Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
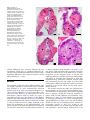
Anat Embryol (2006) DOI 10.1007/s00429-006-0086-0 O R I GI N A L A R T IC L E Matı́as Pandolfi Æ Fabiana L. Lo Nostro Akio Shimizu Æ Andrea G. Pozzi Æ Fernando J. Meijide Graciela Rey Vazquez Æ M. Cristina Maggese Identification of immunoreactive FSH and LH cells in the cichlid fish Cichlasoma dimerus during the ontogeny and sexual differentiation Accepted: 24 February 2006 Springer-Verlag 2006 Abstract Follicle-stimulating hormone (FSH) and luteinizing hormone (LH) expressing cells were detected in pituitary, brain and ovary of the Perciform cichlid fish Cichlasoma dimerus. This detection was carried out by immunohistochemistry (IHC) and Western blot techniques using antisera of the Cyprinodontiform Fundulus heteroclitus raised against the conservative region of the teleost bFSH and the bLH subunits. The estimated molecular weights were 24 kDa for LH and 19 and 15 kDa for FSH. In the adult pituitary, both cell types were distributed along mid and ventral zones of the proximal pars distalis (PPD, mid-immunoreactive cells), and along the ventral and dorsal external border of the pars intermedia (PI, high-immunoreactive cells). Double IHC showed that FSH and LH are mainly expressed in different pituitary cells. FSH cells were detected in the pituitary around day 21 after hatching (ah) (prior to sex differentiation), while LH cells were detected by day 60 ah (during the sexual differentiation period). A correlation between gonadal sex differentiation and FSH was demonstrated in a 15 days organ culture system. FSH and LH neurons were localized in the nucleus lateralis tuberis and their fibers project through the ventral hypothalamus, preoptic area and neurohypophysis. FSH neurons differentiated on day 21 ah, while LH neurons appeared on day 15 ah. In the ovary, the immunoreactivity for both FSH and LH was restricted to the cytoplasm of previtellogenic and early vitellogenic oocytes. Keywords Teleostei Æ Cichlidae Æ Gonadotropins Æ Sexual differentiation Æ Ontogeny Introduction M. Pandolfi (&) Æ F. L. Lo Nostro Æ F. J. Meijide G. R. Vazquez Æ M. C. Maggese Laboratorio de Embriologı́a Animal, Departamento de Biodiversidad y Biologı́a Experimental, Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires, Ciudad Universitaria, Pabellón 2, Buenos Aires C1428EHA, Argentina E-mail: pandolfi@bg.fcen.uba.ar Tel.: +54-11-45763348 Fax: +54-11-45763384 M. Pandolfi Æ F. L. Lo Nostro Æ A. G. Pozzi Æ M. C. Maggese Consejo Nacional de Investigaciones y Tecnicas (CONICET), Departamento de Biodiversidad y Biologı́a Experimental, Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires, Ciudad Universitaria, Pabellón 2, Buenos Aires C1428EHA, Argentina A. Shimizu National Research Institute of Fisheries Science, Fisheries Research Agency, Kanazawa, Yokohama 236-8648, Japan A. G. Pozzi Laboratorio de Biologı́a del Desarrollo, Departamento de Biodiversidad y Biologı́a Experimental, Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires, Ciudad Universitaria, Pabellón 2, Buenos Aires C1428EHA, Argentina Follicle-stimulating hormone (FSH) and luteinizing hormone (LH) are glycoproteins that are synthesized in the vertebrate pituitary, and their principal role is the control of gametogenesis and gonadal steroidogenesis in adults. In several tetrapod and teleost species, two different types of gonadotropins (GtHs) were isolated and it was demonstrated that each GtH contains two different subunits named a and b (Yoshiura et al. 1999; Yaron et al. 2001). In a given vertebrate species, the aFSH and aLH subunits are identical while b subunits are different, which gives the specific biological function of each GtH. In salmonid species, characterized for spawning only once a year, the different roles of FSH and LH were studied in detail (Suzuki et al. 1988; Tyler et al. 1991). The functions of FSH as well as its variation along the reproductive cycle are not very clear in salmonid species examined so far. In general, FSH is considered as a vitellogenic hormone, while LH is related with gonadal maturation and steroidogenesis. Several studies conducted during the development of teleost fishes, demonstrated different plasma levels and different expression patterns of FSH and LH in the pituitary (Dufour et al. 2000). Taken together, these results suggest specific functions of FSH and LH in the onset of puberty and control of gametogenesis. Traditionally it was thought that LH and FSH were exclusively synthesized in vertebrate pituitary cells. However, immunoreactivity to these hormones was detected in rodent and human brains (Croxatto et al. 1964; Emanuelle et al. 1981; Hostetter et al. 1981). Likewise, in the salmon Oncorhynchus nerka, it was reported that some neurons from preoptic nuclei expressed FSH and LH-like glycoproteins (Parhar et al. 1995). In the tilapia Oreochromis niloticus, FSH and LH were cloned and sequenced, and the presence of their respective mRNAs in the brain was confirmed by RT-PCR (Parhar et al. 2003). Although the precise functions of the brain-derived FSH and LH are still unknown, the distribution of their ir-axons along the brain and neurohypophysis suggest a probable neuromodulator function in brain nuclei related with the olfactory system and the control of pituitary endocrine cells. On the other hand, cell bodies