H5 27- 180 Mechanism of DCCI-Promoted Coupling O H C6H11N C O + ZNHCHC O NHCH2COCH2CH3 CH2C6H5 C6H11NH O H2NCH2COCH2CH3 H C6H11N O C C6H11N Dr. Wolf's CHM 424 OCCHNHZ CH2C6H5 27- 181 The Active Ester Method A p-nitrophenyl ester is an example of an "active ester." p-Nitrophenyl is a better leaving group than methyl or ethyl, and p-nitrophenyl esters are more reactive in nucleophilic acyl substitution. Dr. Wolf's CHM 424 27- 182 The Active Ester Method O O ZNHCHCO NO2 + H2NCH2COCH2CH3 CH2C6H5 chloroform O ZNHCHC O NHCH2COCH2CH3 + HO CH2C6H5 Dr. Wolf's CHM 424 NO2 (78%) 27- 183 Solid-Phase Peptide Synthesis: The Merrifield Method Dr. Wolf's CHM 424 27- 184 Solid-Phase Peptide Synthesis In solid-phase synthesis, the starting material is bonded to an inert solid support. Reactants are added in solution. Reaction occurs at the interface between the solid and the solution. Because the starting material is bonded to the solid, any product from the starting material remains bonded as well. Purification involves simply washing the byproducts from the solid support. Dr. Wolf's CHM 424 27- 185 The Solid Support CH2 CH CH2 CH CH2 CH CH2 CH The solid support is a copolymer of styrene and divinylbenzene. It is represented above as if it were polystyrene. Cross-linking with divinylbenzene simply provides a more rigid polymer. Dr. Wolf's CHM 424 27- 186 The Solid Support CH2 CH CH2 CH CH2 CH CH2 CH Treating the polymeric support with chloromethyl methyl ether (ClCH2OCH3) and SnCl4 places ClCH2 side chains on some of the benzene rings. Dr. Wolf's CHM 424 27- 187 The Solid Support CH2 CH CH2 CH CH2 CH CH2 CH CH2Cl The side chain chloromethyl group is a benzylic halide, reactive toward nucleophilic substitution (SN2). Dr. Wolf's CHM 424 27- 188 The Solid Support CH2 CH CH2 CH CH2 CH CH2 CH CH2Cl The chloromethylated resin is treated with the Bocprotected C-terminal amino acid. Nucleophilic substitution occurs, and the Boc-protected amino acid is bound to the resin as an ester. Dr. Wolf's CHM 424 27- 189 The Merrifield Procedure CH2 CH CH2 CH CH2 O – BocNHCHCO CH CH2 CH CH2Cl R Dr. Wolf's CHM 424 27- 190 The Merrifield Procedure CH2 CH CH2 CH CH2 O CH CH2 CH CH2 BocNHCHCO Next, the Boc protecting group is removed with HCl. Dr. Wolf's CHM 424 R 27- 191 The Merrifield Procedure CH2 CH CH2 CH CH2 O CH CH2 CH CH2 H2NCHCO DCCI-promoted coupling adds the second amino acid Dr. Wolf's CHM 424 R 27- 192 The Merrifield Procedure CH2 CH CH2 CH CH2 O BocNHCHC R' CH O CH2 CH CH2 NHCHCO R Remove the Boc protecting group. Dr. Wolf's CHM 424 27- 193 The Merrifield Procedure CH2 CH CH2 CH CH2 O H2NCHC R' CH O CH2 CH CH2 NHCHCO R Add the next amino acid and repeat. Dr. Wolf's CHM 424 27- 194 The Merrifield Procedure CH2 CH CH2 O CH O + H3N peptide C NHCHC R' Dr. Wolf's CHM 424 CH2 CH O CH2 CH CH2 NHCHCO R Remove the peptide from the resin with HBr in CF3CO2H 27- 195 The Merrifield Procedure CH2 CH CH2 CH CH CH2 CH CH2Br O O + H3N peptide C NHCHC R' Dr. Wolf's CHM 424 CH2 O – NHCHCO R 27- 196 The Merrifield Method Merrifield automated his solid-phase method. Synthesized a nonapeptide (bradykinin) in 1962 in 8 days in 68% yield. Synthesized ribonuclease (124 amino acids) in 1969. 