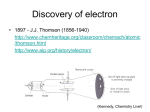
Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
MASS SPECTROMETRY OF OLIGOSACCHARIDES Joseph Zaia* Department of Biochemistry, Boston University School of Medicine, 715 Albany St., R-806, Boston, Massachusetts 02118 Received 20 March 2003; received (revised) 6 August 2003; accepted 6 August 2003 I. II. III. IV. V. Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Characteristics of Tandem Mass Spectra of Carbohydrates . . . . . . . . . . . . . . . . . . . . . . . . . . . . A. Ionization of Carbohydrates . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1. Electrospray Ionization (ESI) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2. Matrix-Assisted Laser Desorption/Ionization (MALDI) . . . . . . . . . . . . . . . . . . . . . . . . B. Nomenclature for the Fragmentation of Glycoconjugates . . . . . . . . . . . . . . . . . . . . . . . . . . C. Tandem MS of Native Oligosaccharide Molecular Ions . . . . . . . . . . . . . . . . . . . . . . . . . . . 1. Protonated Ions . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2. Deprotonated Ions . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3. Alkali and Alkaline Earth Adducted Ions . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . D. Tandem MS of Permethylated and Peracetylated Oligosaccharides . . . . . . . . . . . . . . . . . . E. Tandem MS of Reductively Aminated Carbohydrates . . . . . . . . . . . . . . . . . . . . . . . . . . . F. Discrimination of Monosaccharide Linkages . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . G. Gas-Phase Degradation of Oligosaccharides . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . H. Computer-Based Approaches for Interpretation of Oligosaccharide Product-Ion Mass Spectra I. Internal Residue Loss Rearrangements of Oligosaccharide Ions During CID . . . . . . . . . . . . J. Conclusions . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Analyzers for Mass Spectrometry of Carbohydrates . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . A. Analysis of Permethylated Carbohydrates Using High Temperature GC/MS . . . . . . . . . . . . B. Analysis of Carbohydrates with MALDI-TOF MS . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . C. Analysis of Carbohydrates with MALDI Q-oTOF MS . . . . . . . . . . . . . . . . . . . . . . . . . . . D. Analysis of Carbohydrates with ESI Q-oTOF MS . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . E. Analysis of Carbohydrates with QIT MS . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . F. Analysis of Glycoconjugates with FT MS . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . G. Conclusions . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Tandem Mass Spectrometry of Glycopeptides . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . A. Ionization of Glycopeptides . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . B. CID of Glycopeptides . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1. Selective Identification of Glycopeptides with Tandem MS . . . . . . . . . . . . . . . . . . . . . 2. CID of O-Linked Glycopeptides . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3. CID of N-Linked Glycopeptides . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . C. Electron Capture Dissociation of Glycopeptides . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . D. Conclusions . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Mass Spectrometry of Sialylated Glycoconjugates . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . A. Permethylation of Sialylated Oligosaccharides . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . B. MALDI-MS of Sialylated Glycoconjugates . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1. Anionic Dopants for Analysis of Sialylated Glycoconjugates . . . . . . . . . . . . . . . . . . . . 2. Methyl Esterification to Stabilize Sialic Acid Residues . . . . . . . . . . . . . . . . . . . . . . . . 3. Perbenzolylation to Stabilize Sialic Acid Residues . . . . . . . . . . . . . . . . . . . . . . . . . . . 4. High-Pressure MALDI of Sialylated Glycoconjugates . . . . . . . . . . . . . . . . . . . . . . . . . ———— Contract grant sponsor: NIH/NCRR; Contract grant number: P41RR10888; Contract grant sponsor: Glycosciences Research Award, Neose Technologies. *Correspondence to: Joseph Zaia, Department of Biochemistry, Boston University School of Medicine, 715 Albany St., R-806, Boston, MA 02118. E-mail: [email protected] Mass Spectrometry Reviews, 2004, 23, 161– 227 # 2004 by Wiley Periodicals, Inc. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 162 . 163 . 163 . 163 . 163 . . 164 . 165 . 165 . 166 . 166 . 169 . 170 . 171 . 178 . 181 . 183 . 184 . 186 . 186 . 186 . 187 . 187 . 187 . 188 . 188 . 189 . 189 . 190 . 190 . 191 . 191 . 193 . 193 . 195 . 195 . 195 . 196 . 196 . 196 . 196 & ZAIA . . . . . . . . . . . . . . . . . . . . . . . . . 197 198 199 199 199 199 199 202 202 202 203 204 205 206 206 206 207 208 208 209 211 211 214 214 215 Abbreviations . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 215 References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 216 VI. VII. VIII. C. ESI MS of Sialylated Oligosaccharides . . . . . . . . . . . . . . . . . . . . . . . . . D. Tandem MS of Sialylated Oligosaccharides . . . . . . . . . . . . . . . . . . . . . . . E. Conclusions . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Mass Spectrometry of Sulfated Oligosaccharides . . . . . . . . . . . . . . . . . . . . . . A. Derivatization . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . B. Ionization Methods . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1. Fast Atom Bombardment . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2. MALDI . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . a. MALDI of Sulfated Peptides . . . . . . . . . . . . . . . . . . . . . . . . . . . b. Direct MALDI of Sulfated Oligosaccharides . . . . . . . . . . . . . . . . c. Use of Basic Peptides for MALDI of Polysulfated Oligosaccharides d. MALDI Analysis of Protein-Sulfated Oligosaccharide Complexes . 3. ESI of Sulfated Oligosaccharides . . . . . . . . . . . . . . . . . . . . . . . . . . . 4. On-Line Separation Systems for Sulfated Carbohydrates . . . . . . . . . . . C. Tandem MS of Sulfated Oligosaccharides . . . . . . . . . . . . . . . . . . . . . . . . 1. Lessons from CID of Sulfated Peptides . . . . . . . . . . . . . . . . . . . . . . . 2. Tandem MS of Mono- and Di-Sulfated Oligosaccharides . . . . . . . . . . . 3. Precursor-Ion and Neutral-Loss Scans for Sulfated Glycoconjugates . . . 4. Determination of Positional Sulfation Isomers in GAG Disaccharides . . 5. Tandem Mass Spectrometric Quantification of GAG Disaccharides . . . . 6. Tandem Mass Spectrometric Analysis of GAG Oligosaccharides . . . . . a. CS Oligosaccharides . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . b. Heparin/HS Oligosaccharides . . . . . . . . . . . . . . . . . . . . . . . . . . . D. Conclusions . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Overall Conclusions . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Glycosylation is a common post-translational modification to cell surface and extracellular matrix (ECM) proteins as well as to lipids. As a result, cells carry a dense coat of carbohydrates on their surfaces that mediates a wide variety of cell–cell and cell– matrix interactions that are crucial to development and function. Because of the historical difficulties with the analysis of complex carbohydrate structures, a detailed understanding of their roles in biology has been slow to develop. Just as mass spectrometry has proven to be the core technology behind proteomics, it stands to play a similar role in the study of the functional implications of carbohydrate expression, known as glycomics. This review summarizes the state of knowledge for the mass spectrometric analysis of oligosaccharides with regard to neutral, sialylated, and sulfated compound classes. Mass spectrometric techniques for the ionization and fragmentation of oligosaccharides are discussed so as to give the reader the background to make informed decisions to solve structure-activity relations in glycomics. # 2004 Wiley Periodicals, Inc., Mass Spec Rev 23:161–227, 2004 Keywords: carbohydrates; oligosaccharides; mass spectrometry; sialylated; sulfated; glycomics I. INTRODUCTION With progress in proteomics comes an increasing interest in the importance of glycosylation. Most cell-surface and 162 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . secreted proteins are glycosylated—a fact that impacts on efforts to understand the biological relevance of specific protein expression and modification patterns. Unlike the core proteins, glycans are expressed as a set of variations on a core structure and are polydisperse in nature. Therefore, glycosylation increases the complexity of protein molecules and causes them to migrate as diffuse bands or spots on SDS–PAGE gels to complicate efforts to identify protein expression patterns that correlate with disease states. Glycans serve to modulate protein function and may influence folding, biological lifetime, and recognition of binding partners. For example, surface carbohydrates serve as the interface between the cell and its environment, and define self versus non-self. Many pathogens recognize particular carbohydrates, and structural studies have led to progress in this area. Mass spectrometry is an important tool for the structural analysis of carbohydrates, and offers precise results, analytical versatility, and very high sensitivity. Whereas mass spectrometric analysis options for proteins and peptides are well-defined relative to those for carbohydrates, tandem mass spectrometric product-ion patterns are more complex, and the results depend on the types of derivative and precursor ion used. In most cases, one analyzes protonated forms of underivatized proteins and peptides and collisional-induced dissociation (CID) results MASS SPECTROMETRY OF OLIGOSACCHARIDES in definition of a complete sequence (Biemann & Martin, 1987) or of a partial sequence that is useful in the identification of modified peptide residues (Papayannopoulos, 1995) or in database searching (Mann & Wilm, 1994). With carbohydrates, one has the choice of using peralkyl or reducing terminal derivatives, or of analyzing native structures. One also has the choice of protonated, metalcationized, or deprotonated ions. As a result, the carbohydrate mass spectrometry field is reflected by a divergence of published structural analysis methods in recent years. For protein mass spectrometrists who wish to learn about carbohydrates and glycobiologists who wish to learn mass spectrometry, the plethora of publications that describe seemingly divergent approaches may seem overwhelming. The intent of this review is to summarize the historical and recent developments in carbohydrate mass spectrometry to facilitate informed decision-making on analysis routes. Section II explains common ionization methods used for carbohydrates as well as characteristic fragmentation methods for different carbohydrate compound classes. Section III describes the characteristics of different modern mass analyzers used for carbohydrates. The remaining sections specifically address the analysis of glycopeptides (Section IV), sialylated glycoconjugates (Section V), and sulfated oligosaccharides (Section VI). II. CHARACTERISTICS OF TANDEM MASS SPECTRA OF CARBOHYDRATES A. Ionization of Carbohydrates 1. Electrospray Ionization (ESI) Conventional ESI MS (Meng, Mann, & Fenn, 1988; Fenn et al., 1990) involves the pumping of a solution (a forced flow) into the ion source, and has been observed to produce relatively weak ion signals for native oligosaccharides compared to those for peptides and proteins (Burlingame, Boyd, & Gaskell, 1994; Reinhold, Reinhold, & Costello, 1995). Nano ESI (Wilm & Mann, 1994), on the other hand, produces ion signals that are comparable between the peptide and carbohydrate compound classes (Bahr et al., 1997). It, therefore, appears that the hydrophilicity of oligosaccharides limits the surface activity in ESI droplets and that, with small droplets, their sensitivity is significantly enhanced. The results with nano ESI are consistent with the conclusion that the sensitivity increase observed when oligosaccharides are derivatized, reducing their hydrophilicity, is due to an increase in surface activity rather than in volatility (Karas, Bahr, & Dulcks, 2000). The fact that the ESI of carbohydrates appears to be more effective with the nano scale, has important implications. Interfaces for on-line ESI LC/MS typically produce droplet sizes that are large relative to those produced by spraying & from a 1–2 mm orifice nanospray emitter, and thus the spray characteristics are typical of forced flow. An exception appears to be the use of 10 mm fused-silica spray tips with post-column splitting of HPLC effluents (Gangl et al., 2001). It may be possible to design a continuous-flow interface that produces droplet sizes small enough to produce nanospray-like ionization responses for carbohydrates that elute from a chromatography column. 2. Matrix-Assisted Laser Desorption/Ionization (MALDI) The MALDI-TOF ionization efficiency for neutral carbohydrates oligomers has been observed to be constant as the size of the molecule increases, in contrast to that for ESI, where the ionization efficiency decreases with an increasing molecular weight (Harvey, 1993). Although the ionization response drops off with increasing molecular weight for mixtures of large carbohydrate polymers, the use of MALDI for neutral oligosaccharide analysis has advantages over ESI, particularly for applications that involve the profiling of mixtures released from glycoproteins. Quantitation of permethylated carbohydrate mixtures with MALDI has been demonstrated to be reproducible, and the coefficients of variation for those analyses equal those obtained for the same oligosaccharide mixture after derivatization with a chromophore and chromatographic quantitation (Viseux et al., 2001). In addition, information on fucosylation was more detailed with the mass spectrometric approach. The advantages of MALDI in terms of ionization response have to be balanced against the disadvantages of the metastable fragmentation that is caused by the higher internal energies imparted to the ions relative to those resulting from ESI. Although that fragmentation allows the analysis of carbohydrate ions with post-source decay (PSD) on a MALDI-TOF instrument (Viseux, Costello, & Domon, 1999), it complicates MS profiling. Extensive fragmentation of sialic acid residues necessitates either the use of linear mode MALDI-TOF to avoid separating the metastable products (see Section V.D), or of derivatization to stabilize those groups (See Section V.B.2). As a further complication, detection of sialylated glycoconjugates with Fourier transform (FT) MS is limited by the metastable fragmentation in a low-pressure MALDI source and is mitigated through the use of collisional cooling with high pressure (O’Connor & Costello, 2001; O’Connor, Mirgorodskaya, & Costello, 2002). Atmospheric-pressure sources have been developed for quadrupole ion trap (QIT) instruments (Laiko, Moyer, & Cotter, 2000; Moyer & Cotter, 2002; Moyer et al., 2002), and it is likely that those sources will prove to be useful for the analysis of oligosaccharides. The development of a MALDI TOF/TOF analyzer has been described for the analysis of peptides 163 & ZAIA (Medzihradszky et al., 2000; Yergey et al., 2002), and carries the potential benefits of producing high-energy fragmentation. Metastable fragmentation has been a problem with that instrument, and it is likely that its effective use for the analysis of oligosaccharides will require a high-pressure source to limit metastable fragmentation, and thereby allow true high-energy CID. The benefits of high-energy CID of carbohydrates are welldocumented (Domon & Costello, 1988b; Gillece-Castro & Burlingame, 1990; Peter-Katalinic, 1994); see Section II.F for a detailed discussion. B. Nomenclature for the Fragmentation of Glycoconjugates Collisional-induced dissociation (CID) of glycoconjugates results in the observation of ions that correspond to cleavage of the oligosaccharide portion of the molecule. Typically, those ions are produced in greater abundances for the oligosaccharide portion than are those that occur in the aglycon (non-oligosaccharide) portion of glycoconjugates. The nomenclature for oligosaccharide fragmentation used throughout the mass spectrometry field is shown in Figure 1. Fragment ions that contain a non-reducing terminus are labeled with uppercase letters from the beginning of the alphabet (A, B, C), and those that contain the reducing end of the oligosaccharide or the aglycon are labeled with letters from the end of the alphabet (X, Y, Z); subscripts indicate the cleaved ions. The A and X ions are produced by cleavage across the glycosidic ring, and are labeled by assigning each ring bond a number and counting clockwise. Figure 1 shows examples for two cross-ring cleavage ions. Ions produced from cleavage of successive residues are labeled: Am, Bm, and Cm, with m ¼ 1 for the non-reducing end and Xn, Yn, and Zn, with n ¼ 1 for the reducing-end residue. Note that Y0 and Z0 refer to the fragmentation of the bond to the aglycon. When considering tandem MS of oligosaccharides, there are several options regarding the state of the precursor ion, the choice of which will dramatically influence the product-ion pattern, and thus the structural information, produced. For native oligosaccharides, the options in- clude protonated molecule ions, [M þ nH]nþ, deprotonated molecule ions, [M–nH]n, and natriated molecule ions, [M þ Na]þ. Permethylated and peracetylated derivatives are analyzed as either [M þ H]þ or [M þ Na]þ with MALDI, or as multiply charged ions with ESI. Table 1 shows the combination of ionization states and derivatives that have been studied in the literature. Many of the principles of carbohydrate fragmentation with tandem MS were developed with fast atom bombardment (FAB) ionization (also known as liquid secondary ion mass spectrometry), and this review takes those principle into consideration along with later developments. Note that metal-adducted ions, in which charge is carried by protons, will be denoted [M(X) þ H]þ. This nomenclature assumes the adduction of the metal cation with the displacement of a proton, and the positive ion is formed by the addition of a proton. It is understood that the sodium cation is paired with an acidic group. This nomenclature is used here and elsewhere (Costello, Juhasz, & Perreault, 1994; O’Connor, Mirgorodskaya, & Costello, 2002; Zaia & Costello, 2003) because it simplifies the notation of precursor and product ions, and facilitates the labeling in tandem mass spectra. Note that cationized ions, in which charge is carried by the metal, are given as [M þ mX]mnþ, where m ¼ the number of metal ions and n ¼ the cationic charge. Tandem MS of carbohydrates was first studied with FAB ionization, a technique that imposed severe constraints on the analysis. Neutral (containing no amino groups) and basic (containing hexosamine residues) carbohydrates ionize relatively poorly with FAB, and the tandem MS analysis was practically limited to molecules of <1000 Da. Acidic oligosaccharides (those containing Neu5Ac or sulfate) were observed to produce relatively strong ions with negative FAB (Egge & Peter-Katalinc, 1987). The ion signals were observed to be much stronger when the oligosaccharides or glycoconjugates were permethylated or peracetylated. The FAB ionization process was also energetic enough to produce glycosidic-bond and cross-ring cleavages—a phenomenon that was used to obtain structural information on molecules that had been purified to homogeneity (Dell et al., 1983a,b; Egge, Dell, & Von Nicolai, 1983; Carr & Reinhold, 1984). It was also observed that neutral gas collisions enhanced the yield of fragment ions generated from oligosaccharides (Carr et al., 1985). The development of tandem mass analyzers for TABLE 1. Ionization options for tandem MS of oligosaccharides FIGURE 1. Nomenclature for glycoconjugate product ions generated by tandem MS (Modified from Domon & Costello, 1988b). 