that express FSH and LH belong to preoptic areas strongly associated with reproductive behavior (Demski and Sloan 1985, Foran and Bass 1999). A few studies were focused on the ontogeny and functions of GtHs during early development: salmon Salmo gairdneri (Van der Hurk 1982), carp Cyprinus carpio (Van Winkoop et al. 1987), rainbow trout O. mykiss (Saga et al. 1993; Feist and Schreck 1996), platyfish Xiphophorus maculates (Magliulo Cepriano et al. 1994), pejerrey Odontesthes bonariensis (Miranda et al. 2001) and gilthead seabream Sparus aurata (Garcia Ayala et al. 2003). These studies revealed a marked species-specific variation in the chronological appearance of LH and FSH cells during the ontogeny. Particularly, the identification of FSH cells using heterologous antisera was very difficult to perform due to cross-reactions or low specificity between FSH molecules from different species (Vissio et al. 1996; Pandolfi et al. 2001a). Antisera raised against the mummichog Fundulus heteroclitus FSH and LH were successfully used to recognize GtH cells in several species of teleost fishes, because they recognized conserved regions of FSH and LH from teleosts (Shimizu et al. 2003a, b). One of the aims of this study was to identify GTH cells in adult pituitary and brain, and during the early development of the South American cichlid fish Cichlasoma dimerus. This freshwater species adapts easily to captivity and spawns with high frequency during 8 months of the year, providing an appropriate model for developmental studies. In terms of gonadal development, this species was classified as a differentiated gonochorist, in which ovarian differentiation (day 42) preceded testicular differentiation (day 72), revealed by the onset of meiosis (Meijide et al. 2005). A correlation between the innervation of the pituitary gland by GnRHI fibers and the onset of the sexual differentiation process was already reported for this species suggesting that GTHs may be involved in this process (Pandolfi et al. 2002). The sec- ond aim of this study was to evaluate the effect of FSH on the gonadal differentiation in vitro. Materials and methods Animals and tissue processing Thirty C. dimerus adult fish (12 males and 18 females, 11 ± 2 cm and 45 ± 8 g) were collected from Esteros del Riachuelo, Corrientes, Argentina (2725¢S 5815¢W) and kept in large aquaria at 27 ± 1C and 12:12 h photoperiod. Aquaria were well aerated and provided with external filtration. Larvae were obtained from natural spawnings of adult specimens. Adult animals were sacrificed during the reproductive season (Spring– Summer). Pituitaries, brains and gonads were fixed in Bouin’s solution for 24 h. Samples were then dehydrated and embedded in paraplast (Fisherbrand, Fisher, WA, USA). For immunohistochemistry (IHC) techniques, pituitaries were sagittally sectioned at 7 lm while brains and gonads were transversally sectioned at 10 lm. Some other sections, randomly selected, were stained with hematoxilin–eosin, Masson tricromic and periodic-acid schiff (PAS) techniques to further characterize the morphology of gonadotrops. For developmental IHC analysis, daily samples of larvae were obtained from hatching to 15 days after hatching (ah), and every 3 days from day 15 to 90 ah. At each stage, ten animals were used. Antisera Antisera used for this study were raised in rabbit against bFSH and bLH of the cyprinodontiform F. heteroclitus (mummichog) (Fh). The generation and characterization of these antisera were previously described (Shimizu and Yamashita 2002). These antisera were specifically raised against conservative sites of teleost GtHs. For this study, anti-Fh (50-60) bFSH and anti-Fh (91-106) bLH were used. Epitope unmasking In order to enhance antigen immunoreactivity, sections were treated for epitope unmasking after deparaffination and rehydratation. Sections were dipped in an epitope unmasking solution (Target Unmasking Fluid, TUF, Sanbio BV) for 10 min at 90C, cooled at room temperature (RT) and finally washed in distilled water. Single immunohistochemistry After the epitope unmasking treatment, sections were rewashed in phosphate buffered saline (PBS, pH 7.4) and incubated for 15 min in a 0.75% gelatine solution and 30 min in PBS containing 5% non-fat dry milk at RT (blocking solution). Next, they were incubated with the primary antisera anti-bFSH (1:1,000) or anti-bLH (1:2,000) for 18 h at RT. The sections were then washed in PBS and incubated for 45 min at RT with a biotynilated anti-rabbit IgG (1:600) (Vector LAB, Burlingame, CA). Amplification of the signal was achieved by incubating the sections with a peroxidase-conjugated streptavidin (Dako, Carpinteria, CA) diluted 1:300 for 1 h at RT. After three washes in PBS, peroxidase activity was visualized with 0.1% 3,3¢-diaminobenzidine (DAB) in TRIS buffer (pH 7.6) and 0.03% H2O2. Sections were lightly counterstained with hematoxylin, mounted, examined with a NIKON microphot FX microscope and digitally photographed (Nikon, Coolpix 4500). To confirm the specificity of the immunostaining, control sections were incubated with the primary antisera (in their working dilution) preabsorbed with an excess of its respective antigen (100 ng/ll). To avoid false positives caused by the IHC itself, replacement of primary antisera with PBS and omission of secondary antisera were also performed. bFSH and bLH double