369 reactions; 11,391 steps Nobel Prize in chemistry: 1984 Dr. Wolf's CHM 424 27- 197 Secondary Structures of Peptides and Proteins Dr. Wolf's CHM 424 27- 198 Levels of Protein Structure Primary structure = the amino acid sequence plus disulfide links Secondary structure = conformational relationship between nearest neighbor amino acids helix pleated sheet Dr. Wolf's CHM 424 27- 199 Levels of Protein Structure The -helix and pleated sheet are both characterized by: planar geometry of peptide bond anti conformation of main chain hydrogen bonds between N—H and O=C Dr. Wolf's CHM 424 27- 200 -helixes Intra-chain H-bonds Secondary Structure Dr. Wolf's CHM 424 27- 201 -strands Inter-chain H-bonds Secondary Structure Dr. Wolf's CHM 424 27- 202 Pleated Sheet Shown is a sheet of protein chains composed of alternating glycine and alanine residues. Adjacent chains are antiparallel. Hydrogen bonds between chains. van der Waals forces produce pleated effect. Dr. Wolf's CHM 424 27- 203 Pleated Sheet Sheet is most commonly seen with amino acids having small side chains (glycine, alanine, serine). 80% of fibroin (main protein in silk) is repeating sequence of —Gly—Ser—Gly—Ala—Gly—Ala—. Sheet is flexible, but resists stretching. Dr. Wolf's CHM 424 27- 204 Helix Shown is an helix of a protein in which all of the amino acids are L-alanine. Helix is right-handed with 3.6 amino acids per turn. Hydrogen bonds are within a single chain. Protein of muscle (myosin) and wool (-keratin) contain large regions of -helix. Chain can be stretched. Dr. Wolf's CHM 424 27- 205 Secondary Structure The Development of Regular patterns of Hydrogen Bonding, which result in distinct folding patterns -helix Dr. Wolf's CHM 424 -pleated sheets 27- 206 Tertiary Structure of Peptides and Proteins Dr. Wolf's CHM 424 27- 207 Tertiary Structure Refers to overall shape (how the chain is folded) Fibrous proteins (hair, tendons, wool) have elongated shapes Globular proteins are approximately spherical most enzymes are globular proteins an example is carboxypeptidase Dr. Wolf's CHM 424 27- 208 Tertiary Structure This is the 3D structure resulting from further regular folding of the polypeptide chains using H-bonding, Van der Waals, disulfide bonds and electrostatic forces – Often detected by X-ray crystallographic methods Globular Proteins – “Spherical Shape” , include Insulin, Hemoglobin, Enzymes, Antibodies ---polar hydrophilic groups are aimed outwards towards water, whereas non-polar “greasy” hydrophobic hydrocarbon portions cluster inside the molecule, so protecting them from the hostile aqueous environment ----- Soluble Proteins Fibrous Proteins – “Long thin fibres” , include Hair, wool, skin, nails – less folded ----- e.g. keratin - the -helix strands are wound into a “superhelix”. The superhelix makes one complete turn for each 35 turns of the -helix. Dr. Wolf's CHM 424 27- 209 In globular proteins this tertiary structure or macromolecular shape determines biological properties Bays or pockets in proteins are called Active Sites Enzymes are Stereospecific and possess Geometric Specificity The range of compounds that an enzyme excepts varies from a particular functional group to a specific compound Emil Fischer formulated the lock-and-key mechanism for enzymes All reactions which occur in living cells are mediated by enzymes and are catalysed by 106-108 Some enzymes may require the presence of a Cofactor. This may be a metal atom, which is essential for its redox activity. Others