164 MASS SPECTROMETRY OF OLIGOSACCHARIDES biological macromolecules allowed the selection of individual precursor ions and subsequent fragmentation, thus eliminating the need to purify oligosaccharides to homogeneity. Using FAB tandem MS, it became generally accepted to analyze sodium adducts of permethylated oligosaccharides as [M þ Na]þ ions—as will be discussed in detail below. The development of ESI greatly expanded the capabilities of the tandem MS of oligosaccharides by improving, relative to FAB, the strength of the ion signals produced from a given quantity of oligosaccharide. Although the permethylated oligosaccharides were observed to produce the strongest signals, the direct analysis of native oligosaccharides was possible for larger precursor ions than was possible with FAB. That increase in sensitivity was such that researchers had the choice to ionize native or permethylated oligosaccharides or glycoconjugates as protonated, deprotonated, or alkali-adducted ions. Because the information content of product-ion mass spectra depends to a large extent on the state of the precursor ion, it is important to examine the available information in this area. The following sections summarize the fragmentation behavior of oligosaccharides and oligosaccharide portions of glycoconjugates with regard to the state of the precursor ion so as to allow the reader to choose the appropriate analysis options. Recent work in this area is summarized, with regard to overall trends for carbohydrate fragmentation. Product-ion mass spectra of glycoconjugates are considerably more complicated than are those of peptides because of their branching structure. For such branching structures, fragmentation occurs from the non-reducing end of each antenna and from the reducing end to give rise to a higher complexity. The input of energy into the molecule by collision is most likely to break single bond glycosidic linkages. Using low energy CID, fragmentation of glycosidic linkages is most likely; fragmentation across the sugar rings is less so because two covalent bonds must be cleaved. It does occur, however, and provides important information on the location of substituents on branching monosaccharide residues. Research has shown that CID product-ion mass spectra provide information on the stereochemistry of individual sugar residues (Mueller et al., 1988), the linkage position (Laine et al., 1988), and branching structure (Carr et al., 1985; Domon & Costello, 1988a). Oligosaccharides that contain the same monosaccharides linked with a different branching structure often show distinct product-ion patterns because the steric environments differ between such isomers and result in different bond energies and ion abundances in product-ion mass spectra (Laine et al., 1988). Today, it is not possible to determine all of the linkages and branching patterns for a complex branched oligosaccharide. However, much work has been conducted in the past 15 years to improve the & degree to which the product-ion patterns generated from those molecules may be directly interpreted, as opposed to their being used as mass fingerprints, whose pattern reflects but does not predict the structure. C. Tandem MS of Native Oligosaccharide Molecular Ions 1. Protonated Ions The CID fragmentation patterns differ according to the oligosaccharide sequence, size, and class of subunits, as discussed below, and are used to differentiate subtle structural differences. CID fragmentation of [M þ H]þ ions generated from asialo glycoconjugates results in the observation of abundant Bm and Yn ions; those patterns are useful in assigning the oligosaccharides sequence (Domon & Costello, 1988b). Cleavage to the reducing side of HexNAc residues is favored, resulting in abundant fragment ions (Egge, Dell, & Von Nicolai, 1983). Therefore, such fragmentation is useful to define the presence of lactosamine (Gal-GlcNAc) disaccharides in the antennae of N-linked oligosaccharides. CID fragmentation of neutral glycoconjugates (gangliosides) results in the observation of abundant Bm and Yn ions, indicating that charge is not localized in those ions (Domon & Costello, 1988b). The abundances of ions produced from cross-ring cleavages is comparatively low for [M þ H]þ ions generated from that compound class. Those results are similar to the metastable fragmentation observed for those molecules in FAB MS (Egge & Peter-Katalinc, 1987; Gunnarson, 1987). When the protonated ions generated from small oligosaccharides are subjected to CID, a greater diversity of information is produced than that obtained from large oligosaccharides (Dell, 1990; Gillece-Castro & Burlingame, 1990). Specifically, glycosidic bond cleavages are more likely to predominate in product-ion mass spectra of large ions, with the result that less detailed information regarding the branching structure is obtained. Comparatively more subtle information, with regard to cross-ring cleavage ions, may thus be obtained for small ions. Those observations have led to efforts to increase the information content of product-ion mass spectra. Conditions during ESI MS of native oligosaccharides can be manipulated to favor the production of protonated, natriated, or ammoniated precursor ions (Duffin et al., 1992). Natriated oligosaccharides are produced in the presence of a low concentration of sodium acetate. Ammoniated adducts formed in the presence of ammonium acetate readily decay to protonated ions due to the energy required to desolvate ions during the electrospray process. Sialylated oligosaccharides were observed to produce abundant [M–H] ions in the negative-ion mode with ESI MS. The yield of glycosidic bond product ions in high-energy CID 165 & ZAIA was observed to be higher for protonated relative to natriated native oligosaccharide ions (Orlando, Bush, & Fenselau, 1990; Duffin et al., 1992). Cleavages to the reducing side of HexNAc residues are observed in abundance, particularly to those residues located on complex N-linked antennae. High-mannose oligosaccharides produce a series of product-ions from successive losses of hexose residues, resulting in a generally more uniform product ion pattern than observed for complex Nlinked sugars that contain HexNAc residues near the nonreducing termini. Thus, the pattern of CID product ions is useful to define the class of N-linked oligosaccharide (highmannose, hybrid, or complex). 2. Deprotonated Ions Neutral oligosaccharides have been observed to ionize relatively poorly relative to their basic (amino sugarcontaining) and acidic counterparts. Sialylated oligosaccharides are amenable to analysis as [M–H] ions, and are observed to produce product ions as abundant as those from metastable fragmentation with FAB ionization (Derappe et al., 1986; Egge & Peter-Katalinc, 1987). A series of molecules with Neu5Ac at the non-reducing terminus fragment to produce ions that almost exclusively contain the acidic group. This is contrasted with the fragmentation of an asialo ganglioside in which both reducing and non-reducing fragmentation results (Domon & Costello, 1988b). Fragmentation to the reducing side of HexNAc is facile in the CID of [M–H] ions. The importance of mobile protons in analyzing the fragmentation pattern becomes evident when comparing studies of deprotonated polysaccharides with their protonated forms. Interestingly, the high-energy CID production mass spectrum of the [M–H] ion generated from maltoheptaose (1,4-linked glucose heptamer) displays only 2,4Am ions (Gillece-Castro & Burlingame, 1990). By contrast, the [M–H] ion produced from the chitobiose core oligosaccharide, a branched (Man)2-Man-GlcNAcGlcNAc structure, displays abundant cross-ring cleavages with limited glycosidic bond fragmentation. The linear maltoheptaose spectrum displays a smooth decrease in ion abundances as the m/z values decrease, whereas the pattern generated from the chitobiose core is defined by a very abundant 2,4A3 ion. The discontinuous product-ion pattern of the chitobiose indicates branching. Protonated ions generated from the same molecules produce very abundant glycosidic bond cleavages with the very low abundance of cross-ring cleavages. The profusion of glycosidic bond cleavages of native oligosaccharides in CID product-ion mass spectra seems to be related to the presence of free hydroxyl groups. Hydroxylic hydrogen migration has been implicated in glycosidic-bond cleavages (Hofmeister, Zhou, & Leary, 166 1991), and the abundance of such ions appear to limit the abundance of cross-ring cleavage ions. In accordance, certain cross-ring cleavage ions, observed in the production mass spectra of permethylated oligosaccharides, have been observed to be absent from spectra acquired on native oligosaccharides (Harvey, Bateman, & Green, 1997). In protonated or alkali-cationized oligosaccharides, the presence of HexNAc residues places a charge on the amide nitrogen. The proximity of that charge to the reducing-side glycosidic oxygen evidently predisposes that bond to scission. Although a similar pattern is observed for deprotonated oligosaccharides, it is not observed for chondroitin sulfate glycosaminoglycans which consist of [UroA(1–3)GalNAcSulfate(1–4)]n, in the negative ESI mode (Zaia, McClellan, & Costello, 2001; McClellan et al., 2002). The CID production mass spectrum of a triply charged CS hexamer (n ¼ 3) results in abundant glycosidic bond cleavages to the reducing side of uronic acid residues but not to the reducing side of GalNAc residues. This difference is evidence that the glycosidic bond to the reducing side of GalNAc is significantly less labile than is the bond to the reducing side of UroA residues. This result is in marked contrast to the observations made for deprotonated oligosaccharides with FAB MS (Domon & Costello, 1988b). For further discussion on this issue, see Section VI.C.6.a. 3. Alkali and Alkaline Earth Adducted Ions The fragmentation of natriated native oligosaccharides by high-energy CID results in different product-ion patterns than are obtained from protonated ions (Orlando, Bush, & Fenselau, 1990). Specifically, the production of cross-ring cleavages is enhanced relative to the levels observed in the product-ion mass spectra of protonated molecular ions. Furthermore, the abundance of ions produced from cross-ring cleavages were reduced as the collision energy was lowered from 8 to 2 keV; that reduction indicates that the use of high-energy CID increases the information content of product-ion spectra. The Low-energy CID spectra of monolithiated disaccharides show abundant reducing end cleavages, but little differentiation between different linkages. Dilithiated disaccharides produce product-ion patterns that readily differentiate linkage isomers (Zhou, Ogden, & Leary, 1990). The dilithiated ions have lithium substitution for a hydroxyl hydrogen, and that substitution is believed to initiate the ring-cleavage rearrangements. Using [M(Li) þ Li]þ ions, it was possible to assign the linkages of Glc(1– 6)Glc(1–6)Glc(1–6)Glc versus the (1–4) linked isomer. PSD allows the analysis of product-ions that result from low-energy processes. PSD of natriated ions produced from native oligosaccharides resulted in an abundant series of glycosidic-bond and cross-ring cleavages. MASS SPECTROMETRY OF OLIGOSACCHARIDES Cleavages adjacent to HexNAc residues were facile, as observed for other dissociation techniques (Spengler et al., 1994). PSD analysis of natriated and protonated forms of Gal(b1-3)-GlcNAc(b1,3)-Gal(b1-3)-Glc-Bz (benzyl amino derivative) is instructive (Lemoine, Chirat, & Domon, 1996). The natriated form produces abundant Bm and Yn ions, indicating that charge is located without preference to either end of the molecule. The protonated form produces primarily Yn ions, indicating that the charge resides on the basic benzylamino group on the reducing terminus. The B2 ion is also observed from facile cleavage of the GlcNAc residue. PSD of a series of protonated N- and O-linked benzylamino structures was also shown to result in predominantly Yn ions. MALDI FTMS studies on alkali cationized native oligosaccharides have shown that the yield of fragmentation correlates with the degree of branching (Cancilla et al., 1996). Fragmentation yields were highest for oligomers with the least branching, and were inversely related to cation size, following the order Hþ>Liþ>Naþ>Kþ> Rbþ>Csþ. Mechanism of fragmentation of protonated ions is likely to be charge-induced, whereas that for cesiated ions is likely to be charge-remote. Charge-remote fragmentation requires more energy than charge-induced fragmentation, and the degree to which this occurs increases with increasing cation size. The effects of protons versus alkali metal cations may be rationalized based on differences in the coordination of the glycosidic oxygen. As shown in Figure 2, protonation is likely to be localized to the glycosidic oxygen—the most basic in the structure (Cancilla et al., 1996). Metal ions, on the other hand, can undergo coordination with several atoms simultaneously, resulting in less destabilization of the glycosidic bond. This result is consistent with a higher barrier to glycosidic-bond fragmentation for the alkali cationized ions. In addition to the glycosidic oxygen atom, acidic residues coordinate alkali metal cations, and this factor has & been investigated as a means for stabilization and linkage determination (Penn, Cancilla, & Lebrilla, 2000). CID product-ion mass spectra of sialylated milk oligosaccharides were acquired on [M(X) þ X]þ ions, where X ¼ Li, Na, K, or Cs, and showed that sialic acid residues are able to bind two cesium cations but only one lithium or sodium cation. It appears that small cations, with a high charge density, repel each other, making binding of two cations energetically unfavorable. Alkali-metal cationization and subsequent positiveion CID MS results in three possible fragmentation pathways, as shown in Figure 3 (Cancilla et al., 1999). The complex may dissociate to lose the metal cation (path A), leaving the oligosaccharides as a neutral, and producing no detailed information. The complex may undergo glycosidic bond cleavage (path B) with cation retention by one of the fragments. The complex may also undergo cross-ring cleavage (path C). The likelihood of dissociation of the cation increases with size, and is greatest for Csþ and least for Liþ and Naþ. Activation energies for glycosidic-bond cleavage have been determined to be lowest for Liþcoordinated oligosaccharides. As a result, use of this cation maximizes glycosidic bond fragmentation. The low energy barrier results in the observation of abundant fragment ions by MALDI MS, particularly for FT analyzers for which the ion lifetime is long. Glycosidic-bond cleavages are believed to be charge-induced, and Liþ appears to mimic protons by having high charge density and associating with glycosidic oxygen atoms. The use of Csþ is best for producing MALDI ions stable enough to be detected in instruments, such as the FTMS, with long ion lifetimes. That cation is large and interacts with several sites on the oligosaccharide to minimize the destabilization of the glycosidic bond (Tseng et al., 1997). Activation energy barriers have been studied with semiquantative methods, and have been found to be independent of alkali metal ions. Those cleavages appear to be charge-remote, occurring some distance from the charged site. Glycosidic-bond FIGURE 2. Fragmentation of (a) protonated and (b) alkali-cationized glycosidic bonds (Modified from Cancilla et al., 1996). 167 & ZAIA FIGURE 3. Three possible fragmentation pathways for metal-catio- nized oligosaccharides. Reprinted with permission of Cancilla et al. (1999). Copyright 1999 American Chemical Society. cleavages appear to be favored under conditions where a cation is associated with a glycosidic oxygen atom. Crossring cleavages occur under conditions where the cation binds simultaneously to several oxygen atoms and the degree of destabilization of glycosidic linkages is minimized. Analysis of protonated and natriated N-linked oligosaccharides by ESI Q-oTOF MS resulted in the conclusion that the latter produces the more-informative product-ion profiles (Harvey, 2000a). Cross-ring cleavages were observed to the reducing terminal GlcNAc residue and, at low abundances, to antenna residues. The abundance of crossring cleavage ions diminished with increasing number of HexNAc residues in the antennae; that result is consistent with the idea that cross-ring cleavages are most abundant in the absence of labile glycosidic bonds. During CID of an oligosaccharide molecular ion, the transfer of energy to rotational and vibrational modes competes with glycosidic-bond cleavage (Mendonca et al., 2003). The degree to which peralkylated oligosaccharides undergo glycosidic bond cleavage increases with the size of the alkyl group; i.e., methyl<ethyl<propyl<butyl< pentyl. The larger alkyl groups are less free to rotate about the glycosidic bonds, and therefore dissociate at a higher rate to relieve steric and torsional strain. Certain rotational states are inaccessible for large alkyl groups, so that transition states that lead to dissociation are ordered and give rise to reduced energies of activation. The smaller derivatives will have a less-ordered transition state and thus higher entropy of activation. The anomeric configuration of (1–4) linked disaccharides can also be distinguished by the CID of peralkylated protonated ions (Mendonca et al., 2003). Product ion-to-parent ion ratios are higher for the b-isomer (cellobiose) than the a-isomer (maltose) for all alkyl groups. The ratios reflect the differences in crowding 168 between a- and b-glycosidic linkages. The degree of discrimination is highest for the largest alkyl group tested (pentyl), reflecting the greater degree of conformational restriction. The anomeric differentiation in (1–6)-linked peralkyl derivatives is not observed due to the high conformational flexibility of this linkage. Unambiguous determination of linkages of increasingly large oligosaccharides requires the coordination with relatively more equivalents of metal ions, as shown with ESI (Fura & Leary, 1993). In addition to the amount, the choice of metal is important for large oligosaccharides. Alkaline earth elements (Mg2þ and Ca2þ) should, when coordinated with oligosaccharides, produce similar product-ion profiles as dilithiated compounds. Lithium has a high charge-to-radius ratio and is similar to Mg2þ in that both ions have a small radius and form stable compounds with small anions. Calcium ions, with a larger radius, are expected to coordinate more efficiently with branched oligosaccharides. Note that [M þ Na]þ and [M þ K]þ ions are isobaric with [M þ Mg H]þ and [M þ Ca–H]þ ions, respectively. Thus, the use of isotopically pure 26Mg and 44 Ca may be necessary to establish that an observed adduct is due to the divalent cation rather than to sodium or potassium. As the concentration of oligosaccharide decreases, the optimal metal-to-oligosaccharide ratio increases. For example, at 1.5 nmol/mL, the optimum ratio is 1:1, but at 12 pmol/mL, the ratio is 10:1 (Fura & Leary, 1993). Results of the CID of coordinated trisaccharides were much clearer for Ca2þ than for Mg2þ. Here, Ca2þ formed only a few well-defined product ions, whereas the latter produced complex patterns due to the presence of competing disaccharide pathways. The radius of Mg2þ (0.65 Å) is much smaller than that of Ca2þ (0.99 Å) and is thus expected to coordinate many more sites on the trisaccharide molecule. Binding of Ca2þ is more selective in its sites of coordination, and thus produces simpler product-ion patterns. Coordination with Mg2þ results in more abundant cross-ring cleavages than do two equivalents of Liþ due to the increased ability of the divalent ion to polarize and bind tightly. Tighter binding leads to more cross-ring cleavages. With larger oligosaccharides, Liþ induces product-ion patterns that are similar to those by Hþ, giving rise to only glycosidic bond cleavages (Fura & Leary, 1993). The abundance of natriated ions is maximized at high cone voltages (Harvey, 2000a)—conditions that also result in in-source fragmentation. Because of this factor, the precursor-ion pattern is not necessarily reflective of the glycan composition. With divalent metals (Ca2þ, Mg2þ, Co2þ, Cu2þ, Mn2þ), the dominant species formed from chloride salts were [M þ X]2þ (Harvey, 2001). The ability of divalent metal ions to ionize neutral oligosaccharides follows the order Ca2þ > Mg2þ > Mn2þ > Co2þ > Cu2þ. The metal-adducted oligosaccharides resulted in singly and doubly cationized product ions with abundances MASS SPECTROMETRY OF OLIGOSACCHARIDES highest for the Ca2þ ions. Because 1þ and 2þ product ions are present simultaneously, the CID profiles are relatively complex. The type of fragment ions observed for N-linked oligosaccharides are similar to those observed with natriated ions in that the only cross-ring cleavage ions observed in abundance were 0,2A and 2,4A of the reducingterminus GlcNAc residue. The primary advantage of divalent cations, the high relative abundance of the adducted ion, particularly for Ca2þ, is offset by the increased complexity of the CID spectra. At the time of this writing, Co2þ (Sible, Brimmer, & Leary, 1997) and Ca2þ (Harvey, 2001) are the most promising divalent metals for CID of native oligosaccharides. D. Tandem MS of Permethylated and Peracetylated