immunohistochemistry After the epitope unmasking treatment, some sections were washed in distilled water and then in PBS for 20 min. After that, they were preincubated in blocking solution (5% non-fat dry milk) for 1 h and finally with the anti-Fh (91-106) bLH diluted 1:2,000 for 18 h at RT. Sections were then washed in PBS and incubated for 1 h at RT with an anti-rabbit IgG coupled to peroxidase (1:200) (Dako, Carpinteria, CA). Sections were washed 10 min in PBS and developed with DAB. As a result, bLH expressing cells showed a dark brown color. As both primary antisera were raised in rabbit, it was necessary to perform a blocking protocol against free epitopes in order to avoid interactions between the first and second antigen detection systems. After developing bLH cells with DAB, sections were washed again in distilled water and then deep in a 10 mM glicine-HCl (pH 2) solution for 1 h at 40C and after that, in a 8 M urea solution for 1 h at 40C. Sections were then washed in distilled water and PBS and preincubated again in blocking solution for 1 h at RT, and finally with the anti-Fh (50-60) bFSH for 18 h at RT. After a 10 min wash in PBS, sections were incubated with a 1:200 dilution of anti-rabbit IgG coupled to alkaline phosphatase (Dako, Carpinteria, CA) for 1 h at RT. Sections were washed for 10 min and then developed using an alkaline phosphatase kit (BCIP/NBT, Vector Blue, Dako). As a result of this technique, bFSH expressing cells displayed a dark blue color. Western blot analysis To test the specificity of F. heteroclitus bFSH and bLH antisera in C. dimerus, an analysis on 15% sodium dodecylsulfate-polyacrylamide gel electrophoresis (SDSPAGE) followed by Western blot was performed. Five adult males and five adult females were anesthetized with 0.1% benzocaine and killed by decapitation. Ten pituitaries and two ovaries were homogenized in 1 ml of Tris–HCl buffer 50 mM pH 7.4 with 10 ll of protease inhibitor cocktail (Sigma, Saint Louis, MO). Samples were processed for SDS-PAGE followed by Western blot as was previously described for this species (Pandolfi et al. 2005). The primary antisera dilutions were 1:2,000 for anti-Fh (50-60) bFSH and 1:4,000 for anti-Fh (91-106) bLH. Control lanes of pituitary homogenates were performed by replacing the primary antisera with Tris buffer and by incubating pituitary homogenates with the preadsorbed primary antibodies in their working dilution with and excess of LH or FSH (Fig. 2e, f). Treatment of undifferentiated gonads using recombinant human FSH (rhFSH) in vitro Eighty 30-day-old larvae were anesthetized in a 0.1% benzocaine solution, quickly washed in 70% ethanol and transferred to sterilized capsules containing physiological solution. The undifferentiated gonads, together with the kidney and some portions of the dorsal body wall, were dissected under stereoscopic microscope and transferred to multi-well culture plates containing 300 ll of culture media (L15 in a 60% V/V dilution, supplemented with 10% fetal bovine serum, 10 mM Hepes, 100 IU/ml of penicillin and 100 lg/ml of streptomycin). Gonads were maintained in culture for 15 days at 25C and the media was completely changed every 2 days. Two independent experiments with N = 40 each were performed. The viability of the gonads (measured by the cytoarchitecture of fixed gonads after 15 days in culture) was always above 80% in every treatment. The following protocol was used: Days 1–3 (30–33 days ah): The 40 gonads only with culture media. Days 4–11 (34–41 ah): 10 gonads only with culture media (control), and 30 gonads with the three different concentrations of rhFSH (10 gonads/concentration: 0.5, 1 and 2 lg/ml). Days 12–15 (42–45 ah): Again, the 40 gonads only with culture media. The rhFSH concentrations were chosen based on some published works related to the effect of this hormone on the onset of meiosis (Maekawa et al. 1995; Ito and Ave 1999; Yazawa et al. 2002). After 15 days in culture media, gonads were fixed in Bouin’s solution and embedded in paraffin, then sectioned and stained with hematoxilin–eosin. The onset of meiosis in control and treated group was observed. Data analysis was performed with non-parametric statistics using a frequency analysis with a v2 test and William’s correction for adjusting the analysis to our sample size (N). Results Localization of bFSH and bLH-ir cells in the adult pituitary The pituitary of C. dimerus has two clearly different zones: the adenohypohysis (ADH) and the neurohypohysis (NH), and, as in all teleosts, the NH sends deep branches to the ADH, especially into the pars intermedia (PI). Gonadotrops are PAS + basophilic cells localized mainly in the proximal pars distalis (PPD) and the external border of the PI. bLH expressing cells A sagittal pituitary section inmunostained with anti-Fh (91-106) bLH is shown in Fig. 1a. A moderated LH-ir cell population is located in central, ventral and Fig. 1 Single label immunohistochemistry (IHC). a Sagittal section of an adult pituitary immunostained with anti-Fh (91-106) bLH. b Details of LH-ir cells in central zones of the PPD. Typical vacuolated cytoplasm can be observed in the inset. c Detail of high LH-ir cells in the external border of the PI. d Sagittal section of an adult pituitary immunostained with anti-Fh (50-60) bFSH. e FSH-ir cells (arrowheads) in central zones of the PPD. f FSH-ir cells in the