may require the presence of an organic molecule, such as NAD+, called a Coenzyme. If the Cofactor is permanently bound to the enzyme, it is called a Prosthetic Group. Dr. Wolf's CHM 424 27- 210 For a protein composed of a single polypeptide molecule, tertiary structure is the highest level of structure that is attained Myoglobin and hemoglobin were the first proteins to be successfully subjected to completely successful X-rays analysis by J. C. Kendrew and Max Perutz (Nobel Prize for Chemistry 1962) Quaternary Structure When multiple sub-units are held together in aggregates by Van der Waals and electrostatic forces (not covalent bonds) Hemoglobin is tetrameric myglobin For example, Hemoglobin has four heme units, the protein globin surrounds the heme – Takes the shape of a giant tetrahedron – Two identical and globins. The and chains are very similar but distinguishable in both primary structure and folding Dr. Wolf's CHM 424 27- 211 Tertiary structure Hb monomer (or myoglobin) Dr. Wolf's CHM 424 Quaternary structure Hb 22 tetramer 27- 212 Carboxypeptidase Carboxypeptidase is an enzyme that catalyzes the hydrolysis of proteins at their C-terminus. It is a metalloenzyme containing Zn2+ at its active site. An amino acid with a positively charged side chain (Arg-145) is near the active site. Dr. Wolf's CHM 424 27- 213 Carboxypeptidase Disulfide bond Zn2+ Arg-145 N-terminus C-terminus tube model Dr. Wolf's CHM 424 ribbon model 27- 214 What happens at the active site? •• • O• + H3N peptide C O NHCHC – R Dr. Wolf's CHM 424 O H2N + C Arg-145 H2N 27- 215 What happens at the active site? •• • O• + H3N peptide C O NHCHC – R O H2N + C Arg-145 H2N The peptide or protein is bound at the active site by electrostatic attraction between its negatively charged carboxylate ion and arginine-145. Dr. Wolf's CHM 424 27- 216 What happens at the active site? •• • O• + H3N peptide C Zn2+ O NHCHC – R O H2N + C Arg-145 H2N Zn2+ acts as a Lewis acid toward the carbonyl oxygen, increasing the positive character of the carbonyl carbon. Dr. Wolf's CHM 424 27- 217 What happens at the active site? Zn2+ •• • O• + H3N peptide C O NHCHC – R O H2N + C Arg-145 H2N H • O• • • H Water attacks the carbonyl carbon. Nucleophilic acyl substitution occurs. Dr. Wolf's CHM 424 27- 218 What happens at the active site? Zn2+ H2N + C •• • O• + H3N peptide C •• – O •• •• H2N O + H3NCHC – R Dr. Wolf's CHM 424 Arg-145 O 27- 219 Coenzymes Dr. Wolf's CHM 424 27- 220 Coenzymes The range of chemical reactions that amino acid side chains can participate in is relatively limited. acid-base (transfer and accept protons) nucleophilic acyl substitution Many other biological processes, such as oxidation-reduction, require coenzymes, cofactors, or prosthetic groups in order to occur. Dr. Wolf's CHM 424 27- 221 Coenzymes NADH, coenzyme A and coenzyme B12 are examples of coenzymes. Heme is another example. Dr. Wolf's CHM 424 27- 222 Heme H 2C CH H 3C CH3 N N CH CH2 Fe N H 3C HO2CCH2CH2 N CH3 CH2CH2CO2H Molecule surrounding iron is a type of porphyrin. Dr. Wolf's CHM 424 27- 223 Myoglobin C-terminus Heme N-terminus Heme is the coenzyme that binds oxygen in myoglobin (oxygen storage in muscles) and hemoglobin (oxygen transport). Dr. Wolf's CHM 424 27- 224 Protein Quaternary Structure: Hemoglobin Dr. Wolf's CHM 424 27- 225 Protein Quaternary Structure