Oligosaccharides The conversion of glycans to hydrophobic derivatives enhances their signal strengths regardless of the ionization technique used. The two most common derivatization procedures involve peracetylation (Bourne et al., 1949) and permethylation (Ciucanu & Kerek, 1984); both can be carried out in high yield (Dell, 1990). Permethylation has come to be the preferred method because it results in a smaller mass increase and a greater volatility (McNeil et al., 1982; Dell et al., 1983a,b). Peracetylated derivatives may be detected in the positive mode as [M þ Na]þ ions; CID fragmentation results in preferential cleavages to the reducing side of HexNAc residues (Dell et al., 1983). As with permethylated derivatives, [M þ H]þ ions from peracetylated oligosaccharides fragment to produce little multiple-bond fragmentation (Domon, Müller, & Richter, 1990). This result is an important advantage because the m/z values of such ions are, in many cases, not distinguishable from those of primary fragment ions generated from underivatized oligosaccharides. Therefore, internal fragment ions produce unique mass values in permethylated and peracetylated oligosaccharide product-ion mass spectra. As a consequence, linear and branched oligosaccharides are easily distinguished for peracetylated [M þ H]þ ions despite the lack of crossring cleavage ions (Domon, Müller, & Richter, 1990). Permethylation is often preferred over peracetylation; for example, for the oligosaccharides with free hexosamine amino groups that characterize the glycan core of glycoinositol phospholipid anchor. Those residues will be rendered indistinguishable from those that are acetylated if they are peracetylated. Permethylation produces a quaternary ammonium cation from the free amine groups that is differentiated by mass from the N-acetylhexosamine residues, and also gives rise to unique CID fragmentation (Baldwin et al., 1990). Permethylated oligosaccharides are most often ionized as [M þ Na]þ ions and produce reducing and non-reducing & terminal product ions with approximately equal abundances and preferential fragmentation to the reducing side of HexNAc residues (Egge, Dell, & Von Nicolai, 1983). Fragmentation of the substituents linked to the 3-position of HexNAc residues is also preferred (Egge & PeterKatalinc, 1987). In general, ions that correspond to Neu5Ac oxonium ions are abundant in fragmentation spectra of sialylated oligosaccharides and glycoconjugates, in some cases complicating the task of locating the position of substitution because ions with glycosidic bonds to those residues are in low abundance. Fragmentation of [M þ H]þ generated from permethylated oligosaccharides also produces useful CID production mass spectra. Cleavage to the reducing side of HexNAc is facile, and subsequent stages of MS are quite useful for differentiating isomers. The protonated molecule ions produce exclusively glycosidic-bond cleavage with lowenergy CID on a triple quadrupole instrument (Viseux, de Hoffmann, & Domon, 1997). Although multiple bond fragmentation is observed, permethylation allows the ions to be unambiguously identified on the basis of the exposed free hydroxyl groups. HexNAc residues substituted at position3 undergo a specific elimination that identifies that type of interglycosidic linkage. Tandem mass spectrometric analysis with MSn of permethylated N-linked oligosaccharides has been facilitated by removal of labile groups. The most labile groups are Neu5Ac and HexNAc residues for typical oligosaccharides, and subsequent stages of MS proved information on the non-reducing terminus and core structure that is not obtained in the MS2 profile (Weiskopf, Vouros, & Harvey, 1997, 1998; Reinhold & Sheeley, 1998; Sheeley & Reinhold, 1998; Viseux, de Hoffmann, & Domon, 1998). Exposure of core structures produces a fragment-ion pattern upon subsequent CID that appears to be independent of the history of formation of the ion. Figure 4 compares product ions of m/z 1143.6, (GlcNAc)2(Man)3, generated by MS4 of a sialylated biantennary structure and by MS3 of asialo biantennary structure NA2 (the specific fragmentation routes are shown in the figure). The structure of the non-reducing antennae may also be elucidated by MSn, producing linkage information based on the presence of cross-ring cleavages, and/or comparison to reference mass spectra (Reinhold & Sheeley, 1998; Sheeley & Reinhold, 1998; Weiskopf, Vouros, & Harvey, 1998). The protonated ions fragment at energies < natriated ions, and the product-ion patterns contain different features. Permethylated oligosaccharides may be desalted with reversed phase chromatography, resulting in their ionization as protonated species (Laine et al., 1988, 1991; Viseux, de Hoffmann, & Domon, 1997, 1998). For example, Figure 5 shows (a) the ESI mass spectrum of milk oligosaccharide lacto-N-tetraose [Gal(b1-3)GlcNAc(b13)Gal(b1-4)Glc), LNT], (b) MS2 of the protonated ion, 169 & ZAIA FIGURE 4. MSn fingerprinting of N-linked oligosaccharide core for facile determination of sugar branching and substitution: (a) MS4 of the glycan core of the disialo, biantennary human transferrin glycan, m/z 1143.5; (b) MS3 of the glycans core of the asialo, galactosylated biantennary oligosaccharide (NA2) from human fibrinogen, m/z 1143.7. Reprinted with permission from Weiskopf, Vouros, & Harvey (1998). Copyright 1998 American Chemical Society. and (c) MS2 of the natriated ion. The protonated ion fragments to produce fragments that contain the nonreducing terminus. The natriated ion, on the other hand, fragments to produce abundant Bm and Yn ions. The protonated ions undergo eliminative losses of substituents to the 3-position of GlcNAc to produce an Em ion. Those ions result from facile fragmentation of corresponding Bm ions, as shown in Figure 6, and are useful to identify the mass of the substituent on the 3-position. If the 3-position is unsubstituted, then the loss of a methoxy group is observed. If a monosaccharide residue is located on the 3-position, then the Em ion will reflect that loss. The Em ions were not observed for natriated ions. Fragmentation of protonated permethylated oligosaccharides, therefore, appears to have significant advantages over the natriated variants, by virtue of the specific losses that indicate the pattern of substitution of HexNAc residues. Unfortunately, those advantages are offset by other problems, such as internalresidue rearrangements (see Section II.I). An LC/MS approach has been developed for the online analysis of permethylated oligosaccharides with an ion trap (Delaney & Vouros, 2001). The released oligosaccharides were reductively aminated with 2-aminobenzamide to facilitate UV detection and permethylated. Ions were subjected to data-dependent MS2 as they eluted from a reversed phase HPLC column as sodium adducts. This 170 analysis entails the software-controlled detection of ions followed by isolation and CID of those ions that meet abundance or other exclusion criteria. The most abundant product ion was automatically selected for MS3. The HPLC-separation step carries the advantage that isomeric structures are likely to be separated to some degree. As in other QIT MSn analyses, the abundance of cross-ring cleavages are quite low. The data are, therefore, most useful as mass fingerprints, and the HPLC separation should, therefore, facilitate the building of a library of reference structures in a manner similar to that published (Viseux, de Hoffmann, & Domon, 1998; Tseng, Hedrick, & Lebrilla, 1999). E. Tandem MS of Reductively Aminated Carbohydrates The reducing-terminus aldehyde group of oligosaccharides is easily reacted with alkylamines. As shown in Figure 7, the amine forms a Schiff base with the aldehyde that cyclizes to form a glycosylamine. The Schiff base is usually reduced to form a secondary amine because this amine is more stable than a glycosylamine. Such a reductive amination of glycans provides several benefits for MS analysis. The derivative usually contains a chromophore to enhance its chromatographic detectability. Derivatives that MASS SPECTROMETRY OF OLIGOSACCHARIDES & FIGURE 6. Fragmentation of permethylated N-acetylhexosamine-con- taining oligosaccharide B ions, glycosylated in the 3-position (a) and in the 4-position (b). Reprinted with permission from Viseux, de Hoffmann, & Domon (1997). Copyright 1997 American Chemical Society. FIGURE 5. ESMS spectrum of permethylated LNT (a), and ESMS/MS spectra of protonated (b), and natriated (c) molecular species measured at collision offset voltages of 15 and 40 V, respectively. Reprinted with permission from Viseux, de Hoffmann, & Domon (1997). Copyright 1997 American Chemical Society. increase the hydrophobicity of the oligosaccharide will typically increase signal intensity obtained from any ionization technique (Wang et al., 1984; Carr et al., 1985; Gillece-Castro & Burlingame, 1990; Poulter & Burlingame, 1990; Harvey, 2000b). A number of amine groups have been used for reductive amination in conjunction with ESI CID MS, and their structures are shown in Figure 8 (Harvey, 2000b). The choice of the amine influences the ionization efficiency, and the effects are different with MALDI versus ESI, as shown in Figure 9. Although the more-hydrophobic derivatives tend to produce better ionization responses for both ionization modes, the 2-aminoacridone derivative produces a remarkable difference in response between the two ionization techniques. That derivative is widely used in fluorophore-assisted carbohydrate electrophoresis, and has been used in mass spectrometric studies (Okafo et al., 1997). It should be noted that the ESI measurements in Figure 9 were made at flow rates of 50–200 nL/min, where the droplet size is closer to high flow rate rather than nano flow. It is, therefore, possible that the less hydrophobic derivatives will produce stronger ion signals with nanospray, just as was observed with underivatized oligosaccharides (Bahr et al., 1997; Karas, Bahr, & Dulcks, 2000), resulting from the increased surface activity from smaller droplet size. The nature of the reductive amination derivative was found to have comparatively little effect on the CID profiles of natriated ions, indicating that the cation associates with the monosaccharide residues rather than with the derivative (Harvey, 2000b). The profiles were also similar to those of natriated native oligosaccharides. Crossring cleavages were present in low relative abundances, requiring extended summation times to achieve an adequate signal-to-noise ratio for proper identification. F. Discrimination of Monosaccharide Linkages High-energy CID of cationized permethylated oligosaccharides furnishes information that identifies (2–3) versus (2–6) linked Neu5Ac residues, as well as (1–3) versus (1– 4) linked hexose on a HexNAc residue (Lemoine et al., 1991). Discrimination between Neu5Ac linkages is based on the different affinities for sodium cations that gives rise 171 & ZAIA FIGURE 7. Reductive amination of oligosaccharides. to dramatically different product-ion patterns. Discrimination between hexose linkages on HexNAc residues is based on the facile elimination of the (1–3) linked group (Laine et al., 1988; Laine, 1989). The abundance of structurally useful cross-ring cleavage ions have been shown to depend on the collision energy and the collision gas (Lemoine et al., 1993). The low-energy CID of the [M þ Na]þ ion generated from permethylated oligosaccharides produces primarily single-bond glycosidic-bond cleavage. At high energy (4 keV), double-bond cross-ring cleavages predominated in the product-ion mass spectrum, with single-bond glycosidic bond cleavages in low relative abundances. The use of argon rather than helium as the collision gas also results in more efficient fragmentation of the precursor-ion, and in an enhancement of the abundance of cross-ring cleavage FIGURE 8. Structures of common amines used with reductive amina- tion. Reprinted by permission of Elsevier from Harvey (2000b). 172 ions. The results are consistent with the conclusion that the cross-ring cleavage ions arise through charge-remote fragmentation processes (Adams, 1990). Cationization at different glycosidic linkages may give rise to a set of charge-remote fragmentation reactions, the sum of which is the product-ion spectrum acquired at high collision energy (Lemoine et al., 1993). The observations of characteristic peaks that differentiate linkage were originally made by several groups, as summarized below. 1. Low-energy FAB CID mass spectra acquired for [M þ H]þ ions generated from three trisaccharides, Fuc(a1-X)GlcNAc(b1-3)Gal(b1-O-methyl), where X ¼ 3, 4, or 6, clearly show differences in the abundance for Y2 ions at m/z 398 and B2 at m/z 350. The resulting m/z 350/398 ratio indicates the Fuc linkage position in the order of (1–6) > (1–4) > (1–3) (Laine et al., 1988). 2. Direct stereochemical assignment of monosaccharide units was made with low-energy CID of the glycosylated antibiotic papulacandin (Müller et al., 1988). FIGURE 9. Comparison of MALDI (solid bars) and ESI (shaded bars) signal strengths from the high mannose N-linked glycan (GlcNAc)2 (Man)5 derivatized by reductive amination with each of eight different amines. Reprinted by permission of Elsevier from Harvey (2000b). MASS SPECTROMETRY OF OLIGOSACCHARIDES FIGURE 10. Structures of peracetylated monosaccharide oxonium ions, m/z 331. Chemical ionization was used to produce [M þ H]þ ions for the acetylated molecule. The peracetylated monosaccharide oxonium ion (see Fig. 10 for structures) was generated in the source, and was subjected to isolation and CID fragmentation. The CID profile of that ion was compared to the oxonium ion at the same & m/z value, generated from acetylated methyl glycosides of Gal, Glc, and Man, respectively. The spectral pattern for the acetylated Gal oxonium ion is easily distinguished from those of Glc and Man, based on the high abundance of the product ion at m/z 127. The profiles of acetylated Glc and Man are not distinguishable. 3. Low-energy CID product-ion mass spectra generated for peracetylated disaccharide oxonium ions generated from Glc(b1-X)Glc, where X ¼ 2, 3, 4, and 6 (Domon, Müller, & Richter, 1989), show distinct patterns of abundances, for product ions at m/z 109, 169, 211, 271, 331, 457, 559, and 619. In an attempt to make objective distinctions among these spectral fingerprints, the authors use similarity values that reflect vectors in ndimensional space, where n is the number of m/z values. FIGURE 11. Structures of peracetylated lacto-N-fucopentaoses, LNF-I, LNF-II, and LNF-III, [M þ H]þ ¼ m/z 1484. The high-energy CID product ions diagnostic of each isomer are shown. 173 & ZAIA 4. Low-energy CID of [M þ H]þ ions clearly distinguishes peracetylated isomeric lacto-N-fucopentaoses (LNF, see Fig. 11 for structures) (Domon, Müller, & Richter, 1990). The CID profile of the linear LNF-I structure is distinguished from the branched structures by the presence of a B2 ion at m/z 561. The branched structures are distinguished from each other by the presence of En ions (see Section II.D) at m/z 500 (from LNF-II) and 558 (from LNF-III), formed by secondary fragmentation of the branched B2 ion to lose the substituent on the 3-position of the GlcNAc residue. 5. Distinct product-ion profiles were obtained from isomeric glucose disaccharides with negative ion B/E linked-scan product-ion MS (Garozzo et al., 1990). Specifically, although the (b1–6) linkage is characterized by abundant product ions at m/z 251 (90), 281 (60), 311 (30) and 323 (18), only m/z 281 is abundant in the profile of the (b1–4) linkage (the loss from the precursor ion is indicated in parentheses). The (b1–3) linkage displays abundant ions at m/z 161 (180) and 179 (162) that are absent or of very low abundance for the other isomers. The (b1–2) linkage is characterized by an abundant ion at m/z 263 (78). Those losses can be used to determine linkages in the production profiles generated from oligosaccharides. 6. Infrared MALDI desorption of isomeric disaccharides results in distinctive fragmentation to the reducing ring that is useful in assigning linkage (Spengler, Dolce, & Cotter, 1990). As observed in the negative mode, above, losses are observed that characterize each linkage. 7. As mentioned in Section II.C.3, low-energy CID MS of [M(Li) þ Li]þ ions generated from Glc(b1-X)Glc, where X ¼ 4, 6, results in distinct product-ion patterns from cross-ring cleavages (Zhou, Ogden, & Leary, 1990). Specifically, the (1–6) linked disaccharide fragments to produce ions at m/z 235, 265, and 295 from cross-ring cleavage of the reducing-end residue. The m/z 265 residue is absent in the profile of the (1–4) linked disaccharide. It was also shown that linkage patterns in linear glucose tetramers could be differ- entiated based on those characteristic cross-ring cleavage processes. 8. The formation of cross-ring cleavages to the reducingend residue with low-energy CID are dependent on linkage (Hofmeister, Zhou, & Leary, 1991). The disaccharide linkage isomers gentiobiose (b1,6), laminarbiose (b1,3), and sophorose (b1,2) are clearly differentiated based on characteristic patterns of the abundance of product ions at m/z 331 (–H2O), 289 (– C2H4O2), 259 (–C3H6O3), 229 (–C4H8O4), 187 (–C6H10O5), and 169 (–C6H12O6), as indicated in Figure 12. 9. High-energy CID product-ion mass spectra of sodiumducted methylated oligosaccharides has been shown to discriminate between (2–3) and (2–6) linkages of Neu5Ac (Lemoine et al., 1991), see Section V.D. High-energy CID of permethylated Neu5Ac(a2-3) Gal(b1–3)GlcNAc(b1–3) Gal(b1–4)Glc and Neu5Ac (a2–6)Gal(b1–4)GlcNAc(b1–3)Gal(b1–4)Glc, respectively, showed differences in product-ion pattern that is attributable to Neu5Ac linkage. Specifically, the (2–3) linked isomer fragmented to form abundant ions from cleavage of the glycosidic bond adjacent to the Neu5Ac residue, corresponding to C1 (m/z 356) and Z4 (m/z 864). Both ions are absent in the spectrum of the Neu5Ac (2–6) linked isomer. Comparison of low- and high-energy CID of protonated ions generated from glycoalkaloids indicates that production patterns are strongly dependent on the collision energy (Claeys et al., 1996). The abundance of the ions that result from Yn glycosidic bond cleavages are high with lowenergy CID, whereas those that result from cross-ring cleavages, primarily 1,5Xn, are high with high-energy CID. The high-energy mass spectra are more complex than those acquired at low energy, but contain the structurally valuable cross-ring cleavages. High-energy CID occurs on a much shorter time-scale than low-energy CID, and is much more likely to produce cross-ring cleavages from a charge-remote mechanism. By contrast, low-energy CID occurs on a longer time-scale, and involves multiple FIGURE 12. Fragmentation of the [M þ Li]þ ion generated from gentiobiose, (a) cross-ring cleavages, (b) glycosidic bond cleavages (Modified from Hofmeister, Zhou, & Leary, 1991). 174 MASS SPECTROMETRY OF OLIGOSACCHARIDES collisions with gas molecules, resulting in cleavage only to the most labile bonds. The question remains regarding how best to produce cross-ring cleavages while minimizing the extent to which multiple bond fragmentation occurs. Lowenergy CID instruments are by far the most readily available at the time of this writing. High- and low-energy CID of high-mannose N-linked oligosaccharides are compared in Figures 13 and 14, respectively. The high-energy CID data show nearly complete series of glycosidic-bond and cross-ring cleavages, including those produced from cleavage of the branched mannose residues (A4 and X3 ions). The most abundant cross-ring cleavage ions observed in the low-energy CID mass spectra of high mannose N-linked oligosaccharides (Fig. 14) are those that occur to the reducing terminal GlcNAc residue. Although other An ions are observed, the abundances are significantly lower than the cross-ring cleavage ions in the high-energy CID product ion mass spectra. Those results demonstrate that high-energy CID yields significantly more informative mass spectra than low-energy data. At this time, however, there are no commercially available instruments that can practically accomplish high-energy CID on biologically relevant sample quantities (low picomol quantities). Magnetic-sector instruments offer high-energy CID, but scan far too slowly & to be of practical use. MALDI TOF/TOF instruments are available, and theoretically produce high-energy CID. Low-energy product ions are often observed, however, due to metastable fragmentation (Yergey et al., 2002). The development of high-pressure MALDI sources may allow high-energy CID data to be obtained for oligosaccharides. For that purpose, it would also be desirable to develop an ESI source for the TOF/TOF analyzer. Although protonated oligosaccharides fragment at lower energies, natriated ions produce a higher yield of cross-ring cleavage ions (Orlando, Bush, & Fenselau, 1990). Highenergy CID mass spectra were acquired on natriated ions generated from MALDI, and were analyzed with a hybrid magnetic sector orthogonal injection time-of-flight analyzer (Bordoli et al., 1994; Bateman et al., 1995). The majority of the product ions observed in the tandem mass spectra resulted from remote-site fragmentation. Highmannose N-linked sugars (Man9) produce a complete series of Bm, Yn, and 1,5Xn ions, resulting in a highly complex product-ion pattern (Harvey, Bateman, & Green, 1997). The abundances of multiple bond fragment ions are low, and it appears that the product-ion mass spectra are valuable. Because the spectra were acquired on native molecules, the identities of some ions cannot be determined due to the possibilities of multiple bond fragmenta- FIGURE 13. High-energy CID mass spectrum of the high-mannose N-linked sugar Man9GlcNAc2. The ion labeled Man4 (m/z 671.3) is an internal fragment ion. Reprinted with permission from Harvey, Bateman, & Green (1997)). Copyright 1997 John Wiley & Sons Limited. 