external border of the PI where some FSH ir-fibers could be observed in the NH. Scale bars: 20 lm. HYP hypothalamus, IR infundibular recess, PI pars intermedia, PPD proximal pars distalis, RPD rostral pars distalis marginal zones of the PPD (Fig. 1b). There is also a high ir-population in the external border of the PI (Fig. 1c). These cells are arranged in cords or small groups along the PPD. They have conspicuous processes directed towards the NH branches (Fig. 2c), and they show a vacuolated cytoplasm (Fig. 1b, inset). bFSH expressing cells A sagittal pituitary section inmunostained with anti-Fh (50-60) bFSH is observed in Fig. 1d. A population of FSH cells showing a moderate immunoreactivity is detected in central and ventral zones of the PPD (Fig. 1e). There is also a FSH high ir-population in the external border of the PI (Fig. 1f). In general, FSH-ir cells are scarce and smaller when compared with LH cells, and localized in more anterior positions of the PPD. These endocrine cells are arranged in cords of two or three cells A D B E C F or isolated. They are spherical or poliedrical in shape, and they show a vacuolated cytoplasm. A double IHC for FSH and LH in an adult pituitary section can be observed in Fig. 2a, b. LH cells show a brown color and FSH cells a blue one (Fig. 2c, d). Both FSH and LH cells present the same distribution in midsagittal and lateral sections. No apparent differences in cell number and immunoreactivity could be detected between males and females. Western blot analysis showed that C. dimerus bLH has a MW of 24 kDa in pituitary. For bFSH two specific ir-bands were obtained: 19 and 15 kDa (Fig. 2e, f). ir fibers in the NH (Fig. 1f). These ir-fibers can also be observed in the infundibular stalk, preoptic area and ventral hypothalamus. The neuronal cells bodies that send these projections are located in the preoptic area (nucleus parvocellular and magnocellular) and in the hypothalamic nucleus lateralis tuberis (Fig. 3b). Although both GtHs are expressed in the same brain areas, this expression does not occur in the same neurons. Treatment of sections with preabsorbed antisera abolished fibers and cell bodies immunostaining. Ontogeny of FSH and LH endocrine cells and neurons Localization of bFSH-ir and bLH neurons and their projections in adults bFSH expressing cells Adult pituitaries show not only ir-endocrine cells but also a lot of LH-ir fibers (Fig. 3a) and some scarce FSHFig. 2 bFSH and bLH double IHC. Double IHC in sagittal sections of an adult male (a), and an adult female (b), showing LH cells in brown and FSH cells in blue. c, d Details of FSH (blue) and LH (brown) cells in the PPD, where the LH cell processes directed towards the NH branches can be observed (arrows in c). Scale bars: 20 lm. e Western blot analysis of pituitary homogenates showing a single ir-band for LH (24 kDa) and two ir-bands for FSH (15 and 19 kDa). f Western blot analysis of pituitary homogenates incubated with the preadsorbed antibodies: 1 anti-LH preadsorbed with FSH, 2 anti-LH preadsorbed with LH, 3 anti-FSH preadsorbed with LH, 4 antiFSH preadsorbed with FSH. HYP hypothalamus, IR infundibular recess, PI pars intermedia, PPD proximal pars distalis, RPD rostral pars distalis At day 21 ah, the first pituitary FSH-ir cells are detected in the anterior ADH (Fig. 3c). Close to the onset of the sexual differentiation period (day 42 ah) the number of A B C D E F Fig. 3 Single label IHC. a LHir fibers in the NH (arrows) contacting the PPD and PI. b LH-ir cell bodies and fibers in the hypothalamic NLT. c Sagittal section at the pituitary level of 21-day-old larvae in which FSH cells could be detected in ventral zones of the PPD (arrowheads). d Coronal section of 21-day-old larvae in which a small population of FSH neurons in the NLT was detectable. e Sagittal section at the pituitary level of 15-day-old larvae in which a strong innervation of the PI by LH-ir fibers (arrowheads) could be detected. Details of hypothalamic fibers in the inset. f Sagittal section at the pituitary level of 60-day-old larvae in which the expression of LH pituitary cells starts. Scale bars: 20 lm. HYP hypothalamus, ir immunoreactive, IR infundibular recess, NH neurohypophysis, NLT nucleus lateralis tuberis, PI pars intermedia, PPD proximal pars distalis, RPD rostral pars distalis A B C D E F FSH-ir cells increases and they can be observed along the PPD and the external border of the PI. At least at day 90, when this ontogenetic study finished, the number of FSH-ir cells is still increasing. Preoptic and hypothalamic FSH-ir neurons are synchronically differentiated with FSH-ir pituitary cells at day 21 ah (Fig. 3d). bLH expressing cells At day 15 ah, some neuronal bodies in the hypothalamus sending their projections through the PI are observed (Fig. 3e). The pituitary ir-cells are detected later than the respective neurons. The differentiation of this population occurred at day 60 ah, after the sexual differentiation period was started. The first ircells are detected in central zones of the PPD (Fig. 3f). Data of this ontogenetic study is summarized in Table 1. Expression of FSH and LH in gonads Along the ontogenetic study immunoreactivity for FSH and LH was also detected in ovaries. This was evidenced by the presence of FSH and LH ir-material in the cytoplasm of the oocytes of this species. Ovaries sampled during winter