Some proteins are assemblies of two or more chains. The way in which these chains are organized is called the quaternary structure. Hemoglobin, for example, consists of 4 subunits. There are 2 chains (identical) and 2 chains (also identical). Each subunit contains one heme and each protein is about the size of myoglobin. Dr. Wolf's CHM 424 27- 226 Protein Structure Primary structure is the amino acid sequence. Secondary structure is how the amino acids in sequence fold up locally. Examples are -helixes and -strands and loops. Tertiary structure is the 3-dimensional folding of the secondary structural elements and connecting loops in space. Quaternary structure is the association of multiple subunits, each with a tertiary structure and each a unique gene product. Dr. Wolf's CHM 424 27- 227 Stabilization of Protein Structure Electrostatic interactions involve the interaction of (+) and (-) charged side groups. Hydrogen bonds involve sharing of a hydrogen atom between two eletronegative atoms (e.g., O, N). Van der Waal’s forces are weak forces based on optimal overlap of adjacent electronic orbitals. Can be repulsive. Hydrophobic interactions are, by far, the most powerful force stabilizing protein structure. Basis of force is entropy gain realized by burying hydrophobic residues. Dr. Wolf's CHM 424 27- 228 Cofactors Cofactors are exogenous molecules that associate with proteins to yield full activity. In the absence of cofactor, protein is an apoprotein. Co-enzymes are soluble and associate transiently with enzyme during catalytic cycle. An example is vitamin K in activation of blood clotting enzymes. Prosthetic groups are covalently attached to the protein. Examples are heme, in hemoglobin, and riboflavin, in flavoproteins. Dr. Wolf's CHM 424 27- 229 REVERSIBLE DENATURING Oxidation RS SR RS H Reduction Prof. Linus Pauling Dr. Frederick Sanger, Prof. R. B. Merrifield Nobel Prize for Chemistry Nobel Prize for Chemistry 1984 1958 and 1980 Automated Peptide Synthesis Dr. Wolf's CHM 424 Peptide sequencing 27- 230 Globular Proteins Globular proteins fold so as to "bury" the hydrophobic side chains, minimizing their contact with water Most polar residues face the outside of the protein and interact with solvent Most hydrophobic residues face the interior of the protein and interact with each other Packing of residues is close, but protein interiors contain some empty space The empty space is in the form of small cavities Dr. Wolf's CHM 424 27- 231 Globular Proteins Globular proteins fold so as to "bury" the hydrophobic side chains, minimizing their contact with water Most polar residues face the outside of the protein and interact with solvent Most hydrophobic residues face the interior of the protein and interact with each other Packing of residues is close, but protein interiors contain some empty space The empty space is in the form of small cavities Dr. Wolf's CHM 424 27- 232 Objectives Diagram and describe the effect of oxygen on the position of iron relative to the heme plane. Describe how cooperative binding of oxygen by hemoglobin improves its effectiveness as an oxygen carrier. Describe the relationship between Hb structure to the Bohr effect and explain its physiological significance.. Discuss how carbon dioxide affects the affinity of Hb for oxygen and why this is physiologically