175 & ZAIA FIGURE 14. Low-energy CID of high-mannose N-linked oligosaccharides. Reprinted with permission from Harvey (2000a). Copyright 1997 John Wiley & Sons Limited. tion. Natriated native complex N-linked oligosaccharides produce abundant Bm ions from cleavage to the reducing side of HexNAc residues, especially near the non-reducing termini. Generally, the appearances of those mass spectra were indicative of the oligosaccharide structures as fingerprints, but were not amenable to direct interpretation. Although the abundances of cross-ring cleavage ions in the MS2 stage are typically low with low-energy CID, the use of higher-order MSn experiments enables more detailed information to be produced on oligosaccharides. The MS2 stage often serves to remove the most labile saccharide substituents from complex oligosaccharide ions to allow the selection and subsequent fragmentation of less-labile portions of the molecule. Cross-ring cleavages are in greater abundances in those subsequent MS stages, allowing more detailed information on the nature of underlying glycosidic linkages to be obtained (Weiskopf, Vouros, & Harvey, 1998). For complex N-linked oligosaccharides, the removal of antennae and labile Fuc and Neu5Ac residues permits the identification of the core pentasaccharide structure. Thus, multiple MS stages may be used to expose a common core structure exposed from different oligosaccharide molecules, resulting in identical product-ion patterns (Viseux, de Hoffmann, & Domon, 1998; Weiskopf, Vouros, & Harvey, 1998). Because of these observations of substructural fingerprints, several groups have proposed to use multistage MS to virtually degrade complex oligosaccharides to common core structures; the spectra could be 176 searched against a structural library (Viseux, de Hoffmann, & Domon, 1998; Tseng, Hedrick, & Lebrilla, 1999) or interpreted with a computer program (Gaucher, Morrow, & Leary, 2000; Ethier et al., 2002). Studies on lithium- and sodium-cationized disaccharides have demonstrated conclusively that low-energy CID results in glycosidic-bond and cross-ring cleavages (Hofmeister, Zhou, & Leary, 1991; Asam & Glish, 1997). Patterns of cross-ring cleavage ions are linkage-dependent, as shown for lithium-cationized (1–2), (1–3), (1–4), and (1–6) linked glucose disaccharides, as shown in Table 2 (Asam & Glish, 1997). The linkages in glucose oligomers could also be determined, as shown in Figure 15 for MS2 and MS3 data collected on lithium-cationized trimers of isomaltose [Glc(a1–6)Glc(a1–6)Glc] and panose [Glc(a1–6)Glc(a1–4)Glc] (Asam & Glish, 1997). The MS2 data (panels a and c) show product-ion patterns at m/z 229, 259, and 289 that are consistent with (1–6) linkages, referring to Table 2, for both isomers. The non-reducing end linkage is indicated by ions at m/z 392, 422, and 452 that correspond to the same cross-ring cleavages with a hexose residue attached. Multistage MS of cobalt-coordinated milk oligosaccharides has been used to determine precise linkage positions (Konig & Leary, 1998). This determination was shown for the lacto-N-fucopentaoses (LNFP) I, II, III, and V, that were all of composition (Gal)2(Glc)(GlcNAc)Fuc. During lowenergy CID, the metal-coordinated oligosaccharide ion begins to dissociate with specific cross-ring cleavages from MASS SPECTROMETRY OF OLIGOSACCHARIDES & TABLE 2. Lithium-cationized disaccharide MS/MS product ions and abundances [reprinted by permission of Elsevier from Asam & Glish (1997)] a Intensity relative to base peak. Cross-ring cleavage ion types in Domon–Costello nomenclature. c Galactosyl(1 ! 4)glucose, all others glucosyl(1 ! X)glucose. b FIGURE 15. (a) MS/MS spectrum of lithium-cationized isomaltotriose, (b) MS3 spectrum of the C2 ion of lithium-cationized isomaltotriose, (c) MS/MS spectrum of lithium-cationized panose, and (d) MS3 spectrum of the C2 ion of lithium-cationized panose. Note that the circular symbols in this figure depict MSn transitions, and do not refer to monosaccharide residues. Reprinted by permission of Elsevier from Asam & Glish (1997). 177 & ZAIA the reducing end, followed by non-reducing end cleavages (Zhou, Ogden, & Leary, 1990; Hofmeister, Zhou, & Leary, 1991). Linkages are indicated by the presence of specific neutral losses in the MS2 profiles, as shown in Table 3. Multistage MS data were acquired on LNFP III and LNFP V (Fig. 16), differing in the position of fucosylation and the Gal-GlcNAc linkage (see figure for structures) (Konig & Leary, 1998). The results are significant in that cross-ring cleavages are produced in the MS2 stage despite the presence of GlcNAc and Fuc residues; both are known to be labile. For each stage of MS, cross-ring cleavages, expressed as losses from the precursor ion, indicated the linkage position of the reducing-terminus residue. Selection of the Cm ions in subsequent stages of MS allows other linkages to be determined, as shown for the figure insets. The MS5 product-ion spectrum of LNFP III corresponds to the fragmentation of Gal(1–4)GlcNAc (Fig. 16d), in which an ion is observed at m/z 280, which corresponds to the loss of 161 from the precursor ion. The MS5 product-ion profile of LNFP V corresponds to fragmentation of Gal(1– 3)GlcNAc (Fig. 16i), and features an ion at m/z 279, produced by loss of 162 from the precursor ion. Convincing evidence from stable isotope-labeling shows that its formation occurs by a mechanism that is distinct from the m/z 280 ion in Figure 16d (Konig & Leary, 1998), and the cobalt complexes also differentiate GalGlcNAc linkages. Although the cobalt complexes do not provide direct evidence for the fucose linkages, dissociation of [M–H] ions from the same solutions provided this information. The usefulness of CID of cobalt complexes, however, is limited by two factors: (1) the relatively high m/z of the precursor ion, and (2) the difficulty in obtaining MS5 and subsequent fragmentation steps to obtain the linkage information on larger oligosaccharides. To get around the fragmentation efficiency problem, it has been possible to produce the necessary Cm ions in a single stage of MS2 (Konig & Leary, 1998). The MS3 experiments on each Cm ion provided the same structural information as in the MSn TABLE 3. MS2 results obtained from CID experiments of [M þ Co–H]þ for the indicated disaccharides with specific neutral losses depending on their linkage position [reprinted by permission of Elsevier from Konig & Leary (1998)] 178 experiments shown above. That approach will be useful to minimize sample consumption by reducing the number of ion-isolation steps necessary to obtain the linkage information. Transition-metal ligand attachment has been used as a derivatization method to improve oligosaccharide ionization efficiency (Gaucher & Leary, 1998; Desaire & Leary, 1999a,b; Gaucher, Pedersen, & Leary, 1999). Accordingly, the synthesis of divalent metal-coordinated N-glycosides has been shown to stabilize sialic residues and to provide determination of linkages (Leavell & Leary, 2001). The required N-glycosides are easily synthesized with diethylenetriamine, mixed with metal salts, and diluted prior to ESI MS analysis (Gaucher, Pedersen, & Leary, 1999). The structure of the complexes in which the metal is coordinated to the diethyenetriamine nitrogens at the reducing terminus is shown in Figure 17. Losses of sialic acid were dramatically reduced by transition-metal complexation in the order of the Irving-Williams series: CoII< NiII<CuII>ZnII. As shown in Figure 18, the pattern of neutral losses from metal-ligated precursor ions differ for (2–3) and (2–6) linked sialic acid linkages. One problem with the methodology is that the metallation reaction results in several species for each trisaccharide, including protonated, natriated, and transitionmetallated ions, giving rise to a complex mass profile. Also, the stabilization of sialic acid residues is believed to involve deprotonation of the carboxyl group, and it is, therefore, not clear how large a molecule will work with this approach because non-reducing terminal Neu5Ac residues will be distant from the derivatized reducing terminus. G. Gas-Phase Degradation of Oligosaccharides Although permethylated oligosaccharides ionize readily as sodium adducts, protonated ions may be observed by dissolving samples in ammonium acetate (Viseux, de Hoffmann, & Domon, 1998). Alternatively, permethylated oligosaccharides may be desalted and/or fractionated with reversed-phase chromatography or solid-phase extraction. Multistage tandem MS of [M þ H]þ ions generated from two permethylated isomeric milk oligosaccharides that differ in structure by the linkage of a single GlcNAc residue were observed, as shown in Figure 19. The data show the production of distinct patterns, primarily due to the E2 ion that results from facile elimination of the substituent on the 3-position of GlcNAc residues. The LNT oligosaccharide contains a Gal(1–3)GlcNAc linkage. Loss of the Gal residue occurs through E2 fragmentation to form an ion at m/z 228, which is not detected in Figure 19a,b due to the fact that it is below the ion-trap detection window. That ion is detected in the MS3 stage acquired by selecting and MASS SPECTROMETRY OF OLIGOSACCHARIDES & FIGURE 16. (a–e) MSn of LNFP III; (f–j) MSn of LNF PV. Reprinted by permission of Elsevier from Konig & Leary (1998). (Continued on next page.) 179 & ZAIA FIGURE 16. (Continued) fragmenting the B2 ion, as shown in Figure 19c,d. LNnT, containing Gal(1–4)GlcNAc, fragments to produce an abundant E2 ion at m/z 432 from the elimination of the methoxy group on GlcNAc position 3. Those E2 ions are shown in MS3 product-ion mass spectra acquired by selecting and fragmenting the B2 ions for either isomer (lower panels). In further work, it was shown that the FIGURE 17. Structure of the Mn(II) N-glycoside complex. Reprinted by permission of Elsevier from Leavell & Leary (2001). 180 pattern produced from the fragmentation of the B2 ion is the same, regardless of the history of formation of the ion. Specifically, 964 ! B2 ! produces the same pattern as 964 ! B3 ! B2 ! for a given isomer. Therefore, mass spectrometric fragments generated from large oligosaccharides may be subjected to CID, and the resulting product ion mass spectra compared directly with those of known compounds, such as those in Figure 19. Thus, it is possible to build a series of reference spectra to allow a comparison with unknown oligosaccharides to facilitate interpretation. It is clear that disaccharide reference product-ion spectra allow the differentiation of (1–3), (1–4), and (1–6) Hex-HexNAc linkages (Viseux, de Hoffmann, & Domon, 1998). The same approach was shown to be useful for the differentiation of fucosylation linkage patterns in milk oligosaccharides. The results are not as clear, however, for sialylated oligosaccharides. The MS/MS product-ion profiles of Neu5Ac(2–3)Gal(1–3)GlcNAc and Neu5Ac (2–6)Gal(1–3)GlcNAc were used as references for the MASS SPECTROMETRY OF OLIGOSACCHARIDES & FIGURE 18. MS3 of (a) [Co(II)/dien/a(2–3)-sialyllactose–H]þ and (b) [Co(II)/dien/a(2–6)-sialyllac- tose–H]þ. Reprinted by permission of Elsevier from Leavell & Leary (2001). determination of sialic acid linkages of milk oligosaccharides. As shown in Table 4, it was observed that (2–6) linked oligosaccharide LST-c produced a pattern that matched that of 6-SLN, a (2–6) linked compound used as a reference; however, the pattern for (2–3) linked oligosaccharide LST-a did not match the corresponding reference 3-SLN. A more reliable alternative was to hydrolyze the permethylated sialic residues under acid conditions, and to perdeuteromethylate the exposed hydroxyl groups. The resulting spectra may be directly compared to those of asialo reference compounds; the deuterium mass-shifts indicate the position of the sialylated residues. H. Computer-Based Approaches for Interpretation of Oligosaccharide Product-Ion Mass Spectra An automated computer program has been described that calculates theoretical PSD mass spectra from different 181 & ZAIA FIGURE 19. ESMSn of permethylated tetrasaccharides: MS2 spectra of [M þ H]þ ions at m/z 904 of LNT (a) and LNnT (b) (collision energy 18%); MS3 spectra of B2 fragments at m/z 464 of LNT (c) and LNnT (d) (collision energy 15%). Substructures derived from oligosaccharides are depicted with black symbols. Reprinted with permission from Viseux, de Hoffmann, & Domon (1998). Copyright 1998 American Chemical Society. TABLE 4. Intensities of fragment ions generated from permethylated sialylated subunits [reprinted with permission from Viseux, de Hoffmann, & Domon (1998), Copyright 1998 American Chemical Society] 182 MASS SPECTROMETRY OF OLIGOSACCHARIDES pyridylamidated N-linked oligosaccharide structures (Mizuno et al., 1999). The computer program takes into account the following fragmentation processes: (1) monosaccharide residue loss from the non-reducing termini; (2) subsequent monosaccharide residue losses from the aforementioned ions, generating a Yn ion series; (3) the complementary Bm ions; (4) formation of internal fragment ions from loss of pyridylamidated residues from the Yn ions; (5) formation of internal fragments from Bm ions; and (6) formation of Zn, as well as the complementary Cm ions. The program assigns the monosaccharide compositions of the losses observed from the precursor ion in acquired PSD mass spectra, and thereby provides a useful interpretation tool. Simulated mass spectra are calculated for candidate structures that may be manually matched against the observed data. The program does not appear to differentiate the PSD mass spectra calculated for branching isomers. A computer-based approach for interpretation of native oligosaccharide MSn data has been published (Gaucher, Morrow, & Leary, 2000). That program, known as STAT, calculates all possible monosaccharide compositions for a given precursor-ion and those for observed product-ions. The output lists all possible structures that may fit the precursor-ion mass and product-ion pattern. The program is useful for oligosaccharides that contain up to 10 residues, and a long list of possible structures is generated from the input data. A scoring system ranks the structures in terms of the degree to which they match the data, and the correct structure was found to be high-ranked. The number of structural possibilities may be narrowed with methylation analysis and/or biological data that eliminate structures because they lack certain core structures. The algorithm uses m/z values, and does not yet incorporate abundance information. By incorporating abundances, it may be possible to expand the discrimination of the program. Recent work has built on the STATalgorithm as applied to the interpretation of product-ion mass spectra of Nlinked oligosaccharides (Ethier et al., 2002). In that case, sugars are derivatized with phenyl-3-methyl-5-pyrazolone (PMP) (Honda et al., 1989; Saba et al., 1999), and tandem mass spectra were acquired with a Q-oTOF instrument. The product-ion patterns of protonated precursor ions generated by MALDI were dominated by Yn ions (Saba et al., 1999). I. Internal Residue Loss Rearrangements of Oligosaccharide Ions During CID High-energy CID of protonated native oligosaccharides that contain (1–2) linkages showed an unexpected loss of 162 u from the precursor-ion that could only be explained by loss of an internal residue (Kovacik et al., 1995). The & internal residue-loss ion does not appear in high-energy CID mass spectra of natriated ions or of protonated peracetylated ions. Protonated native and protonated permethylated oligosaccharides of (1–6) Gal oligomers have both been observed to produce abundant internal-residue losses from certain Yn ions (Brüll et al., 1997). Although a- and b-linked internal residues may both be lost, peracetylated derivatives did not undergo the rearrangement. In further work, protonated, natriated, and deprotonated ions produced from native and permethylated oligosaccharides were analyzed by high- and low-energy CID MS/MS (Brüll et al., 1998). Because natriated and deprotonated ions both failed to undergo internal residue losses in either high- or low-energy modes of fragmentation, it appears that protons are necessary to trigger the rearrangements. The rearrangements occur for a number of linkages and monosaccharide compositions, including HexNAc-containing oligosaccharides. Protonated native sialyl Lewis tetrasaccharide alkyl glycosides have been observed during low-energy CID to undergo internal-residue rearrangements (also described as sequence isomerization) through a migration of fucose residues toward the non-reducing terminal sialic acid residue (Ernst, Müller, & Richter, 1997). Those conclusions are based on the observation of fucosylated Bm ions that are not expected, based on the well-characterized known structures. The proposed mechanism for fucose migration is shown in Figure 20. Analysis of the highest Bm ion (B3 for sialyl Lewis structures) with MS3 resulted in much lower abundances of rearrangement ions. Mass spectrometric selection of a Bm ions corresponds to the removal of the aglycon alkyl group, and seems to favor better charge fixation, reduced proton mobility, and reduced rearrangement. Protonated ions produced from reductively aminated oligosaccharides undergo intramolecular fucose rearrangements towards the derivative (Harvey et al., 2000). The rearrangement is seen in [M þ H]þ and [M þ 2H]2þ ions, but is absent in [M þ Na]þ precursor ions. The fact that rearrangement is only observed for protonated ions is consistent with a charge-induced mechanism, and a proposal that involves protonation of the reducing terminal derivative is shown in Figure 21. The presence of rearrangements that isomerize the sequences in protonated oligosaccharide tandem mass spectra may lead to erroneous conclusions regarding the presence of more than one isomer in a mixture. The use of alkali or other metals is therefore advised when analyzing unknown oligosaccharides structures. It may be feasible to select appropriate ions, such as Bm, to minimize the extent of rearrangement (Ernst, Müller, & Richter, 1997), but this selection requires further investigation. Because lithiated oligosaccharides produce product-ion patterns that more closely resemble those produced from a 183 & ZAIA FIGURE 20. Proposed migration of (incipient) fucosyl C1 carbenium ion towards sialyl residue to yielding irregular m/z 438 and 600 ions (paths 1 and 2, respectively) via sequence-rearranged [M þ H]þ intermediate. Reprinted with permission of Elsevier from Ernst, Müller, & Richter (1997). protonated ions (Fura & Leary, 1993), it is not clear whether the use of this metal will prevent internal-residue rearrangement. Until further investigation has been made, it seems prudent to analyze natriated oligosaccharides to avoid rearrangements. J. Conclusions At the time of this writing, the most promising means to produce cross-ring cleavages by low-energy CID MS for carbohydrate oligomers is through the use of transition- FIGURE 21. Proposed mechanism for fucose migration during the fragmentation of 2-AB derivatized 20 FL. Reprinted from Harvey et al. (2002). Copyright 2002 American Chemical Society. 