season and during the pre and postspawning periods of summer season were treated for GtHs IHC. In all cases, the FSH-ir and LH material was Table 1 Ontogenetic expression of bFSH and bLH pituitary cells and neurons in C. dimerus Day after hatching Body length (mm) bFSH pituitary cells bFSH neurons 15 21 60 6 ± 0.4 8 ± 0.6 17 ± 1.5 + + + + bLH pituitary cells + bLH neurons + + + Fig. 4 Single label IHC. Transversal sections of adult female ovary. a Previtellogenic oocytes with a FSH-ir cytoplasm. b Ovarian control section treated with the preabsorbed anti-FSH. c Previtellogenic oocytes with a LH-ir cytoplasm. d Ovarian control section treated with the preabsorbed anti-LH. Controls were counterstained with hematoxylin. Scale bars: 20 lm. e Western blot analysis of ovarian homogenates showing a single ir-band for LH (24 kDa) and 2 ir-bands for FSH (15 and 19 kDa) A B C D E localized within the cytoplasm of previtellogenic and early vitellogenic oocytes (Fig. 4a, c). Ovarian control sections treated with the preabsorbed anti-FSH or antiLH are shown (Fig. 4b, d). Western blot analysis of ovarian homogenates revealed the presence of the same ir-bands detected in the pituitary (Fig. 4e). On the other hand, no immunoreactivity was detected in the testis. Table 2 Percentage of viability, and of undifferentiated and differentiated gonads in both experiments Viability (%) Undifferentiated Differentiated gonads (%) gonads (%) Control 0.5 lg/ml rhFSH 1 lg/ml rhFSH 2 lg/ml rhFSH 85 90 90 85 100 28 11 12 0 72 89 88 Significant differences were observed in the percentage of differentiated gonads among the three treatments when compared to the control. N = 20 for each treatment, P < 0.05 Effect of rhFSH on undifferentiated gonads in vitro The undifferentiated gonads belonging to 30-day-old larvae could be maintained in culture conditions for 15 days, and most of the gonads treated with rhFSH underwent sexual differentiation (Table 2). We named ‘‘undifferentiated gonads’’ (UG) those containing only undifferentiated germ cells (Fig. 5a, b), and ‘‘differentiated gonads’’ (DG) those in which early meiotic cells (oocytes) were detected (Fig. 5c, d). During normal gonadal development in C. dimerus, ovarian differentiation starts around day 42, while the onset of testicular differentiation occurs 30 days later (Meijide et al. 2005). Then, DGs at day 45 under culture conditions were considered as ovaries under the process of sex differentiation, because testicular differentiation does not occur until later in development. It is important to notice that none of the control gonads (placed 15 days in culture media without rhFSH) underwent sexual differentiation while most of the treated gonads did (Table 2). No sig- Fig. 5 Culture of undifferentiated gonads. a Transversal section of an explant treated for 15 days in control culture conditions (day 45 of larval development), in which an undifferentiated gonad was observed (arrow). b Details of undifferentiated gonad with germ cells surrounded by somatic cells. c Transversal section of an explant treated for 15 days in culture conditions containing rhFSH (day 45 of larval development), in which a differentiated gonad was observed. d Detail of a differentiated gonad with oogonia and oocytes. In all cases, sections were stained with hematoxilin–eosin. Scale bars: 20 lm. G gonad, Gc germ cell, K kidney, M muscle, MS mesentery, N notocord, Nt neural tube, Og oogonia, Oo pachytene oocytes, Sc somatic cell A B C D nificant differences were observed between the two experiments. There were no significant differences between the three rhFSH concentrations used in this study. Significant differences were observed between control and treatments (P < 0.05). Discussion The synthesis of FSH and LH in different cellular types was already reported in several teleost species: mummichog (Calman et al. 2001), mediterranean yellowtail (Garcia Ayala et al. 1998), red seabream (Kagawa et al. 1998), pejerrey (Miranda et al. 2001) and salmon (Nozaki et al. 1990). However, in other species, the use of heterologous antisera failed to detect two types of gonadotrops (Vissio et al. 1996; Saga et al. 1999; SeguraNoguera et al. 2000; Pandolfi et al. 2001a). The results of our immunohistochemical analysis revealed that in adult C. dimerus, FSH and LH are mainly produced in different cell populations of the adenohypophysis. The precise location of the different GtH cells was based on two previous anatomical and embryological studies on C. dimerus pituitary gland (Pandolfi et al. 2001a, b). In this species, FSH expressing cells are mainly located in central and ventral zones of the PPD and, in a minor proportion, in the marginal border of the PI. LH expressing cells show a similar pattern but with a higher cell density in the PPD when compared to FSH. The immunoreactivity differences observed between PPD cells (moderated) and PI cells (high) for both GTHs could reflect a differential regulation of their production according to the period chosen for this study (reproductive season). The antisera used for this study were obtained from the mummichog F. heteroclitus (Cyprinodontidae; Cyprinodontiforms, Acantopterygii). The synthetic peptides used as antigens, carried the conserved sequences of teleost FSH and LH (Shimizu and Yamashita 2002; Shimizu et al. 2003a, b). These antisera were successful on detecting