significant. Explain the effect of bisphosphoglycerate (BPG) on the affinity of Hb for oxygen and how this is related to altitude and HbF. Explain how carbon monoxide (CO) binds to Hb and its affinity relative to that of oxygen.. Describe the molecular basis of thalassemias and the aberrant Hb that are produced in these diseases.. List three embryonic forms of Hb.. Dr. Wolf's CHM 424 27- 233 Myoglobin: 2o and 3o aspects Myoglobin is a single peptide chain of 153 residues arranged in eight -helical regions labeled A-H. The heme cofactor is the oxygen binding site so it is necessary for myoglobin’s function, oxygen storage in mammalian muscle tissue. His E7 and F8 are important for binding the heme group within the protein and for stabilizing bound oxygen. Dr. Wolf's CHM 424 27- 234 Myoglobin and Hemoglobin Mb is monomer, Hb is a tetramer (22). Hb subunits are structurally similar to Mb, with 8 helical regions, no -strands and no water. Both contain heme prosthetic group Both Mb and Hb contain proximal and distal histidines. Affinity of Mb for oxygen is high, affinity of Hb for oxygen is low. Dr. Wolf's CHM 424 27- 235 Myoglobin &Hemoglobin Two related protein for O2 transportation. Mb has one chain Hb has four chains Each chain has two parts: a globin ( protein) and a heme ( non-protein) Dr. Wolf's CHM 424 27- 236 Myoglobin • An O2 transport protein in muscle • A Globin( globular soluble protein), 151 residues that contains 8 -helices (A,B,C,…..H) •Contains a heme •prosthetic group Binds heme in hydrophobic pocket. Polar groups exposed to solvent, Non-polar groups buried. Dr. Wolf's CHM 424 27- 237 Myoglobin: Dr. Wolf's CHM 424 o 2 and o 3 structure 27- 238 The Heme Prosthetic Group • Protoporphyrin with Fe(II) • Covalent attachment of Fe via His F8 side chain • Additional stabilization via hydrophobic interaction • Fe(II) state is active, Fe(III) [oxidized] • Fe(II) atom in heme binds O2 Dr. Wolf's CHM 424 27- 239 Binding of O2 to Heme • Binding of O2 to a free heme group is irreversible ( heme- heme sandwich) • Enclosure in a protein( globin) allows reversible binding O2 has only limited solubility (1 X 10-4 M) in water Solubility problem overcome by binding to proteins • Binding of O2 alters heme structure Bright scarlet color of blood in arteries Dark purple color of blood in veins Dr. Wolf's CHM 424 27- 240 The Heme Group - CH2CH2COO OOC CH2CH2 H3C N - N CH3 Fe(II) H2C N CH CH3 CH3 Dr. Wolf's CHM 424 N Pyrrole ring CH CH2 27- 241 N of His F8 binds to 5th coordination site on heme iron Oxygen binds to 6th coordination site on heme iron Dr. Wolf's CHM 424 27- 242 His E7 acts as a gate to favor oxygen binding over carbon monoxide. Dr. Wolf's CHM 424 27- 243 Hemoglobin A tetrameric protein two -chains (141 AA) two -chains (146 AA) four heme cofactors, one in each chain The and chains are homologous to myoglobin. Oxygen binds to heme in hemoglobin with same structure as in Mb but cooperatively: as one O2 is bound, it becomes easier for the next to bind. Dr. Wolf's CHM 424 27- 244 Hemoglobin • Ubiquitous O2 transport protein • A globular soluble protein, 2X2 chains (164 kDa) • and chains 44% identical • All helical secondary structure (like myoglobin) • quaternary structure -subunit 141 residues -subunit 146 residues • Extensive contacts between subunits Mix of hydrophobic, H-bond, and ionic