184 MASS SPECTROMETRY OF OLIGOSACCHARIDES metal complexes in combination with MSn. It may be possible to add appropriate metals to HPLC mobile phases either pre- or post-column and to detect complexes that would facilitate sequential identification of residues by MSn (Kohler & Leary, 1995; Konig & Leary, 1998). The observation that metals bind preferentially to alditols (Tseng, Hedrick, & Lebrilla, 1999) bears further investigation in that context. The extent to which metal complexation and MSn will prove valuable depends largely on the compound class. Linear oligosaccharides such as the glycosaminoglycans and those with a low degree of branching such as milk oligosaccharides and some O-linked sugars are likely to produce useful results with this approach. Highly branched N-linked sugars are likely to be too complex to produce data that can be directly interpreted. The value of tandem MS of N-linked sugars appears to be in classifying the type; i.e., high-mannose, hybrid, or complex, and the degree of branching. The spectra cannot be interpreted de novo because of the isomeric antenna structures and the overall complexity. It is also possible to make conclusions about the relative degree of branching based on the appearance of the product-ion profiles. The data do, however, appear to serve as a fingerprint for the N-linked structures. In that light, it would be valuable to construct libraries of MSn data on known N-linked structures, defining those that can be differentiated by mass spectrometric degradation. As detailed in this section, this work has already begun. Another development that may further the analysis of oligosaccharides is the MALDI TOF/TOF tandem mass spectrometer. That instrument is theoretically able to produce cross-ring cleavages, and may thus increase the information content of oligosaccharide mass spectra by increasing the abundances of cross-ring cleavages. The ions must be produced without metastable fragmentation, however, or else all of the ion current will go to more labile glycosidic bond cleavages. Because metastable fragmentation has been a problem with MALDI TOF/TOF of peptides (Yergey et al., 2002), carbohydrate mass spectrometrists are practically limited to low-energy CID at this time. The approaches for mass spectrometric analysis of carbohydrates are summarized in Figure 22. If the released glycans are available in quantities of 5 mg or greater, then permethylation is recommended. The procedure is quite robust, as evidenced by its wide use, and it increases the ionization responses from ESI and MALDI. In addition, tandem mass spectra of permethylated carbohydrates are more informative than those of native molecules because it is possible to identify cleaved glycosidic bonds by virtue of the free hydroxyl group created. This factor also allows one to identify internal fragment ions because they will not be isobaric with single-bond cleavage ions. The permethylated carbohydrates can be analyzed directly with ESI or MALDI, to obtain semi-quantitative responses, or fractio- & FIGURE 22. Analytical pathways for mass spectrometric analysis of carbohydrates. nated with reversed phase HPLC to limit mixture complexity. The most fruitful approach to CID is to build an understanding of the fragmentation of a library of known structures by acquiring MSn data. The MSn profiles of unknowns can be compared to that library to facilitate interpretation. In many cases, the quantities of released oligosaccharides will be below the level that is fruitful to attempt permethylation. This limit is primarily due to losses associated with the comparatively large solution volumes necessary during the extraction step that eliminates permethylation as a viable option for analysis of glycans released from SDS–PAGE gel spots or from small pieces of tissue. For such samples it is therefore necessary to gain as much information as possible using the native structures. One possibility for analysis is to separate the carbohydrate mixture into neutral and acidic pools using an anion exchange cartridge. This serves to simplify the mixtures entering the mass spectrometer, prevent overlapping peaks, and eliminate suppression problems. Neutral carbohydrates would be analyzed as sodium (or other metal) cations in the positive mode using either ESI or MALDI, and acidic oligosaccharides would be analyzed in the negative mode using either ionization technique. A second possibility is to methyl esterify the unseparated glycans to allow simultaneous ionization of all sugars with semiquantitative responses (Karlsson, Karlsson, & Hansson, 1995; Powell & Harvey, 1996). Analysis of all released carbohydrates would be accomplished at once with minimal sample handling, an advantage for SDS–PAGE gel spots. Tandem mass spectrometric analysis of neutral carbohydrates is best carried out from metal cationized ions in the positive mode or from deprotonated ions. It is not likely that CID spectra will be directly interpretable, 185 & ZAIA and thus investigators are advised to build a library of known structures from their biological system, and to use the information and experience gained to interpret data generated from unknowns. III. ANALYZERS FOR MASS SPECTROMETRY OF CARBOHYDRATES A. Analysis of Permethylated Carbohydrates Using High Temperature GC/MS Capillary gas chromatography (GC) is a fast and sensitive separation technique that is easily interfaced with MS. Molecules eluting from the GC column are subjected to electron impact (EI) ionization and mass spectra acquired. These spectra are characterized by abundant fragment ions and a low abundance of an ion corresponding to the intact molecular ion. Although GC has been widely used to separate monosaccharide residues after degradation (Lindberg & Lonngren, 1978; Geyer & Geyer, 1994), it has also been used in the analysis of derivatized intact oligosaccharides (Hallgren & Lundblad, 1977; Fournet et al., 1980; Nilsson & Zopf, 1982; Nilsson & Zopf, 1983) of up to 15 monosaccharide units in size (Wang, Matsuura, & Sweeley, 1983). The combination of GC with MS provides the significant benefit that mixture components are separated prior to being analyzed by EI MS. Because EI spectra result in complex fragment-ion patterns, a high-resolution separation technique, such as GC, is essential because, under favorable conditions, only one mixture component enters the MS source at a given time. Permethylation of oligosaccharides is easily performed, using the solid sodium hydroxide method (Ciucanu & Kerek, 1984), and that derivative is the most widely studied by high-temperature GC/MS. As with all GC/MS methods, the analysis is limited by the volatility of the analyte molecules. Using thin-film capillary GC columns combined with high temperatures, the analysis of permethylated oligosaccharides up to 2500 Da has been demonstrated (Karlsson, Carlstedt, & Hansson, 1989). The EI fragments of permethylated oligosaccharides correspond to Bm ions, using standard nomenclature (Domon & Costello, 1988b). Although the precursor ion is often absent, the pattern of fragment ions is often useful for differentiation of structural isomers. This result is in sharp contrast to ESI and MALDI, where the intact protonated molecule ion is usually abundant and fragmentation of permethylated oligosaccharides during the ionization process occurs to a much lower degree. It is, therefore, not necessary to separate mixtures prior to analysis with these techniques, and fragment ions may be produced by mass isolation followed by collision-induced dissociation (CID). 186 B. Analysis of Carbohydrates with MALDI-TOF MS Analysis of ions that result from metastable decay in the post-source field-free region of a reflectron time-of-flight mass spectrometer has been used to obtain structural information on biopolymers (Tang et al., 1988; Spengler, Kirsch, & Kaufmann, 1991). That technique entails the sequential lowering of the ion-reflector potential to focus the metastable fragment ions. Now known as PSD, the phenomenon has been observed with MALDI ionization for sialylated glycopeptides (Huberty et al., 1993) as well as oligosaccharides (Harvey et al., 1995; Yamagaki & Nakanishi, 2000). Because MALDI ion signals produced from oligosaccharides are enhanced by reductive amination (Takao et al., 1996) or permethylation (Perreault et al., 1997), the effects of those modifications on the information of MALDI-PSD spectra have been investigated. Reductively aminated oligosaccharides modified with benzylamine or 2-aminopyridine produce a full series of fragment ions that contain the original reducing terminus (Lemoine, Chirat, & Domon, 1996; Okamoto et al., 1997). An optimized calibration procedure has been presented and used for structural characterization of permethylated oligosaccharides by PSD (Viseux, Costello, & Domon, 1999). Cleavage of a(2–3)-linked sialic acids in sialyllactoses has been observed with PSD to occur more readily than cleavage of the a(2–6)-linkage (Yamagaki & Nakanishi, 1999), allowing the isomers to be distinguished based on B1 ion abundance. PSD has been combined with sequential exoglycosidase treatment for oligosaccharide microsequencing (Sato et al., 2000). Product-ion patterns indicative of (1–3) versus (1–4) linkages at the nonreducing termini of 2-aminobenzamide-derivatized tetrasaccharides were observed in this work. The observation of PSD fragment ions has also been used to differentiate Lewis-type linkage isomers in trisaccharides (Yamagaki & Nakanishi, 2000). Localizing a fixed positive charge at the reducing end of carbohydrates has been accomplished by reductive amination with benzylamine followed by N,Ndimethylation (Broberg, Broberg, & Duus, 2000). PSD profiles have been shown to differ for trisaccharide linkage isomers that derive from Lewis-type oligosaccharides (Yamagaki & Nakanishi, 2000). The resulting MALDI PSD spectra were considerably simpler than observed in typical PSD spectra, and showed a series of reducingterminal fragment ions. PSD experiments have, therefore, been demonstrated to be useful in the analysis of oligosaccharides. As with peptides, however, PSD is not the method of choice for fragmentation of carbohydrates. The necessity of stitching together spectra derived from several reflector voltages makes calibration difficult. The number of laser shots required to obtain a coverage of the entire m/z region MASS SPECTROMETRY OF OLIGOSACCHARIDES at sufficient signal-to-noise ratio is also quite high. Further, the product-ion resolution is poor relative to that generated by CID. The recent availability of MALDI sources for analyzers such as the FT, Q-oTOF, and QIT enable far more efficient production and detection of product ions than is possible with PSD. C. Analysis of Carbohydrates with MALDI Q-oTOF MS The development of MALDI ionization sources for quadrupole-orthogonal time-of-flight (QoTOF) type instruments (Morris et al., 1996; Krutchinsky et al., 1998; Loboda et al., 2000) has improved precursor-ion selection and substantially higher product-ion resolution than with MALDI-PSD TOF (Harvey et al., 2000). Using MALDI QoTOF MS, it is possible to obtain a CID MS/MS production spectrum of a sulfated N-linked sugar, and to observe Bm and Yn ions for the entire structure (Wheeler & Harvey, 2001). Spectra of sialylated glycans are reported to be simpler for MALDI QoTOF MS than MALDI-TOF MS due to the absence of metastable fragmentation (Harvey et al., 2000). The analysis of singly charged ions is seen as an advantage relative to that of multiply charged ions generated by ESI (Harvey, 2000a) in that the spectra are easier to interpret. Neutral and acidic glycans released from glycoproteins separated by gel electrophoresis have been derivatized and subjected to reductive amination with 3-acetylamino-6-acetylaminoacridine. Fragmentation observed with MALDI QoTOF MS/MS in the positive mode showed that fragment ions that contain the derivatized reducing terminus were predominant, facilitating the interpretation of the spectra (Hanrahan et al., 2001). D. Analysis of Carbohydrates with ESI Q-oTOF MS The development of ESI QoTOF MS (Morris et al., 1997; Shevchenko et al., 1997) has substantially increased the sensitivity and resolution at which carbohydrate tandem mass spectra can be obtained. Alkali-adducted ions can be analyzed with ESI QoTOF MS (Harvey, 2000a). The use of divalent cations, including Mg2þ, Ca2þ, Mn2þ, Co2þ, and Cu2þ in ESI QoTOF MS/MS, has been studied and found to induce more abundant ions than monovalent metal ions (Harvey, 2001). Most observed fragment ions corresponded to glycosidic bond cleavage rather than cross-ring cleavage. A new derivative for reductive amination, N-(2diethylamino)ethyl-4-aminobenzamide (Harvey, 2000c), has been studied and produces more abundant ions for carbohydrates than do other aromatic-amine derivatives (Harvey, 2000b). Analysis of sialylated sugars with negative-ion QoTOF MS/MS has shown that fragmentation is dramatically influenced by Neu5Ac linkage position & (Wheeler & Harvey, 2000). In that work, it was observed that native deprotonated oligosaccharide ions fragment to produce an ion at m/z 306, corresponding to a 0,4Am, from cleavage across the ring to which the sialic acid is a2-6 linked. Because that ion is absent in the CID profiles of a23 linked isomers, the presence of that ion is diagnostic for the a2-6 linkage. E. Analysis of Carbohydrates with QIT MS Over the last decade, tandem-in-time instruments, in which the multiple stages of mass spectrometry are performed in the same space, as in the FT MS (Marshall, Wang, & Ricca, 1985) and QIT MS (Johnson & Yost, 1985; March & Hughes, 1989; March & Londry, 1995; March, 2000) have become more common. Those tandem-in-time instruments have inherently greater MS/MS efficiency due to their configuration, as compared to tandem-in-space instruments, leading to greater sensitivity and selectivity. The QIT MS has emerged as a remarkably sensitive and selective instrument that is small, low in cost, and capable of efficient MS/MS (Johnson et al., 1990; Todd, 1991; March, 1998; March, 2000). On commercially available QIT MS instruments, it is possible to accurately isolate and fragment a precursor ion, and to selectively fragment only that m/z value, thus minimizing the extent to internal fragment ions are formed. Because of the small time necessary to acquire an MSn mass spectrum, it is possible to couple the QIT MS to liquid chromatography to gain more specific information about a sample. Newer capabilities of commercially available QIT MS instruments include the ability to develop methods for intelligent, data-dependent scanning. Using ESI QIT MS, it is possible to remove non-reducing terminal substituents that suppress cross-ring and core-carbohydrate cleavage in the tandem mass spectra of permethylated complex N-linked oligosaccharides (Reinhold & Sheeley, 1998; Sheeley & Reinhold, 1998; Weiskopf, Vouros, & Harvey, 1998). The data produced from MS3 and MS4 stages allow sequence and linkages to be obtained. Sequential stages of MS have been used as virtual degradative steps to reduce the structures of complex sialylated and fucosylated oligosaccharides to subunits, the spectra of which can be matched against known standards (Viseux, de Hoffmann, & Domon, 1998). Mass spectrometric degradation of oligosaccharides has been combined with a library of known structures to characterize substructural motifs expressed by families of N-linked glycoproteins (Tseng, Hedrick, & Lebrilla, 1999). Different glycan structures contain common substructural motifs that result in common features in tandem mass spectrometric profiles. The tandem mass spectra of known substructural motifs constitute a catalog library, against which the data generated from unknowns are searched. 187 & ZAIA F. Analysis of Glycoconjugates with FT MS Detailed characterization of carbohydrate and glycolipid structures often requires multi-stage decomposition (MSn), making the application of trapped ion type instruments such as the QIT and FTMS advantageous. The FTMS has the considerable advantage over the QIT that ions are detected with high resolution and mass accuracy. The widespread use of FTMS in the analysis of biomolecules has been limited, however, by the lack of well-developed software to enable the facile isolation and fragmentation of ions for MSn experiments. It is anticipated that instruments will be introduced in the near future that will correct this problem, and thereby allow broader use of FTMS. FTMS is compatible with ESI and MALDI, and may be used with several different dissociation techniques, including sustained off-resonance irradiation (SORI) CID (Gauthier, Trautman, & Jacobson, 1991), infrared multiphoton dissociation (IRMPD) (Little et al., 1994), and electroncapture dissociation (ECD) (Zubarev, Kelleher, & McLafferty, 1998). SORI-CID is a general method for the collisional activation of ions trapped in an FTMS cell, and is frequently used for structural analysis of proteins and polypeptides (Guan, Marshall, & Wahl, 1994; Hofstadler et al., 1994; Huang et al., 1994; Senko, Speir, & McLafferty, 1994; Wu et al., 1995; Little et al., 1996; Solouki et al., 1996; Heck & Derrick, 1997; Kelleher et al., 1998; Kelleher et al., 1999; Maier et al., 2000), carbohydrates (Cancilla, Penn, & Lebrilla, 1998; Solouki et al., 1998; Cancilla et al., 1999; Gaucher et al., 2000; Penn, Cancilla, & Lebrilla, 2000), and oligonucleotides (Hettich & Stemmler, 1996; Flora, Hannis, & Muddiman, 2001). With SORI-CID, the use of many low-energy collisions has the advantage of being a slow-heating method so that ions fragment mainly through the lowest energy fragmentation channel. IRMPD, another low-energy fragmentation method, has previously been shown to be an efficient and selective fragmentation method for sequencing of proteins (Little et al., 1994; Dufresne, Wood, & Hendrickson, 1998) and DNA (Little & McLafferty, 1995; Little et al., 1996). IRMPD has been used to analyze N-linked glycopeptides with the result that extensive glycan fragmentation is observed with no ion from cleavage of the peptide backbone (Håkansson et al., 2001). IRMPD has also been used to analyze sulfated oligosaccharides derived from chondroitin sulfate and to result in more extensive losses of SO3 than observed with SORI-CID (McClellan et al., 2002). ECD has emerged as a new odd-electron fragmentation method for FT MS (Zubarev, Kelleher, & McLafferty, 1998), based on partial recombination of multiply protonated polypeptide molecules with thermal electrons (Kruger et al., 1999). Peptide backbone cleavage occurs at 188 the Ca –N bond rather than at the amide linkage as occurs with classical CID or IRMPD. The exothermic electron capture reaction induces the formation of an atomic hydrogen H., the energetic transfer of which to the backbone carbonyl permits the formation of primarily the zn.