gonadotrops of several acantopterygian species (Shimizu et al. 2003a, b). For the anti-Fh (50-60) bFSH, the only way of obtaining positive reactions is pre-treating sections at 90C for 10 min in the unmasking solution. It is probable that the FSH molecule experience a partial epitope masking due to the histological procedures performed (fixation, dehydration, etc.). The specificity of our GTHs detection system was reinforced by the preabsortion controls and by the Western blot of pituitary and ovary homogenates. C. dimerus bLH showed a MW of 24 kDa, 3 kDa higher than that of the mummichog bLH (21 kDa). This difference in MW may reflect a major glycosilation degree of C. dimerus bLH. For bFSH, two specific ir-bands were obtained (C. dimerus: 19 and 15 kDa; mummichog: 18 kDa). It is probable that the 15 kDa bFSH band is a degradation product of the 19 kDa one, or a different isoform recognized by the same antisera. The vacuolated cytoplasm of FSH and LH cells in adults probably corresponds to the typical development of the RER previously described by electron microscopy in several teleost species (Quesada et al. 1988; Garcı́a Ayala et al. 1998). This vacuolization of gonadotrops seems to correlate mainly with age/sexual status, because they were not observed in larvae or juveniles. During the study of the adult pituitary, the presence of FSH-ir and LH-ir fibers was detected in the NH and infundibular stalk. Serial brain sections showed that the cells bodies, which project those fibers, were located in the POA (parvocelullar and magnocelluar preoptic nuclei) and in the hypothalamus (nucleus lateralis tuberis). In the control sections, the immunostaining was completely abolished. In salmon and tilapia, FSH and LHlike immunoreactivity was found in the preoptic nucleus. Also in the tilapia, the mRNA brain levels for these GTHs were quantified and they were lower than the ones quantified for pituitary (Parhar et al. 2003). Immunoreactivity for LH was also reported in arcuate nucleus neurons, pars tuberalis cells and hypothalamic axons from rodents (Gross and Page 1979; Hostetter et al. 1981). It was then hypothetized that the brain derived LH was originated from the pituitary LH that reached the brain from the circulatory system or by selective absorption (tanocytosis) from the brain–spinal fluid (Pacold et al. 1978). Experimental evidences showed that after hypophysectomy, it is possible to measure pituitary hormones in brain (Emanuel et al. 1981; Hojvat el al. 1982). The precise role of brain-derived FSH and LH in C. dimerus is not very clear, but, the abundance of iraxons in the ventral forebrain and in the NH suggests the existence of a neuromodulatory function together with a possible function in the control of pituitary cells activity. In rodents, it was shown that an intracerebral injection of LH modulated not only the synthesis and release of pituitary LH (Emanuele et al. 1981) but also the electric activity of some hypothalamic neurons (Kawakami and Sakuma 1974; Teresawa et al. 1969). Moreover, in the tilapia it was clearly demonstrated the FSH and LH expressing neurons also express AVT, and this neurohypohysial hormone was synthesized in preoptic neurons, which are strongly related with the control of some reproductive behaviors (Demski and Sloan 1985; Foran and Bass 1999; Parhar et al. 2003). Interestingly, FSH and LH neurons are expressed in POA regions that also express GnRH1 and GnRH3 in this species (Pandolfi et al. 2005). Although this occurs in morphological different neurons, it is possible that some synapsis between GnRH and FSH and/or LH neurons take place. Future studies will be focused in the relationship between GnRH and GTHs at the brain level. Another fact that reinforces the specificity of our detection system is the difference in the timing of appearance of LH and FSH both in brain and pituitary: LH neurons (day 15), FSH pituitary cells and neurons (day 21), LH pituitary cells (day 60). In this species, FSH cells appeared prior to the onset of sexual differentiation, while LH cells were differentiated several days after this process started. Then, we decided to study only the effect of FSH on sex differentiation, because at the day the experiment started (day 30) and ended (day 45) there was no expression of pituitary LH. This variation in the timing of expression of FSH and LH was also observed in several teleost species (Mal et al. 1989; Nozaki et al. 1990; Saga et al. 1993; Magliulo-Cepriano et al. 1994; Miranda et al. 2001). The results of this studies suggested that in salmonid species and paltyfish, as in C. dimerus, the differentiation of FSH cells preceded the differentiation of LH cells. Only in pejerrey, it was shown a first appearance of LH cells. It was described that GtHs are important for the regulation of the hypothalamus–pituitary–gonad axis (Schreibman et al. 1982). In salmonids, FSH, but not LH, was involved in the gonadal initial grow and development (Saga et al. 1993). When C. dimerus undifferentiated gonads were left alone in culture media, the onset of gonadal differentiation did not occur as observed in vivo, suggesting an important role of the brain–pituitary–axis during sex differentiation in this species. We postulated that one of the factors involved in this process is FSH because its expressing cells were already differentiated at the larval period chosen