interactions 11 (22)- 35 residues, 12 (21)- 19 residues Dr. Wolf's CHM 424 27- 245 Dr. Wolf's CHM 424 27- 246 Each chain is in ribbon form. The heme groups are in space filling form Dr. Wolf's CHM 424 27- 247 Oxygen Binding Curves Hemoglobin and myoglobin respond differently to increase in O2 concentration. Myoglobin shows normal saturation behavior while hemoglobin shows cooperative behavior. Each oxygen added to a heme of Hb makes addition of the next one easier. The myoglobin curve is hyperbolic. The hemoglobin curve is sigmoidal. Dr. Wolf's CHM 424 27- 248 Hemoglobin O2 Binding Curve Binding curve is sigmoidal Artery: high pO2, loading of protein Vein: lower pO2, unloading from protein P50(hemoglobin) = 26 torr, adjusts as needed!! *Drastic change in pO2 over physiological range* Dr. Wolf's CHM 424 27- 249 Oxygen Binding Curves-2 Dr. Wolf's CHM 424 27- 250 Hemoglobin Equilibrium O2 + H ,CO 2, BPG T R (low affinity) (high affinity) Dr. Wolf's CHM 424 27- 251 A Quaternary Structure Change One alpha-beta pair moves relative to the other by 15 degrees upon oxygen binding This large change is caused by movement of Fe by only 0.039 nm when oxygen binds Dr. Wolf's CHM 424 27- 252 Oxygen binding by hemoglobin Dr. Wolf's CHM 424 27- 253 Allosteric Effectors • The R or T state can be stabilized by the binding of ligands other than O2. 1. H+. Lower pH favors the T state which causes Hb to release bound O2. This is known as the Bohr Effect. 2. CO2. Release of CO2 lowers pH via conversion to HCO3-: CO2 + H2O HCO3- + H+. Reinforces Bohr Effect 3. Bisphosphoglycerate (BPG). Regulation of activity via binding more strongly to T state, helps to release O2. Increase in levels of BPG helps adaptation to high altitude- faster than making more hemoglobin. Also important in hypoxia diseases (e.g. anemia) Dr. Wolf's CHM 424 27- 254 The Bohr Effect Competition between oxygen and H+ Discovered by Christian Bohr Binding of protons diminishes oxygen binding Binding of oxygen diminishes proton binding Important physiological significance-O2 saturation of Hb responds to pH Dr. Wolf's CHM 424 27- 255 The Bohr Effect Dr. Wolf's CHM 424 27- 256 Bohr Effect II Carbon dioxide diminishes oxygen binding CO2 produced in metabolically active tissue (requires oxygen) Hydration of CO2 in tissues and extremities leads to proton production CO2 + H2O HCO3- + H+ These protons are taken up by Hb forcing more oxygen to dissociate The reverse occurs in the lungs Dr. Wolf's CHM 424 27- 257 Carbon Monoxide Poisoning • Heme Fe(II) binds many other small molecules with structures similar to O2 including: CO, NO, H2S • O2 is actually binds to these other molecules, particularly CO. • When exposed to CO, even at low concentrations, O2 transport proteins will be filled with CO limiting their vital O2 capacity. Dr. Wolf's CHM 424 27- 258 2,3-Bisphosphoglycerate An Allosteric Effector of Hemoglobin The sigmoid binding curve is only observed in the presence of 2,3-BPG Since 2,3-BPG binds at a site distant from the Fe where oxygen binds, it is called an allosteric effector Dr. Wolf's CHM 424 27- 259 2,3-bisphosphoglycerate (2,3-BPG) is a negative allosteric effector of O2 binding to Hb - binds tighter to deoxyHb 2,3-BPG Dr. Wolf's CHM 424 27- 260 Heme in hemoglobin Proximal His (F8) Proximal His (F8) Proximal His (F8) =C =N =O =C =N =O Fe Proximal