þ and cnþ ions (Stensballe et al., 2000). Peptide fragmentation with ECD typically cleaves all Ca –N bonds along the peptide backbone, often yielding a complete sequence, depending on the signal-to-noise ratio of the resulting fragment ions. The notable exception to that rule is that proline, being cyclic around the Ca –N bond, does not generate fragment ions. ECD is also the preferred method for the analysis of modified peptides because backbone cleavages are usually observed (Kelleher et al., 1999; Mirgorodskaya, Roepstorff, & Zubarev, 1999; Stensballe et al., 2000; Shi et al., 2001). The use of ECD in the analysis of carbohydrates is starting to be appreciated. It is well-accepted that sialic acid (Neu5Ac) and fucose (Fuc) residues are readily lost during low-energy CID, a fact which can make identification of the branching location difficult. It is not known at this time the extent to which ECD will allow carbohydrate branching patterns to be determined. ECD has proven to be useful in the analysis of glycopeptides, as will be discussed in Section IV.C. G. Conclusions In summary, any of the modern mass analyzers may be used in the analysis of oligosaccharides. For profiling oligosaccharide mixtures, MALDI has been observed to provide more even ionization response as mass increases than does ESI. ESI, on the other hand, is less likely to cause metastable fragmentation than in MALDI—an important concern for the analysis of fragile sulfated and/or sialylated oligosaccharides. The use of high-pressure MALDI sources to minimize the extent to which metastable fragmentation of oligosaccharide samples occurs is recommended. Conventional low-pressure MALDI-TOF is recommended for profiling oligosaccharide mixtures, provided that measures are taken to address the fragility and ionization potentials of acidic oligosaccharides (see Section V). Although MALDI-TOF is not recommended for sulfated oligosaccharides, particularly those with more than one sulfate group, due to extensive fragmentation in the source (see Section VI.B.2.b), high-pressure MALDI has been observed to be significantly gentler and is applicable to sialylated glycoconjugates. The use of MALDI-QoTOF for tandem MS of oligosaccharides is attractive, particularly for those sample that have been methyl-esterified to stabilize sialic acid linkages. It is clear that this geometry will be sensitive enough to allow analysis of oligosaccharides released from SDS–PAGE gel spots. MASS SPECTROMETRY OF OLIGOSACCHARIDES Although the responses appear to diminish as oligosaccharide molecular weight increases, ESI is more appropriate for use with fragile compound classes. Analysis of sialylated (see Section V.D.) and sulfated (see Section VI.C.) oligosaccharides with negative-mode ESI allows extremely useful CID product-ion patterns to be produced. The QIT analyzer has been shown to be particularly useful for oligosaccharides because of its efficiency in acquiring multiple stages of MS fragmentation. This efficiency allows fragment ions to be produced that are absent in MS2 profiles, increasing the amount of structural information produced. The FTMS carries similar advantages as the QIT but with higher resolution and mass accuracy. That resolution will be particularly useful for the analysis of multiply charged ions such as those that are produced from polysulfated carbohydrates with ESI (see Section VI.C). At the present time, however, the application of FTMS to this and other biomolecular structural questions is limited by the lack of software development in commercial instruments. The development of hybrid QIT-TOF and QIT-FT mass spectrometers will benefit the analysis of oligosaccharides by increasing the resolution and mass accuracy of fragment ions. That increase is particularly important for polysulfated oligosaccharides because the precursor ions are multiply charged. Although QIT-TOF instruments are available with MALDI, problems with metastable fragmentation, similar to those observed with TOF/TOF instruments (Medzihradszky et al., 2000; Yergey et al., 2002), are likely to limit their application to oligosaccharides. The development of high-pressure MALDI and ESI sources is, therefore, important. The MALDI TOF/TOF is the only modern commercial instrument that is capable of high-energy CID, and may prove to be very useful for the production of cross-ring cleavage ions for oligosaccharides—provided that the ions can be produced without extensive metastable fragmentation before the collision cell. Because data published at the present time indicate that there is a problem with metastable fragmentation, resulting in low-energy product ions (Medzihradszky et al., 2000; Yergey et al., 2002), the development of a high-pressure source for that instrument is needed. IV. TANDEM MASS SPECTROMETRY OF GLYCOPEPTIDES A. Ionization of Glycopeptides Glycopeptides ionize poorly with FAB ionization due to their high mass, hydrophilicity, and limited surface activities in liquid matrices (Carr et al., 1990). Synthesis of tBoc derivatives of glycopeptides was observed to increase & the FAB sensitivity over underivatized glycopeptides by a substantial degree (derivatized molecular mass 1522.8) (Medzihradszky et al., 1990). The identification of Nlinked glycosylation sites with FAB relied on the use of PNGase F, an enzyme that releases N-linked glycans and converts the Asn side chains to which they are bound to Asp. Knowing the protein sequence, the one Da shift in mass was used to determine the site of modification from the FAB MS/MS spectra (Carr et al., 1989). It was possible to HPLC-fractionate the components of a proteolytic digest of a glycoprotein before and after glycosidase treatment and to conduct FAB MS analysis of the isolated fractions (Carr & Roberts, 1986). Endoglycosidase cleavage of N-linked glycans in the presence of H218O introduces an isotopic label into the Asp residue created by the enzymatic hydrolysis reaction, the mass signature of which is useful for identification purposes (Küster & Mann, 1999). The development of ESI (Meng, Mann, & Fenn, 1988) resulted in significantly better glycopeptide signal response than was observed with FAB. This improvement allowed the development of in-source fragmentation and precursor-ion scans for the detection of oxonium ions generated from glycopeptides eluting from an HPLC column (Conboy & Henion, 1992; Huddleston, Bean, & Carr, 1993). Analysis of glycopeptides with ESI has been widely applied. The in vitro glycosylation of mucin peptides has been followed with ESI MS to determine the mass and composition of the product glycopeptides (Tetaert et al., 1994). Detection of oxonium ions has been used to provide a detailed insight into site-specific distribution of N-linked groups on immunoglobulin A (Kragten et al., 1995) and for the selective identification of glycopeptides in complex glycoprotein proteolytic digests prior to analysis with MALDI-TOF coupled with exoglycosidase digestion (Yang et al., 1997; Colangelo et al., 1999). An oxonium ion has been used to selectively identify a sialyl Lewisx antigen on a1-acid glycoprotein (Dage, Ackermann, & Halsall, 1998) (for more details, see Section IV.B.1). The usefulness of MALDI-TOF MS for analysis of glycoproteins was recognized soon after its invention (Overberg et al., 1990). The MALDI-TOF mass spectrum of a1-acid glycoprotein exhibited a broader peak than observed for a typical unmodified protein in the same mass range, and it was suggested that this broadness reflected the glycoform heterogeneity (Hillenkamp et al., 1991). The contribution of prompt and metastable fragmentation has been evaluated for human interferon-g and found not to contribute significantly to the MALDI-TOF MS peak width (Mortz et al., 1996). Thus, MALDI-TOF MS peak widths appear to depend on the heterogeneity of glycosylation. Differences in glycosylation between fetal and adult cartilage oligomeric matrix protein have been identified with MALDI-TOF MS analysis of the intact 80 kDa glycopro189 & ZAIA teins as well as proteolytic glycopeptides (Zaia et al., 1997). The acquisition of MALDI-TOF mass spectra in conjunction with glycosidase treatment is quite a useful technique to determine the total mass of carbohydrate on a glycoprotein (Zaia et al., 1997; Colangelo et al., 1999; Jacoby et al., 2000; Tarelli et al., 2000; Zaia et al., 2001). Enzymatic deglycosylation of proteoglycans can also be accomplished in conjunction with MALDI-TOF MS (Zaia et al., 2000). B. CID of Glycopeptides 1. Selective Identification of Glycopeptides with Tandem MS Tandem MS of glycopeptides results in the formation of ions that are useful for recognition purposes (Conboy & Henion, 1992; Carr, Huddleston, & Bean, 1993; Huddleston, Bean, & Carr, 1993). As shown in Figure 23, a complex biantennary N-linked glycopeptide fragments to produce ions that result from neutral losses from the precursor, corresponding to HexmHexNAcn. It is, therefore, possible in theory to identify glycopeptides in complex tryptic digest mixtures with constant neutral-loss scans. Also observed in glycopeptide CID mass spectra are oxonium ions that correspond to Hexþ (C6H11O5þ, m/z 163.06), HexNAcþ (C8H14NO5þ, m/z: 204.09), and HexHexNAc (C14H24NO10þ, m/z: 366.14). Note that the abundance of the Hexþ ion is often quite low. Thus, a scan for the precursors of those oxonium ions would appear to be useful to identify glycopeptides. Neutral-loss scans are carried out most often on triple quadrupole analyzers by scanning precursor ions while monitoring only those product ions that result from a constant-neutral loss. Precursor ions scans are accomplished by scanning the precursor ions while detecting only those product ions at a given m/z value. Because it is possible for a fraction of unmodified peptides to produce CID product ions at the same nominal mass as the carbohydrate oxonium ions, the higher the resolution of the selecting quadrupole, the more selective the measurement. The performances of triple-quadrupole and Q-oTOF analyzers have been compared for precursor-ion scans for phosphopeptides identification (Steen, Küster, & Mann, 2001). The results, showing that greater selectivity was achievable with the Q-oTOF analyzer, are likely to be applicable to the detection of oxonium ions for glycopeptides. The precursor-ion scanning approach has been applied to the selective identification of glycopeptides with reversed phase LC/MS (Carr, Huddleston, & Bean, 1993). That work demonstrated the use of precursor ions scans for m/z 204 for the detection of fetuin glycopeptides. The approach entails comparing the LC/MS total-ion chromatogram with that corresponding to the precursors of m/z 204, and clearly identifies the elution times and m/z values of glycopeptides in the tryptic-digest mixture. Another example of that approach is shown in Figure 24 for the analysis of a recombinant human thrombomodulin tryptic FIGURE 24. Peptide/glycopeptide map of recombinant human thromFIGURE 23. Product-ion MS/MS of m/z 878 [M þ 2H]2þ. The asterisk (*) indicates ions formed from two bond cleavages. Reproduced with permission from Huddleston, Bean, & Carr (1993). Copyright 1993 American Chemical Society. 190 bomodulin tryptic digest. (A) UV chromatogram at 206 nm, (B) total-ion chromatogram produced by positive-ion LC/MS, (C) precursor-ion scan for m/z 204 (HexNAcþ). Reproduced from Itoh et al. (2002) with permission. Copyright 2002, Elsevier. MASS SPECTROMETRY OF OLIGOSACCHARIDES digest mixture (Itoh et al., 2002). The data show that the glycopeptides are clearly identified, based on the m/z 204 precursor ion scan shown in (C). Precursor-ion scans have also been used to identify glycosylated peptides in unseparated tryptic digest mixtures ionized directly with nano ESI MS (Wilm, Neubauer, & Mann, 1996). Glycopeptides present in proteolytic digests of glycoproteins have also been selectively eluted from reversed phase HPLC columns under neutral pH conditions, facilitating their analysis with LC/MS (Ohta et al., 2001). That approach is limited by the complexity of the mixture that may give rise to suppression of glycopeptide ions. 2. CID of O-Linked Glycopeptides High-energy tandem mass spectrometric analysis of a monomannosylated glycopeptide showed peptide-backbone product ions that contained the carbohydrate group (Medzihradszky et al., 1990). In the same work, the tandem mass spectrum of tBoc-YGP-(GalGalNAc)T-P-(GalGalNAc)S-A was dominated by the fragmentation of the carbohydrate side chains rather than of the peptide backbone. An ion that correspond to the unmodified peptide was, however, observed. Analysis of that ion with a subsequent stage of MS would serve to identify the peptide portion of the molecule. High-energy tandem mass spectrometric analysis of an O-(HexHexNAc)-modified glycopeptide indicated an abundant series of peptide-backbone product ions with a single yn ion that contained the disaccharide and indicated its site of modification (Medzihradszky et al., 1996). Abundant and useful peptide-backbone product ions were also observed for a peptide modified with two (HexHexNAc) groups. High-energy tandem mass spectra of O-linked glycopeptides contained relatively more abundant ions produced by glycosidic fragmentation and lessabundant ions produced by peptide-backbone fragmentation as the size and number of carbohydrate modifications increased (Medzihradszky et al., 1996). Low-energy tandem mass spectrometric analysis of a peptide modified with a single glycan of composition (NeuGc)2GalGalNAc resulted in abundant product ions that corresponded to sequential losses of NeuGc, NeuGc, Hex, and HexNAc, respectively, from the precursor ion (Hirayama et al., 1998). The ion that corresponded to the unmodified peptide was abundant, and would be amenable to analysis with a subsequent MS stage if desired. Lowenergy tandem mass spectrometric analysis of synthetic O-linked glycopeptides on a QoTOF instrument has shown that product ions are generated in very low abundance from a cleavage of the peptide backbone, a few of which contain the carbohydrate group (Alving, Paulsen, & PeterKatalinic, 1999). Extended data-accumulation times are required to achieve a sufficient signal-to-noise ratio for these ions. The data sufficed to identify the sites of & glycosylation in synthetic glycopeptides that contained one or two glycans of up to tetrasaccharide size. Abundant ions were observed that corresponded to the unmodified peptide that could be subjected to an additional MS stage for the purpose of identification in the case of an unknown. Monoand difucosylated O-linked glycopeptides produce abundant fragments that corresponded to loss of dHex from the precursor ion on an ESI QoTOF instrument (Macek, Hofsteenge, & Peter-Katalinic, 2001). Peptide-bond product ions that contained the dHex group, however, were observed at very low abundances in the spectra. It appears that, at low collision energy, the ion that corresponds to loss of dHex from the precursor ion does not undergo subsequent fragmentation. The results demonstrated that tandem mass spectra with sufficiently high signal-to-noise ratio can be used to assign sites of O-linked fucosylation at low collision energies. Given the difficulties with assigning O-linked glycosylation sites, chemical elimination of the glycans has been used in conjunction with tandem MS. A procedure based on b-elimination with hydroxylamine incorporates ammonia into the Ser or Thr residue to which the glycans had been attached (Rademaker et al., 1998). An alternative approach that used alkylamines to release the O-linked glycans produced a larger mass difference between the modified and unmodified Ser/Thr residues, facilitating the analysis (Hanisch, Jovanovic, & Peter-Katalinic, 2001). 3. CID of N-Linked Glycopeptides ESI triple-quadrupole tandem mass spectra of highmannose type N-linked glycopeptides result in abundant ions that correspond to fragmentation of the glycan (Bateman et al., 1998; Hirayama et al., 1998; Zeng et al., 1999; Zhu et al., 2000; Hui et al., 2001). An example of a CID profile obtained from a Man9 N-linked glycopeptide is shown in Figure 25 (Nemeth et al., 2001). Because of the lack of HexNAc residues in the antennae, the glycan portion of the molecule fragments at a collision energy high enough to allow peptide-backbone fragmentation to be observed, as shown in the top panel. The ions labeled in the spectrum are identified in Table 5. The ion that corresponds to HexNAc-modified peptides can be generated in-source by using elevated declustering potential and analyzed using MS/MS. A more useful approach, given the complexity of ESI mass spectra generated from glycoprotein proteolytic digests, would be to use MS3 on a QIT or FTMS instrument. The generation of such data would identify the site of modification for the high-mannose structure, and augment the information on the glycan produced in the MS/MS spectrum. A glycopeptide modified with two high-mannose N-linked glycans produces an abundant series of ions that correspond to respective losses of Man and GlcNAc, and displays an 191 & ZAIA FIGURE 25. Nano ESI MS/MS analysis of a triply protonated Man9 N-linked glycopeptide (a) the mass spectrum where peptide yn ions are indicated, and (b) the singly charged deconvoluted mass spectrum of panel a where the sugar-loss ion series is indicated. Reproduced from Nemeth et al. (2001) with permission. Copyright 2001 American Chemical Society. abundant ion that corresponds to the peptide with two GlcNAc residues (Zhu et al., 2000). As shown in Figure 26, complex N-linked glycopeptides undergo fragmentation of labile HexNAc and Neu5Ac residues. The result is that an abundant ion that corresponds to the peptide þ GlcNAc is observed, but that the peptidebond cleavage ions are lower in abundance than observed in CID mass spectra of high-mannose structures. This lack TABLE 5. List of the observed product ions from the nano ESI MS/MS analysis of a Man9 glycopeptide [C12 þ 3H þ Man9GlcNAc2]3þ [reproduced from Nemeth et al. (2001) with permission, Copyright 2001 American Chemical Society] a M, Mannose. G, N-acetylglucosamine. b 192 MASS SPECTROMETRY OF OLIGOSACCHARIDES & FIGURE 26. Product-ion scan of a trisialylated tri-antennary N-linked glycopeptide [M þ 4H]4þ ion. Reprinted with permission from Ritchie et al. (2002). Copyright 2002, Elsevier. of peptide fragmentation behavior makes it difficult to analyze the oligosaccharide and peptide structures in an unknown glycopeptide in a single measurement. The observation of an ion that corresponds to peptide þ GlcNAc allows a variation on the oxonium-ion precursor-ion scan for selective detection of glycopeptides. By scanning an unseparated glycoprotein tryptic digest for precursors to the peptide þ GlcNAc ion (corresponding to the oligosaccharide Y1 ion), it is possible to identify all glycoforms that modify a given peptide (Ritchie et al., 2002). C. Electron Capture Dissociation of Glycopeptides ECD is a powerful tool for peptide sequencing, and allows the determination of the location of post-translational modifications like O-glycosylation (Mirgorodskaya, Roepstorff, & Zubarev, 1999), N-glycosylation (Håkansson et al., 2001), gamma carboxylation (Kelleher et al., 1999), and phosphorylation (Stensballe et al., 2000; Shi et al., 2001). It is believed that because of the high H. affinity of the electron-rich amide carbonyls, minimal fragmentation occurs to post-translational modification groups. It has been noticed, however, that disulfide bonds tend to fragment preferentially with ECD (Zubarev et al., 1999). The finding that ECD produces peptide-backbone fragmentation with minimal fragmentation of glycans is particularly significant because it allows the determination of sites of glycosylation and the size of the glycan. Given that ECD cleaves the peptide backbone without fragmenting the glycan, it is of interest to use it in combination with IRMPD or SORI-CID. IRMPD of glycopeptides results in an extensive fragmentation of the glycan with little or no peptide-bond fragmentation, and thus produces data complementary to that of ECD, as shown in Figure 27 (Håkansson et al., 2001). Cleavages of 11 of the 15 backbone amine position are observed in the ECD profile (a), providing data that would be useful for peptide database searching. The fragmentation observed in the IRMPD mass spectrum of the same glycopeptide (b), resulted exclusively from the glycan, and was sufficient to determine that there are three branching sites in the structure. SORI-CID of glycopeptides also results in preferential fragmentation of the glycan, and has also been used to complement ECD (Kjeldsen et al., 2003). Although the combination of ECD with IRMPD or SORI-CID presently requires a specially modified FTMS instrument and a high level of operator skill, a combined fragmentation approach is likely to be extremely useful for the characterization of protein glycosylation once the technology becomes commercially available. D. Conclusions Mass spectrometric analysis of glycopeptides entails careful consideration of the ionization and the fragmentation characteristics of the glycan portion of those molecules. The goal of the analysis is to obtain as much information on the glycan and peptide structures as possible for a limited quantity of material, such as might be produced from SDS– PAGE gel spots. It is essential not to produce prompt or metastable fragmentation from the ionization process, and thus the use of ESI is indicated at the present time. Although the use of high-pressure MALDI is likely also to produce protonated molecule ions from glycopeptides without metastable decay, it is essential that the sources be tested with this compound class to determine that the conditions are cool enough. Use of the negative mode will maximize the stability of native sialylated glycopeptides 193 & ZAIA FIGURE 27. Comparison of ECD and IRMPD product ion profiles for a glycopeptide. (a) ECD FTICR mass spectrum obtained from the triply protonated N-glycosylated peptide of m/z 1005.5; (b) IRMPD FTICR mass spectrum (displayed in three segments) from the triply protonated N-glycosylated peptide of m/z 1005.5; (c) Peptide sequence and glycan structure of the investigated N-glycosylated peptide. Top: Dissociation sites for ECD. Bottom: Major dissociation sites for IRMPD. Reproduced from Håkansson et al. (2001) with permission. Copyright 2001 American Chemical Society. during the MALDI process. Methyl esterification of sialic acid residues is recommended if the positive MALDI mode will be used. Use of traditional low-pressure MALDI sources in the negative linear mode will be useful for mass profiling. CID fragmentation of glycopeptides results in the production of abundant ions from glycosidic bond and very low abundance ions from peptide-bond fragmentation. It is 194 generally not possible to identify the modified amino acid residues, although the number of possibilities in a given peptide will be limited. Even the determination of the peptide mass from the CID profile is not a straightforward matter. Useful data are obtained from an extended summing of ESI or MALDI CID data to obtain adequate signalto-noise ratios for the low-abundance peptide-backbone cleavage ions. The use of ECD, however, results in MASS SPECTROMETRY OF OLIGOSACCHARIDES abundant peptide-backbone cleavage ions with no fragmentation of the glycan. It