for this study and because it was demonstrated to be involved in the onset of meiosis (Ito and Abe 1999; Yazawa et al. 2002). When treated with rhFSH, 80% of the gonads were differentiated to presumptive ovaries, based on the morphological description available for this species (Meijide et al. 2005). The results of this experiment reinforce our previous hypothesis about a correlation between the innervation of pituitary cells by GnRH1 fibers and the release of FSH that will be acting in any of the important steps of the sexual differentiation process (Pandolfi et al. 2002). Another extracerebral region in which FSH and LH were found is the ovary. The presence of FSH and LHlike material was detected in previtellogenic and vitellogenic oocytes cytoplasm. The specificity of this immunostaining was confirmed by preabsorption tests and western blot. A recent study in another Perciform, the seabream Sparus aurata, showed a novel expression of FSH and LH in the ovary by isolating their corresponding genes (Wong and Zohar 2004). It is probable that these ovarian-derived GtHs act as autocrine and/or paracrine factors involved in intraovarian communication between the oocytes and the follicular cells. Acknowledgments The authors would like to thank Dr. Frank van den Berg (University of Amsterdam) for his aid in the epitope unmasking protocol, and Dr. P. Vissio (Universidad de Buenos Aires) for her constant and disinterested advice. This work was supported by grants from UBACyT X-118 and PICT E-14271. References Calman B, Lin YW, Wallace RA (2001) Preparation and use of specific antibodies to the-I and-II subunits of gonadotropic hormone from Fundulus heteroclitus pituitary. Gen Comp Endocrinol 123:203–209 Croxatto H, Arrau J, Croxatto H (1964) Luteinizing hormone-like activity in human median eminence extracts. Nature 204:584– 585 Demski LS, Sloan HE (1985) A direct magnocellular–preopticospinal pathway in goldfish: implications for control of sex behavior. Neurosci Lett 55:283–288 Dufour S, Huang YS, Rosseau K (2000) Puberty in teleosts: new insights into the role of peripheral signals in the stimulation of pituitary gonadotropins. In: Proceedings of the VI International Symposium on Reproductive Physiology of Fish. University of Bergen, Bergen, pp 455–461 Emanuele N, Oslapas R, Connick E, Kirsteins L, Lawrence AM (1981) Hypothalamic LH may play a role in control of pituitary LH release. Neuroendocrinology 33:12–17 Feist G, Shreck CB (1996) Brain–pituitary–gonadal axis during early development and sexual differentiation in the rainbow trout, Oncorhynchus mykiss. Gen Comp Endocrinol 102:394– 409 Foran CM, Bass AH (1999) Preoptic GnRH and AVT: axes for sexual plasticity in teleosts fish. Gen Comp Endocrinol 116:141–152 Garcia-Ayala A, Garcia-Hernandez MP, Quesada JA, Agulleiro B (1998) Gonadotropic and thyrotropic cells from the Mediterranean yellowtail (Seriola dumerilii; Risso, 1810): immunocytochemical and ultrastructural characterization. Anat Rec 250:448–458 Garcı́a Ayala, Villaplana M, Garcı́a Hernández MP, Chaves Pozo E, Agulleiro B (2003) FSH-, LH- and TSH-expressing cells during development of Sparus aurata L. (Telesotei). An immunocytochemical study. Gen Comp Endocrinol 134:72–79 Gross DS, Page RB (1979) Luteinizing hormone and follicle stimulating hormone production in the pars tuberalis of hypophysectomized rats. Am J Anat 156:285–291 Hojvat S, Emanuele N, Baker G, Connick E, Kirsteins L, Lawrence AM (1982) Growth hormone (GH), thyroid-stimulating hormone (TSH), and luteinizing hormone (LH)-like peptides in the rodent brain: non-parallel ontogenic development with pituitary counterparts. Dev Brain Res 4:427–434 Hostetter G, Gallo RV, Brownfield MS (1981) Presence of immunoreactive luteining hormone in the rat forebrain. Neuroendocrinology 33:241–245 Ito R, Abe SI (1999) FSH-initiated differentiation of newt spermatogonia to primary spermatocytes in germ-somatic cell reaggregates cultured within a collagen matrix. Int J Dev Biol 43(2):111–116 Kagawa H, Kawazoe I, Tanaka H, Okuzawa K (1998) Immunocytochemical identification of two distinct gonadotropic cells (GTH I and GTH II) in the pituitary of bluefin tuna, Thunnus thynnus. Gen Comp Endocrinol 110:11–18 Kawakami M, Sakuma Y (1974) Responses of hypothalamic neurons to the microiontophoresis of LHRH, LH, and FSH under various levels of circulating ovarian hormones. Neuroendocrinology 15:290–307 Maekawa K, Ji ZS, Abe S (1995) Proliferation of newt spermatogonia by mammalian FSH via Sertoli cells in vitro. J Exp Zool 272:363–373 Magliulo-Cepriano L, Schreibman MP, Blüm V (1994). Distribution of variant forms of immunoreactive gonadotropin-releasing hormone and beta-gonadotropins I and II in the platyfish, Xiphophorus maculatus, from birth to sexual maturity. Gen Comp Endocrinol 94:135–150 Mal AO, Swanson O, Dickhoff WW (1989) Immunocytochemistry of the developing salmon pituitary gland. Am Zool 29, 94A Meijide FJ, Lo Nostro FL, Guerrero GA (2005) Gonadal development and sex differentiation in the cichlid fish Cichlasoma dimerus (Teleostei, Perciformes): a light- and electron-microscopic study. J Morphol 264:191–210 Miranda LA, Strüssmann CA, Somoza GM (2001) Immunocytochemical identification of GtH1 and GtH2 cells during the temperature-sensitive period for sex differentiation in pejerrey, Odontesthes bonariensis. Gen Comp Endocrinol 124:45–52 Nozaki