His (F8) Distal His (E7) Heme prosthetic group Dr. Wolf's CHM 424 =C =N =O Fe Distal His (E7) Distal His (E7) Proximal His (F8) Fe =C =N =O Fe Distal His (E7) Fe =C =N =O Distal His (E7) Side view of Hb tetramer 27- 261 Binding of oxygen to heme iron Ferrous is reduced and +2 charge Proximal His (F8) Ferric is oxidized and +3 charge Fe - e =C =N =O Dr. Wolf's CHM 424 Distal His (E7) 27- 262 Effect of oxygen on heme iron =C =N =O FG1 C N F7 FG3 FG2 F6 Proximal His (F8) Fe Plane of heme D istal H is(E 7) Dr. Wolf's CHM 424 27- 263 Effect of oxygen on heme iron =C =N =O N FG1 C FG3 F7 FG2 F6 Proximal His (F8) Plane of heme Fe D istal H is(E 7) Dr. Wolf's CHM 424 27- 264 Effect of oxygen on heme iron =C =N =O FG1 FG1 CC FG3FG3 N N F7 F7 FG2 FG2 F6F6 P roximal P roximal H is 8) H is(F (F 8) Fe Plane of heme Fe D istal H is(E 7) Dr. Wolf's CHM 424 27- 265 Cooperativity Oxygen binding to one subunit of Hb, increases the affinity of the other subunits for additional oxygens. In other words, the first one is the hardest, the rest are easy. Example: square of postage stamps. Book of second four stamps. To To pull stamp, To pull third stamp, you To pull fourth stamp, pull first stamp, youonly you have to break have to break only one you don’t have to break have to break two edges. one edge. edge. any edges. Dr. Wolf's CHM 424 27- 266 Cooperativity 100 BPGEffect Mbalone Hb 80 Hb +Hb BPG 60 40 Sigmoid shape indicates positive cooperativity 20 pO2 vs p50=8 pO2 vs p50=26 0 0 20 40 60 80 100 120 140 160 pO2 (mm Hg) Dr. Wolf's CHM 424 27- 267 Bohr Effect Bohr Effect 100 7.4 7.6 80 7.0 7.2 60 O2 level in arterial blood 40 20 0 O2 level in venous blood Dr. Wolf's CHM 424 0 20 40 60 80 100 120 140 160 pO2 (mm Hg) 27- 268 Hb structural families Alpha family 1,2 - found in adult hemoglobins HbA1, HbA2. z - found in embryonic hemoglobins Hb Gower 1 and Hb Portland. Beta family - found in adult hemoglobin HbA1. d - found in adult hemoglobin HbA2. - found in fetal hemoglobin HbF. e - found in embryonic hemoglobin Hb Gower 1 and Hb Gower 2 Dr. Wolf's CHM 424 27- 269 CO2 effect CO2 Effect 100 pCO2 20 mm 80 pCO2 80 mm 60 40 20 pO2 vs p50=20 pO2 vs p50=40 0 0 20 40 60 80 100 120 140 160 pO2 (mm Hg) Dr. Wolf's CHM 424 27- 270 Effect of BPG BPG is responsible for cooperativity. BPGEffect Hb alone 100 80 Hb + BPG 60 40 High altitude increases BPG, pushing curve further to right 20 pO2 vs p50=8 pO2 vs p50=26 0 0 20 40 60 80 100 120 140 160 pO2 (mm Hg) Dr. Wolf's CHM 424 27- 271 Effect of BPG BPG Side view (R) Dr. Wolf's CHM 424 Side view (T) 27- 272 Effect of BPG - Lys His His + + + Dr. Wolf's CHM 424 27- 273 Hemoglobin Equilibrium O2 + H ,CO 2, BPG T R (low affinity) (high affinity) Dr. Wolf's CHM 424 27- 274 Hemoglobins in normal adults α β α γ α β α γ α δ δ α HbA HbF HbA2 98% ~1% <3.5% Dr. Wolf's CHM 424 27- 275 Dr. Wolf's CHM 424 27- 276 Globin gene clusters cluster, 16p 13.3 z 2 1 1,2= duplicate genes, both expressed cluster, 11p 15.5 e G A d G,=fetal genes, Gly and Ala at postion 136, Dr. Wolf's CHM 424 both expressed 27- 277 Hb structural families Alpha family 1,2 - found in adult hemoglobins HbA1, HbA2. z - found in embryonic hemoglobins Hb Gower 1 and Hb Portland. - (theta) newly discovered embryonic form. Beta family - found in adult hemoglobin HbA1. d - found in adult hemoglobin HbA2. - found in fetal