is, therefore, possible to assign the modified amino acid residues even for cases in which the peptide is modified with several glycan structures. Clearly, the most useful approach for the analysis of glycopeptide is to acquire ECD and CID mass spectra on an FTMS instrument. Alternatively, a combination of ECD and IRMPD can be used. V. MASS SPECTROMETRY OF SIALYLATED GLYCOCONJUGATES A. Permethylation of Sialylated Oligosaccharides Permethylation has been used for 40 years as a technique to improve the analytical properties of glycoconjugates (Hakomori, 1964). The primary advantages of this derivatization step for mass spectrometry are that (1) the ionization responses for FAB, ESI, and MALDI are improved relative to those of the native molecules, and (2) that the product-ion mass spectra are much more informative relative to those of the underivatized sugars. These advantages must be balanced against the disadvantages of the derivatization step itself. The derivatization chemistry places a lower limit on the quantity of sample that can be analyzed, and as a result does not appear to be appropriate for analysis of samples available only in small quantities such as from gel spots. Use of the solid sodium hydroxide technique (Ciucanu & Kerek, 1984) has been found to be effective to permethylate sialylated glycoconjugates, and has come to be widely used (Dell et al., 1994; Perrault & Costello, 1994; Reinhold, Chan, & Reinhold, 1994; Reinhold, Reinhold, & Costello, 1995; Reinhold & Sheeley, 1998; Viseux, de Hoffmann, & Domon, 1998). The technique involves vortexing the dried sample in anhydrous dimethyl sulfoxide in the presence of powdered sodium hydroxide, followed by an incubation at room temperature in the presence of methyl iodide. The reaction is quenched by the addition of water, and the permethylated molecules are extracted into chloroform. Note that this procedure is not recommended for sulfated oligosaccharides because they are not soluble in chloroform. The short Hakomori method is used for those cases (Dell et al., 1994). In addition, the solid sodium hydroxide method is not recommended for base-labile molecules. Permethylated sialic residues are hydrolyzed under basic conditions, and a modification to the permethylation procedure is recommended (Barr et al., 1991). That modification entails back-extracting the permethylated mixture with 30% acetic acid to minimize the chance of baseinduced hydrolysis (Reinhold & Sheeley, 1998; Viseux, de Hoffmann, & Domon, 1998; Viseux, Costello, & Domon, & 1999; Delaney & Vouros, 2001; Viseux et al., 2001). Baseinduced hydrolysis of permethylated sialylated oligosaccharides has been used to simplify the interpretation of the product-ion mass spectra of those molecules (Viseux, de Hoffmann, & Domon, 1998; Viseux, Costello, & Domon, 1999). The hydrolyzed permethylated sialic acid groups leave an open hydroxyl group that may be perdeuteromethylated. The position of sialic acid substitution can be determined from the CID spectrum (see Section V.D). B. MALDI-MS of Sialylated Glycoconjugates Early work on carbohydrate identification from proteolytic digests of recombinant glycoproteins demonstrated that the majority of ions observed in the linear mode for sialylated glycoproteins correspond to metastable ions (Huberty et al., 1993). The loss of most Neu5Ac residues is observed in the reflector mode with a-cyano-4-hydroxycinnamic acid or sinapinic acid MALDI matrices. The use of 2,6-dihydroxyacetophenone with ammonium citrate was subsequently observed to diminish that fragmentation (Pitt & Gorman, 1996). A similar matrix, 2,4,6-trihydroxyacetophenone (THAP) with ammonium citrate, was found to enable the detection of sialylated N-linked sugars without decomposition and to facilitate analysis of sialylated glycopeptides (Papac, Wong, & Jones, 1996). Given this metastable decay, acidic glycoconjugates are best analyzed in the negative linear MALDI-TOF mode. With careful use of threshold laser irradiance and a cool matrix such as DHB, it is possible to detect ions that correspond to intact acidic glycoconjugates. The ease with which this detection is possible varies for different instrumental geometries and source designs, and it is advisable to test sample preparation and source conditions in the negative linear mode with sialylated standards. Without the use of such standards, it is difficult to determine whether ions that differ by the mass of a sialic acid residue result from insource fragmentation or are present in solution (Kakehi et al., 2001). Although neutral and acidic N-linked glycans have been analyzed simultaneously in the positive mode on a MALDI Q-oTOF instrument, it is not clear how much in-source fragmentation of sialylated glycans occurs (Hanrahan et al., 2001). A sialylated glycan has been analyzed in the positive MALDI-TOF mode as a mixture of [M þ H]þ, [M(Na) þ H]þ, and [M(K) þ H]þ ions without reported loss of the sialic acid residue (Charlwood et al., 1999). That observation is consistent with significant variability in the prevalence of in-source decay for sialylated glycans among different instruments from different manufacturers. To simultaneously obtain equal ionization responses for neutral and acidic glycoconjugates, it is best to modify either the MALDI matrix or to esterify the glycans (see below). 195 & ZAIA 1. Anionic Dopants for Analysis of Sialylated Glycoconjugates Neutral oligosaccharides are usually analyzed in the positive ion mode using a MALDI-TOF with adducts of alkali cations such as Liþ, Naþ, or Kþ. Neutral and sialylated oligosaccharides bind those cations differently, and thus produce different ionization responses in positive-ion MALDI-TOF MS. The addition of a low concentration of H2SO4 to the matrix solution results in the observation of [M þ H2SO4] ions for neutral oligosaccharides with a harmane matrix (Wong et al., 1999). Sialylated oligosaccharides produce predominantly [M–H] ions under the same conditions, and a mixture of neutral and acidic oligosaccharides results in the observation of ions that correspond to both oligosaccharides in the same spectrum. In an extension of that work, alkylsulfonates have been used as anionic dopants in place of H2SO4 (Wong, Wang, & Lebrilla, 2000). Of the several alkylsulfonates and matrices tested, the combination of sulfanilic acid with dihydroxyacetophenone was found to be most effective for the analysis of mixtures of neutral and acidic oligosaccharides. The ratio of intensity of a mixture that contained a neutral and an acidic oligosaccharides was found to be linear, with a correlation coefficient of 0.99, as the concentration was varied, indicating that suppressive effects are minimized during the ionization process. It was possible to follow the course of sialic acid hydrolysis, using the combination of sulfanilic acid and dihydroxyacetophenone matrix. 2. Methyl Esterification to Stabilize Sialic Acid Residues Conversion of sialic acid residues to methyl esters (Handa & Nakamura, 1984) allows the detection of sialylated oligosaccharides and ganglioside ions in the positive-ion mode without metastable decay (Powell & Harvey, 1996). The sialylated molecules are dissolved in dimethylsulfoxide, and incubated with methyl iodide at room temperature; MALDI analysis can be accomplished without sample cleanup following removal of solvent. Sialic acids can also be separated from neutral oligosaccharides with anion exchange chromatography, and conveniently and efficiently converted to methyl esters while still bound by eluting dimethylsulfoxide and methyl iodide (Karlsson, Karlsson, & Hansson, 1995). Methyl esterification has also been used to facilitate MALDI-TOF analysis of Nlinked glycans released by treatment of SDS–PAGE gel spots with glycosidases (Küster et al., 1997). Once methylesterified, the glycans are passed through a gel-loader pipette tip that is packed with C18, anion, and cation exchange resins to remove salts and detergents prior to MALDI MS. The acidic residues are rendered neutral 196 through esterification, allowing all released glycans to be analyzed in a single MALDI-TOF mass spectrum. The methodology has been applied to profile an a1-acid glycoprotein from different species with MALDI-TOF MS (Küster et al., 1998). Digests of pectin, a homogalacturonan, have been analyzed as methyl esters, using MALDI-TOF with THAP matrix (Körner et al., 1998), and the method has been used to study enzyme specificities of pectin lyases (Körner et al., 1999). Underivatized polygalacturonans derived from alginate have been analyzed with perfluorosulfonated ionomer (Nafion)-coated probes for MALDI-TOF MS (Jacobs & Dahlman, 2001). The immobilized sulfate groups evidently scavenge sodium and potassium ions, resulting in the detection of [M–H] ions with little or no adduction and much cleaner spectral profiles. Taken together, those studies show how MALDI-TOF can be used to provide useful information on acidic biopolymers. 3. Perbenzolylation to Stabilize Sialic Acid Residues Perbenzolylation (Daniel, 1987) has been shown to be effective to stabilize sialic acid groups and to allow their analysis with positive-mode MALDI-TOF MS (Chen et al., 1999). Carbohydrates may be benzoylated in 50 pmol quantities with benzoyl anhydride followed by purification on a C-18 cartridge. The derivatization step is general in nature, and will work on reduced and unreduced oligosaccharides. The spectra of benzoylated glycans that contain 2-3 linked sialic acid residues produce distinctly different MALDI-TOF mass spectral patterns than those that contain 2-6 linked residues. The former undergo internal lactonization of the carboxyl groups during the derivatization process, resulting in the addition of one fewer benzoyl group than the 2-6 linked sialic acid. The technique may be useful to differentiate sialic acid linkages in relatively pure samples. CID of perbenzylated cerebrosides has also been shown to be useful to differentiate oligosaccharide isomers (Perreault & Costello, 1996). 4. High-Pressure MALDI of Sialylated Glycoconjugates The use of FTMS in the analysis of glycoconjugate mixtures carries the significant advantages that the highest possible resolution and mass accuracies are obtained. The use of MALDI with FTMS, as with other analyzers, is advantageous because it has a lower sensitivity to contaminants than does ESI, and it is more easily applied to samples from multiwell formats. In addition, MALDI is more easily interfaced with surface techniques such as thinlayer chromatography and gel electrophoresis. Because the MALDI source is thoroughly decoupled from the FT MASS SPECTROMETRY OF OLIGOSACCHARIDES analyzer, the resolution and mass accuracy do not depend on the smoothness of the desorption surface. The quality of MALDI-TOF spectra, on the other hand, is significantly degraded by surface irregularities. The use of MALDI FTMS in the analysis of fragile biomolecules has been limited until recently by long-lived metastable decay of MALDI ions prior to detection. Such decay is detrimental to the analysis of mixtures becasue it is not possible to determine whether only a given ion is an independent species, or is one that is produced from fragmentation of another ion. MALDI of ions at elevated pressure has been demonstrated to produce diminished metastable decay (Krutchinsky et al., 1998; Laiko, Baldwin, & Burlingame, 2000; Loboda et al., 2000). As Figure 28 shows, sialylated gangliosides are observed as de-sialylated ions with low pressure MALDI FTMS (O’Connor & Costello, 2001). Desorbing the ions using the same conditions but at elevated pressure (1–10 mbar) results in the observation of the intact sialylated ganglioside with ions produced from losses of sialic acid residues in very low abundances. Those results demonstrate that the thermal stabilization of vibrationally excited MALDI ions dramatically reduces the degree of metastable fragmentation observed for fragile sialylated glycoconjugates. Gangliosides with as many as 5 sialic acid residues have been observed by MALDI FTMS with minimal metastable fragmentation (O’Connor, Mirgorodskaya, & Costello, 2002). Those developments will allow the direct profiling of glycolipids from thin-layer chromatography plates, and will facilitate the analysis of any sialylated glycoconjugate. & C. ESI MS of Sialylated Oligosaccharides Given that ionization is dependent on the oligosaccharide composition with respect to neutral and acidic residues, modification of the carbohydrate structure is necessary to provide a profile that reflects the relative quantities of each mixture component. One way to equalize ionization responses is to remove sialic-acid residues enzymatically, thereby rendering all oligosaccharide structures neutral, assuming that sulfated residues are not present. That step will simplify the mass profile due to the elimination of sialic acid heterogeneity (Küster et al., 1997, 1998). To preserve the sialic acid structure while producing a uniform ionization response, it is necessary to neutralize the acidity of the carboxyl groups by methyl ester formation. The methods described in Sections III.A, V.A, and V.B.2 are applicable to the ESI MS of sialylated oligosaccharides. Permethylation also renders sialylated and neutral oligosaccharides chemically equivalent. It is also possible to permethylate sialylated sulfated glycans, although a more involved procedure must be used (see Section VI.A) (Dell et al., 1994). The permethylated oligosaccharides produce stronger ion signals in ESI due to their increased surface activities (Bahr et al., 1997; Cole, 1997; Karas, Bahr, & Dulcks, 2000). The signals are also less sensitive to the composition of the oligosaccharide. The ESI response is such that quantitative analysis can be performed on oligosaccharides released from recombinant glycoproteins (Viseux et al., 2001). Quantitative characterization of glycan mixtures has traditionally involved reducing-end labeling with a chromophore such as 2-aminobenzamide FIGURE 28. GangliosideGT1b desorbed from ATT matrix (a) without collision gas, and (b) with collision gas. Peaks due to the presence of a lower homolog are marked LH. Reprinted with permission from O’Connor & Costello (2001). Copyright 2001 John Wiley & Sons Limited. 197 & ZAIA TABLE 6. Relative intensities of the components identified by mass spectrometric and HPLC analyses of the sCD4 glycan pool [reprinted with permission from Viseux et al. (2001), Copyright 2001 American Chemical Society] a Symbols: B, biantennary N-glycan; F, fucosyl residue; S, sialyl residue; MM, molecular mass. FIGURE 29. HPLC chromatogram of the 2-aminobenzamide-deriva- tized N-glycans (a) and electrospray mass spectrum of the permethylated oligosaccharide derivatives (b) released from human R1-acid glycoprotein. Symbols: X includes biantennary (B), triantennary (T), and tetraantennary (Q) N-glycans; F, fucosyl residue; S, sialyl residue; L, lactosamine unit. Reprinted with permission from Viseux et al. (2001). Copyright 2001 American Chemical Society. followed by HPLC separation. Figure 29 compares the HPLC chromatogram obtained from a1-acid glycoprotein after labeling with 2-aminobenzamide with the ESI mass spectrum obtained from the same sample after permethylation. The glycans are separated into peaks based on the antenna structure and degree of sialylation. The ESI mass spectrum of the permethylated mixture (lower panel) demonstrates that significantly more detailed information is obtained. Specifically, although the fucosylated glycoform is clearly resolved in the mass spectrum, it is not in the chromatogram of the 2-aminobenzamide-labeled material. As Table 6 shows, good agreement in quantitation was obtained between the two methods. The coefficients of variation values obtained for the mass spectrometric method were reported to be less th an 3% for most measurements. In addition to an increased ionization efficiency and desirable fragmentation patterns (Viseux, de Hoffmann, & Domon, 1998), permethylated oligosaccharides may be analyzed by reversed-phase HPLC. Permethylated oligo198 saccharides may be eluted on-line into an ESI mass spectrometer with acetonitrile/water gradients that contain 1 mM sodium acetate to create stable sodium adducts (Delaney & Vouros, 2001). The technique has been shown to be applicable for the analysis of complex N-linked glycans with or without a 2-aminobenzamide label. For very complex mixtures, it is often not possible to detect minor component in the absence of chromatographic separation. Chromatographic separation simplifies the complexity of the mixture that enters the instrument at any moment, addressing this limitation. Thus, with on-line separation, the problem of overlapping peaks in mass spectra is minimized. An alternative is to remove sialic acid residues enzymatically to reduce the complexity to the point that minor components can be detected without chromatographic separation (Viseux et al., 2001). D. Tandem MS of Sialylated Oligosaccharides Unlike their neutral counterparts, native sialylated oligosaccharides produce abundant signals in negative ionization. Using negative FAB MS, sialylated milk oligosaccharides displayed metastable fragmentation that provided useful structural information (Egge & PeterKatalinc, 1987). A series of molecules with Neu5Ac at the non-reducing termini display almost exclusively fragment ions that contain that residue. This result is contrasted with the fragmentation of [M–H] asialo carbohydrates and glycoconjugates, in which reducing and non-reducing terminal fragmentation is observed (Domon & Costello, 1988b). Fragmentation to the reducing terminal side of HexNAc residues is facile in the product ion spectra of [M–H] ions (Peter-Katalinic, 1994). Interestingly, when Neu5Ac is present as a substituent of a mid-chain Gal MASS SPECTROMETRY OF OLIGOSACCHARIDES residue of a glycosphingolipid, most of the fragment ions contain the acidic residue, indicating that fragmentation centers around this locus (Egge & Peter-Katalinc, 1987; Costello & Vath, 1990). By contrast, ionization of sialylated glycoconjugates (glycosphingolipids) as [M þ H]þ ions results in the preferential elimination of Neu5Ac (Egge & Peter-Katalinc, 1987), and less informative mass spectra. Permethylated glycosphingolipids fragment to produce a pattern that identifies the presence of Neu5Ac, but may not unequivocally locate the substitution position due the high abundance of ions that correspond to the oxonium ion (Costello & Vath, 1990). Oligosaccharides that differ only in the linkage position of sialic acid groups may be differentiated based on high-energy CID patterns. The natriated permethylated derivatives of the pentasaccharide Neu5Ac(2–6)Gal(1– 4)GlcNAc(1–3)Gal(1–4)Glc produce abundant ions that contain the nonreducing terminus and those that contain the reducing terminus in low abundance (Lemoine et al., 1991). The permethylated isomer with (2–3) linked Neu5Ac, on the other hand, produces a pattern in which the most abundant product ions contain the reducing terminus. These observations suggest that the Neu5Ac(2– 6)Hex linkage results in a more stable sodium adduct, resulting in charge-remote fragment ions that contain the non-reducing terminus. The CID product-ion spectra of natriated permethylated oligosaccharides was, therefore, seen as an alternative to that of protonated ions that had the potential to provide additional sensitivity to the linkage structure. E. Conclusions For MS profiling of glycoconjugates and oligosaccharides, it is important to consider that neutral and acidic molecules will produce different ionization responses by MALDI or ESI. In cases where it is desirable to simultaneously obtain a mass profile of all components, chemical modifications are necessary to obtain even ionization responses for all components. Methyl esterification of sialic acid residues renders them neutral, can be accomplished on a small sample quantity (i.e., from glycans released from SDS PAGE gel spots), and is applicable to MALDI and ESI. Alternatively, the mixture of neutral and acidic molecules can be analyzed in the negative MALDI mode, using an anion dopant to equalize the ionization responses. For many underivatized sialylated glycoconjugate or oligosaccharide mixtures, however, the complexity is such that fractionation is required before acquiring mass spectra to avoid overlapping peaks. Ion-exchange separation of neutral and acidic molecules is straightforward and meets the need of mixture simplification. After separation, the neutral molecules may be analyzed with positive-mode MALDI or ESI and the acidic molecules in the nega- & tive mode. Alternatively, the acidic pool can be methyl esterified to facilitate its analysis in the positive mode. Another approach is to permethylate both pools and analyze them with MALDI or ESI with or without reversed-phase fractionation. As