M, Naito N, Swanson P, Dickhoff WW, Nakai Y, Suzuki K, Kawauchi H (1990) Salmonid pituitary gonadotrophs. II. Ontogeny of GTH I and GTH II cells in the rainbow trout (Salmo gairdneri irideus). Gen Comp Endocrinol 77:358–367 Pacold ST, Kirsteins L, Hojvat S, Lawrence AM, Hagen TC (1978) Biologically active pituitary hormones in the rat brain amygdaloid nucleus. Science 199:804–806 Pandolfi M, Paz DA, Maggese MC, Meijide FJ, PG Vissio (2001a) Immunocytochemical localization of different cell types in the adenohypophysis of Cichlasoma dimerus (Teleostei, Perciformes). Biocell 25:35–42 Pandolfi M, Paz DA, Maggese MC, Ravaglia MA, PG Vissio (2001b) Ontogeny of immunoreactive somatolactin, prolactin and growth hormone secretory cells in the developing pituitary gland of Cichlasoma dimerus (Teleostei, Cichlidae). Anat Embryol 203:461–468 Pandolfi M, Parhar IS, Ravaglia MA, Meijide FJ, Maggese MC, Paz DA (2002) Ontogeny and distribution of gonadotropinreleasing hormone (GnRH) neuronal systems in the brain of the cichlid fish Cichlasoma dimerus. Anat Embryol 205:271–281 Pandolfi M, Muñoz Cueto JA, Lo Nostro FL, DownsP PJL, Paz DA, Maggese MC, Urbanski H (2005) The GnRH systems of Cichlasoma dimerus (Perciformes, Cichlidae) revisited: a localization and developmental study using antibodies and riboprobes to the GnRH- associated peptides. Cell and Tiss Res 321:219–232 Parhar IS, Iwata M, Pfaff DW, Schwanzel-Fukuda M (1995) Embryonic development of gonadotropin-releasing hormone neurones in the sockeye salmon. J Comp Neurol 362:256–270 Parhar IS, Soga T, Ogawa S, Sakuma Y (2003) FSH and LH-b subunits in the preoptic nucleus: ontogenic expression in teleost. Gen Comp Endocrinol 132:369–378 Quesada J, Lozano MT, Ortega A, Agulleiro B (1988) Immunocytochemical and ultrastructural characterization of the cell types in the adenohypophysis of Sparus aurata L. Teleost. Gen Comp Endocrinol 72:209–225 Saga T, Oota Y, Nozaki M, Swanson P (1993) Salmonid pituitary gonadotrophs. III. Chronological appearence of GTH I and other adenohypophysial hormones in the pituitary of the developing rainbow trout (Oncorhynchus mykiss irideus). Gen Comp Endocrinol 92:233–241 Saga T, Yamaki K, Doi Y, Yoshizuka M (1999) Chronological study of the appearance of adenohyphophysial cells in the ayu (Plecoglossus altivelis). Anat Embryol 200:469–475 Schreibman MP, Margolis-Kazan H, Halpern-Sebold L (1982) Immunoreactive gonadotropin and luteinizing hormone releasing hormone in the pituitary gland of neonatal platyfish. Gen Comp Endocrinol 47:385–391 Segura-Noguera MM, Laiz-Carrión L, del Rı́o MP, Mancera JM (2000) An immunocytochemical study of the pituitary gland of the white seabream (Diplodus sargus). Histochem J 32:733–742 Shimizu A, Yamashita M (2002) Purification of mummichog (Fundulus heteroclitus) gonadotropins and their subunits, using an immunochemical assay with antisera raised against synthetic peptides. Gen Comp Endocrinol 125:79–91 Shimizu A, Tanaka H, Kagawa H (2003a) Immunocytochemical applications of specific antisera raised against synthetic fragment peptides of mummichog GtH subunits: examining seasonal variations of gonadotrophs (FSH cells and LH cells) in the mummichog and applications to other acanthopterygian fishes. Gen Comp Endocrinol 132:35–45 Shimizu A, Sakai T, Nashida K, Kagawa H (2003b) Universal antisera for immunocytochemical identification of two different gonadotrophs in acanthoptherygian fishes. Fish Physiol Biochem 29:275–287 Suzuki K, Kawauchi H, Nagahama Y (1988) Isolation and characterization of two distinct gonadotropins from chum salmon pituitary glands. Gen Comp Endocrinol 71:292–301 Terasawa EI, Whitmoyer DI, Sawyer CH (1969) Effects of luteinizing hormone on multiple-unit activity in the rat hypothalamus. Am J Physiol 217:1119–1126 Tyler CR, Sumpter JP, Kawauchi H, Swanson P (1991) Involvement of gonadotropin in the uptake of vitellogenin into vitellogenic oocytes of the rainbow trout, (Oncorhynchus mykiss). Gen Comp Endocrinol 84:291–299 Van der Hurk R (1982) Effects of steroids on gonadotropic (GtH) cells in the pituitary of rainbow trout, Salmo gairdneri, shortly after hatching. Cell Tiss Res 224:361–368 Van Winkoop A, Timmermans LPM, Booms GHR (1987) The expression of germ cell differentiation antigens, as defined with monoclonal antibodies, in correlation with the ontogeny of gonadotropic cells in the hypophysis of carp. In: DR Ilder, LW Crim, Jm Walsh (eds) Proceedings of the Third International Symposium on the Reproductive Physiology of Fish. St Johńs, New Foundland, Canada, p 222 Vissio PG, Paz DA, Maggese MC (1996) The adenohypophysis of the swamp eel, Synbranchus marmoratus, an immunocytochemical analysis. Biocell 20:155–161 Wong T, Zohar Y (2004) Novel expression of gonadotropin subunit genes in oocytes of the gilthead seabream (Sparus aurata). Endocrinology 145:5210–5220 Yaron Z, Gur G, Melamed P, Rosenfeld H, Levavi-Sivan B, Elizur A (2001) Regulation of gonadotropin subunit genes in tilapia. Comp Biochem Physiol Part B 129:489–502 Yazawa T, Yamamoto T, Jin Y, Abe S (2002) Follicle-stimulating hormone is indispensable for the last spermatogonial mitosis preceding meiosis initiation in newts (Cynops pyrrhogaster). Biol Reprod 66:14–20 Yoshiura Y, Suetake H, Aida K (1999) Duality of gonadotropin in a primitive teleost, Japanese eel (Anguilla japonica). Gen Comp Endocrinol 114:121–131