hemoglobin HbF. e - found in embryonic hemoglobin Hb Gower 1 and Hb Gower 2 Dr. Wolf's CHM 424 27- 278 FETAL AND NEONATAL ERYTHROPOIESIS TABLE 1. Globin-chain development and composition Developmental stage Hemoglobin type Globin-chain composition Embryo Embryo Embryo Embryo to fetus Fetus to adult Adult Adult Gower 1 Gower 2 Portland Fetal A A2 Fetal Zeta2 , epsilon2a Alpha2, epsilon2 Zeta2, gamma2 Alpha2, gamma2 Alpha2, beta2 Alpha2, delta2 Alpha2, gamma2b a This tetramer may be an epsilon tetrad. b Fetal hemoglobin produced by adults has a different amino acid heterogeneity of the gamma chain at the 136 position than fetal Dr. Wolf's CHM 424 hemoglobin 27- 279 Inherited Hemoglobin disorder Definition: An inherited mutation of the globin genes leading to a qualitative or quantitative abnormality of globin synthesis Dr. Wolf's CHM 424 27- 280 The Thalassemias (quantitative) Syndromes in which the rate of synthesis of a globin chain is reduced beta thalassemia - reduced beta chain synthesis alpha thalassemia – reduced alpha chain synthesis Dr. Wolf's CHM 424 27- 281 Alpha Thalassemias Rare, since gene is duplicated (four genes per diploid). Usually more severe than beta thalassemia because there is no substitute for gene in adults. Almost all thalassemias are deletions In thalassemia major (0/00) - occurrence of HbH (4) and Hb Bart’s (4). BPG is ineffective in HbH & Hb Bart’s. Dr. Wolf's CHM 424 27- 282 Beta thalassemia Impaired production of beta chain beta thalassemia minor – heterozygous (or trait) beta thalassemia major - homozygous Dr. Wolf's CHM 424 27- 283 Beta thalassemia - heterozygous (minor or trait) Target cell Oval cell Dr. Wolf's CHM 424 27- 284 Beta thalassemia major Dr. Wolf's CHM 424 27- 285 Beta Thalassemias More common, since gene is present in only one copy per chromosome. Less severe than thalassemia, since d chain can effectively substitute in adults. The chain can also persist into adulthood (HPFH). In d thal major (d0/d0) excess chains do not form soluble homotetramers. Dr. Wolf's CHM 424 27- 286 Beta thalassemia major No beta chain produced (no HbA) Severe microcytic anemia occurs gradually in the first year of life Marrow expansion Iron overload Growth failure and death Dr. Wolf's CHM 424 27- 287 Dr. Wolf's CHM 424 27- 288 Alpha thalassemia / Normal / - Mild microcytosis /- Mild microcytosis -/- - Hemoglobin H disease - -/- - Hemoglobin Barts – Hydrops Fetalis Dr. Wolf's CHM 424 27- 289 Structural hemoglobinopathy (qualitative) Amino acid substitution in the globin chain e.g. sickle hemoglobin (HbS) Dr. Wolf's CHM 424 27- 290 Sickle cell hemoglobin Glu Glu - - Glu HbS (heterozygous) Sickle cell trait HbS (homozygous) Sickle cell disease - HbA1 Dr. Wolf's CHM 424 27- 291 Dr. Wolf's CHM 424 27- 292 Red Blood Cells from Sickle Cell Anemia Deoxygenation of SS erythrocytes leads to intracellular hemoglobin polymerization, loss of deformability and changes in cell morphology. OXY-STATE Dr. Wolf's CHM 424 DEOXY-STATE 27- 293 Sickle Cell Anemia – blood film Sickle Cells Erythroblasts HowellJolly Body Dr. Wolf's CHM 424 27- 294 Fibres of Sickle Hemoglobin Dr. Wolf's CHM 424 27- 295 Fibres of Sickle Hemoglobin – cross section Dr. Wolf's CHM 424 27- 296 Hemoglobin S Valine is exposed in deoxy-Hemoglobin Dr. Wolf's CHM 424 27- 297 Polymerization of HbS Dr. Wolf's CHM 424 27- 298