described in Figure 22, the choice of derivatization procedure depends on the sample quantity. Methyl esterification is recommended when analyzing glycan isolated from gel spots. VI. MASS SPECTROMETRY OF SULFATED OLIGOSACCHARIDES Sulfated oligosaccharides have historically posed particular analytical challenges due to their acidity and lability. Progress in the mass spectrometric analysis of sulfated glycoconjugates has lagged behind that of other compound classes, with serious implications for the understanding of the biological activities of those structures. Because of those challenges, the uses of mass spectrometry in the analysis of sulfated carbohydrates have been limited in scope. A. Derivatization Sulfate groups are not compatible with the solid sodium hydroxide method of permethylation (see sections III.A and VIII.A) due to their lability under basic conditions and difficulties associated with the extraction step. Those molecules must be permethylated with the Hakomori method (Hakomori, 1964; Dell et al., 1994), a procedure that leaves the sulfate groups underivatized. The molecules may be analyzed directly, or subjected to further derivatization to produce fully derivatized glycans. In the latter scheme, the sulfate groups are chemically hydrolyzed, and the free hydroxyl groups formed are permethylated with trideuteromethyl iodide with the solid sodium hydroxide method (Viseux, de Hoffmann, & Domon, 1998; Viseux, Costello, & Domon, 1999). The positions of the sulfate groups are indicated by the masses of the trideuteromethyl groups. In addition, the glycans are no longer acidic, and will ionize equally well with permethylated derivatives of neutral and sialylated molecules, enabling quantitative mass spectrometric profiling; see Section V.C. B. Ionization Methods 1. Fast Atom Bombardment By nature, sulfated oligosaccharides are easily ionized on account of the acidity of the sulfate groups. Nonetheless, this class has proven difficult to analyze because of the fragility of the sulfate groups during the ionization and dissociation processes. As summarized in Table 7, FAB has 199 & ZAIA TABLE 7. Summary of the use of FAB MS for the analysis of sulfated oligosaccharides (Continued) 200 MASS SPECTROMETRY OF OLIGOSACCHARIDES & TABLE 7. (Continued) a Abundant ions corresponding to losses of 102 u (NaSO3) from the precursor ion were observed. Ion produced from glucosidic-bond cleavages observed in the FAB mass spectrum. c A mixture of natriated precursor ions was observed corresponding to [Mn(Na)–H]. d No losses of 80 u (SO3) or 102 u (NaSO3) were observed from the precursor ion. e Derivatized with a lipid group using reductive amination. f Abundant losses of 80 u (SO3) were observed from the precursor ion. g A glycosphingolipid. h FAB MS of a mixture of peracetylated oligosaccharides in which the sulfate group is not modified. i The oligosaccharide was permethylated, leaving the sulfate groups unmodified. j Losses of KSO3 from the precursor ion were observed. b been used to ionize sulfated carbohydrates with sensitivities in the 5-50 nmol range. Typically, the molecules are observed as sodium adducts; the number of associated sodium ions increases with the number of sulfate groups. For a monosulfated disaccharide, it was possible to observe ions in the positive and negative modes (Carr & Reinhold, 1984). In the positive mode, a mixture of [M(nNa) þ H]þ ions were detected, where n ¼ 2, 3. Abundant ions that corresponded to the loss of 102 u, which corresponded to the loss of NaSO3 with proton replacement, were observed in addition to glycosidic-bond cleavages. In the negative mode, [M–H] and [M(Na)–H] were observed, with no losses of 102 u, in addition to ions that corresponded to glycosidic bond cleavages. For disulfated tetramers, losses of 102 u were observed in the negative mode in addition to glycosidic bond cleavages. Those data demonstrate that the FAB ionization process is energetic enough to break covalent bonds in carbohydrates, and that glycosidic bond fragmentation and losses of NaSO3 occur under similar conditions. Work on monosulfated mucin oligosaccharides shows that positive ionization results in [M(Na) þ H]þ ions with concomitant losses of NaSO3 with proton replacement (Mawhinney & Chance, 1994). Similar results are obtained from disulfated disaccharides in the positive FAB ionization mode (Reinhold et al., 1987). Singly sulfated mucin oligosaccharides may be detected in the negative FAB ionization mode as [M–H] ions, where glycosidic-bond cleavages are observed with no losses of sulfate-containing moieties (Strecker et al., 1987). Disulfated mucin oligosaccharides may be observed as [M(Na) þ H]þ ions in the positive mode, where losses of 102 u and glycosidic bond cleavages are observed. The ionization of polysulfated oligosaccharides can be accomplished in the negative ion FAB mode, resulting in polysulfated protonated molecule ions, losses of NaSO3, and glycosidic bond cleavages (see Table 7 for references). The FAB sensitivity of sulfated oligosaccharides can be enhanced through permethylation or peracetylation (Dell et al., 1988; Khoo et al., 1993; Karlsson, Karlsson, & Hansson, 1996), although losses of NaSO3 are still observed. Polysulfated oligosaccharides as lipid conjugates can be ionized at [M–H] ions, resulting in losses of 80 u (SO3), in addition to glycosidic-bond cleavages (Chai, Rosankiewicz, & Lawson, 1995). In some cases, it is not possible to determine whether losses correspond to 80 or 102 u, due to the presence of a distribution of [M(nNa)]– H] ions (Reinhold et al., 1987; Dell et al., 1988; Mallis et al., 1989; Linhardt et al., 1992). In other cases, it is clear that a loss of 102 u occurs from the protonated molecule ion (Mawhinney & Chance, 1994). For the purposes of this discussion, losses observed in the spectra of polysodiated 201 & ZAIA sulfated oligosaccharide protonated molecule ions will be considered to correspond to 102 u, NaSO3 with proton replacement. A tetrasulfated CS octamer was desalted against ammonium salts with gel filtration, and was analyzed with FAB MS in the negative mode, resulting in an ion that corresponded to [M–3(SO3)–H] (Chai, Kogelberg, & Lawson, 1996). The addition of increasing quantities of NaHCO3 to the sample resulted in successively decreasing abundances of ions produced from losses of NaSO3. Ultimately, it was possible to observe a series of [M(nNa)–H] ions, where n ¼ 3–6, with no losses of NaSO3 or SO3. Those results are consistent with the conclusion that pairing sulfate groups with sodium ions increases their stability during ionization. 2. MALDI a. MALDI of Sulfated Peptides. The Use of MALDI for the analysis of sulfated peptides is reviewed here because the findings are instructive for the analysis of sulfated oligosaccharides. Early studies on sulfated peptides with FAB ionization suggested that positive ions generated from those molecules undergo facile losses of SO3, with the result that the peptide may not appear to be sulfated (Gibson & Cohen, 1990). Monosulfated peptides are observed as [M–H] ions in the negative FAB ionization mode, indicating its use for such molecules. Disulfated peptides, however, are observed as a mixture of [M–H] and [M–SO3 –H] ions by negative FAB ionization (Yagami, Kitagawa, & Futaki, 1995). For such ions, one of the sulfate groups is protonated and one deprotonated, and the results are consistent with the loss of the deprotonated sulfate group during the ionization process. It is possible to detect a sulfated peptide in the negative mode with MALDI-TOF MS as an [M–H] ion, and to observe a combination of [M–SO3 þ H]þ and [M þ H]þ in the positive mode (Talbo & Roepstorff, 1993; Zaia et al., 2001), an effect that can be used to identify sulfated peptides in digest mixtures. The degree to which this metastable decay occurs is dependant on the mass spectrometer source design and on the MALDI matrix used. Although, when using a conventional MALDI-TOF source in the positive mode, no [M þ H]þ ion was detected for a sulfated conotoxin peptide, the use of atmospheric-pressure MALDI ionization resulted in the observation of that ion in addition to those that resulted from SO3 loss (Wolfender et al., 1999; Laiko, Baldwin, & Burlingame, 2000). In the linear negative mode, however, the [M–H] ion was 5–10fold more abundant than the [M–SO3 –H] ion. In the negative reflectron mode, the two ions were equal in abundance, showing that metastable fragmentation occurs to a significant degree. The analysis of sulfated peptides is, therefore, best undertaken in the negative MALDI mode, and the use of atmospheric pressure sources is also war202 ranted. Using atmospheric-pressure MALDI on an ion trap, ions that correspond to the loss of SO3 were observed in the positive mode, and intact [M–H] ions were observed in the negative mode (Moyer et al., 2002). b. Direct MALDI of Sulfated Oligosaccharides. MALDI-TOF MS has been used to detect monosulfated mucin tetrasaccharide alditols as [M(2Na) þ H]þ ions with either 2,5-dihydroxybenzoic acid or 3-aminoquinoline matrices (LoGuidice et al., 1997). The result implies that monosulfated carbohydrates are amenable to MALDI ionization in the positive mode in the sodium-adducted form. In-gel PNGase F digestion has been used to release N-linked glycans from bovine thyroid-stimulating hormone (Wheeler & Harvey, 2001). The neutral oligosaccharides were separated from the sulfated and sialylated oligosaccharides with a porous graphitized carbon column, and the fractions were analyzed with MALDI-TOF MS. The acidic fractions were analyzed with D-arabinosazone as a matrix, and the spectra exhibited abundant [M þ Na]þ ions that correspond to mono- and di-sulfated biantennary N-linked structures. Significantly, several ions that correspond to sulfated glycans were observed with no losses of 80 u (SO3) or 102 u (NaSO3 with proton displacement), indicating that metastable decay does not occur under the conditions used. Sulfated N-linked glycans of bovine peripheral myelin have been analyzed with a trihydroxyacetophenone matrix in the negative MALDI mode (Gallego et al., 2001). The results showed that a mixture of [M–H] and [M(Na)–H] ions were observed with little or no losses of 80 or 102 u. Sulfated N-linked glycans have also been analyzed effectively by negative ion MALDI-TOF with a DHB matrix (Zamze et al., 2001). Chondroitin sulfate solutions digested with chondroitinase ABC or hyaluronidase have been analyzed by MALDI-TOF with a DHB matrix (Schiller et al., 1999). The results for those polysulfated oligosaccharides are similar to those obtained on sulfated oligosaccharides with FAB ionization in that losses of 102 u (NaSO3 with proton displacement) are observed from many of the ions in the spectra. The results are ambiguous because they are consistent with either the presence of a substantial population of undersulfated oligosaccharides in the enzymatic digest mixtures or mass spectrometric fragmentation of the ions. To summarize, traditional low-pressure MALDI can be used for monosulfated and, provided care is taken to use cation adduction, disulfated oligosaccharides. With more than two sulfate groups present, the use of MALDI is not recommended due to abundant metastable losses. Highpressure MALDI has been used to suppress fragmentation of fragile molecules such as polysialylated gangliosides (O’Connor & Costello, 2001; O’Connor, Mirgorodskaya, & Costello, 2002). The extent to which high-pressure MASS SPECTROMETRY OF OLIGOSACCHARIDES & MALDI may be useful to analyze polysulfated carbohydrates is not clear at this time. c. Use of Basic Peptides for MALDI of Polysulfated Oligosaccharides. Polysulfated carbohydrates are detected only in very low abundance with MALDI-TOF, with extensive losses of SO3 and HSO4 (Juhasz & Biemann, 1995). A vastly improved signal is obtained when the polysulfated oligosaccharide is mixed 1:1 with a basic peptide of sequence (RG)n, with abundant peaks that correspond to the complex. The abundance of the ions produced from losses of SO3 were reported to be dependent on the matrix used; the use of 3-hydroxypicolinic acid (3HPA) results in very low abundances for those losses. Further, it was shown that equimolar mixtures of a heparin oligosaccharide and a basic protein, such as angiogenin, produce ions that correspond to the complex in MALDITOF mass spectra acquired from 3-HPA. That work led to marked progress in the analysis of heparin sequences by determining the molecular weight of complexes before and after chemical or enzymatic degradation. Complexes between heparin oligosaccharides and the peptide (RG)19R were ionized from caffeic acid, and the data used to show that the heparin-degrading enzyme heparinase II acts by an endolytic mechanism (Rhomberg et al., 1998). The same conditions were used to show that heparinase I cleaves by a predominantly exolytic mechanism (Ernst et al., 1998). Derivatization of heparin oligosaccharides to reducingterminal semicarbazones has been used to help distinguish products of enzymatic degradation that contain the original reducing- and non-reducing-termini (Rhomberg et al., 1998). Figure 30 illustrates the use of basic peptide complexation to monitor the course of a heparin-lyase digestion of a mixture of heparin-like glycosaminoglycan (HLGAG) oligosaccharides as a function of time (Rhomberg et al., 1998). Sucrose octasulfate (SOS) was used as an internal standard, relative to which the abundances of the HLGAG oligosaccharides were measured to obtain a semi-quantitative digestion profile. HLGAG oligosaccharides H1 and T5 with SOS were digested with heparin lyase I, and aliquots were removed at time points and diluted with matrix solution to stop the reaction. Caffeic acid was used as the matrix, and the solution contained (RG)15 at an approximately 2:1 molar excess over HLGAG oligosaccharides. The results are impressive in that the degradation of H1 to produce T7 can be followed with a relatively simple procedure that involves no derivatization or chromatographic steps. The masses of the HLGAGs are determined by subtracting the observed m/z value of the peptide [M þ H]þ ion (labeled P in Figure 30) from those observed for the peptide-HLGAG complexes. Those results are significant because they show that MALDI-TOF, with its ease of sample handling and well- FIGURE 30. (A) MALDI mass spectrum of a mixture of substrates H1 and T5, as well as sucrose octasulfate (SOS), after a 40-min digestion with heparinase I. (B) Peak ratios of oligosaccharides H1, T5, and T7 relative to SOS during a 2-hr heparinase I digest. Reprinted with permission from Rhomberg et al. (1998)). Copyright 1998 National Academy of Sciences, USA. developed instrumentation, can be used in the mass analysis of HLGAGs. The technique is limited because the use of a complexing peptide requires a great deal of skill and patience, making the experiment time-consuming. In addition, the fact that the charge is carried by the complexing peptide makes it impossible to conduct CID of the HLGAG ions. To gain sequence information requires the use of chemical or enzymatic degradation and the use of pure oligosaccharides. It is possible to mass-analyze HLGAGs without the use of complexing molecules with negative ESI (Chai et al., 1998; Pope et al., 2001) and to obtain sequence information with CID (Zaia & Costello, 2003), see Section VI.C.6.b. A detailed notation system to represent HLGAG sequences has been described, and was used in combination with chemical and enzymatic degradation steps to determine sequences of oligosaccharides involved in anticoagulation, cell growth, and differentiation (Venkataraman 203 & ZAIA et al., 1999b). That approach was used to sequence a 3-Osulfate-containing antithrombin III-binding HLGAG decamer with a caffeic acid matrix and (RG)19R as the basic peptide (Shriver et al., 2000). It was also shown with MALDI-TOF MS that glucosamine 3-O sulfation at the reducing end of a glycosidic linkage imparts resistance to glycosidic cleavage by heparinases I, II, and III (Shriver et al., 2000). Recently, MALDI-TOF MS combined with the semicarbazide derivatization scheme and heparinase digestion have been used in the structural characterization of a 3-O-sulfated octasaccharide that binds to herpes simplex type I glycoprotein D (Liu et al., 2002). The use of basic-peptide complexation is also applicable to the analysis of a complex low molecular weight heparin fraction that consisted of hexamers and octamers with varying numbers of sulfate groups (Sturiale, Naggi, & Torri, 2001). d. MALDI Analysis of Protein-Sulfated Oligosaccharide Complexes. The addition of a heparin decasaccharide to solutions of fibroblastic growth factor (FGF) types 1 or 2 results in marked changes to the MALDI-TOF mass spectra obtained from those proteins (Venkataraman et al., 1999a). It was possible to detect complexes between the protein and decamer, and no such complexes were observed for a heparin hexamer. Whereas FGF-1 forms 1:1 and 2:1 protein:decasaccharide complexes with MALDI-TOF MS, FGF-2 forms 1:1, 2:1, 3:1, and 4:1 complexes under the same conditions. HLGAG tetrasaccharides have been shown to be sufficient to induce FGF-1 dimerization, and the effects of different protein-ligand ratios on complex formation have been shown (Sturiale, Naggi, & Torri, 2001). It has recently been demonstrated with MALDITOF MS that FGF-1 dimerization requires only one 6-Osulfate group heparin tetrasaccharide (Guerrini et al., 2002). Those results demonstrate the utility of MALDI for the analysis of growth factor-heparin complexes, and produce important new data to provide insight on FGF biochemistry. Figure 31 shows a MALDI-TOF MS study of the stoichiometry of the binding of a growth factor (fibroblastic growth factor type 2, FGF2) with its receptor (FGF receptor, FGFR2) as a function of the presence of HLGAGs (Kwan et al., 2001). FGF, FGFR2, and a heparin lyasederived oligosaccharide were mixed in 10 mM sodium phosphate buffer, and were allowed to equilibrate at 48C for 30 min. The sample was spotted onto the MALDI target with 1 mL sinapinic acid solution (saturated in 50% acetonitrile). After drying, the sample was washed with water. In the absence of HLGAG, FGF and FGFR2 both form complexes with a 1:1 stoichiometry (Fig. 31a). In the presence of HLGAG, a detectable 2:2:1 FGF:FGFR2: HLGAG complex is formed, indicating that the oligosaccharide mediates formation of a unique binding stoichiometry (Fig. 31b). The inset shows the effect of the addition 204 FIGURE 31. Competitive binding of dFGF2 to FGFR2. (A) MALDI mass spectrometric profile of a mixture of wild-type FGF2 and the ectodomain of FGFR2. Observed in the mass spectrum are the [M þ H]þ ions for an FGF2 dimer (m/z 30,214) and trimer (m/z 45,132), and FGFR2 monomer (m/z 24,888) and dimer (m/z 49,572), and a 1:1 FGF2FGFR2 complex (m/z 39,896). The theoretical molecular masses for FGF2 and FGFR2 are 15,114 and 24,864 Da, respectively. (B) Mass spectrum of the FGF2/FGFR2 mixture in the presence of a homogeneous HLGAG decasaccharide. Addition of a decasaccharide (Deca) to FGF2/FGFR2 promoted the formation of a 2:2 FGF2FGFR2 complex with an observed [M þ H]þ ion at m/z 82,750 (with the decasaccharide) or m/z 79,872 (without the decasaccharide). The [M þ H]þ ions for two dimeric FGFR2 species were also observed. The first (at m/z 49,592) represents the apo complex, and the second (at m/z 52,474) is a 2:1 FGFR2decasaccharide complex. Inset: Mass spectrum of dFGF2 added to the decasaccharide/ FGF2/FGFR2 mixture shown above. Three high molecular mass complexes were observed: a 2:2 FGF2FGFR2 complex with or without the decasaccharide and a 1:2 dFGF2FGFR2 complex without the decasaccharide. Reprinted with permission from Kwan et al. (2001). Copyright 2001 American Society for Biochemistry and Molecular Biology. of a genetic construct of FGF (dFGF) that corresponds to a dimerized molecule with a histidine tag. The dFGF construct binds to two molecules of FGFRs in the presence of the HLGAG, and forms a peak at slightly higher mass than observed for the 2:2:1 FGF:FGFR2:HLGAG complex. The data were used to support the conclusion that HLGAGs facilitate FGF and/or FGFR2 oligomerization. The results show that MALDI-TOF can be used to determine multiprotein binding stoichiometries. It is interesting that the matrix conditions would be expected to weaken complexes and that, nonetheless, complexes are observed. Although it MASS SPECTROMETRY OF OLIGOSACCHARIDES may be possible to obtain more biologically relevant results with nano ESI and aqueous, neutral pH conditions, such an experiment is more technically challenging than the MALDI-TOF MS. 3. ESI of Sulfated Oligosaccharides Desalted CS oligosaccharides have been analyzed from a 0.5 nM ammonium acetate solution with negative mode ESI (Takagaki et al., 1992), where the most abundant charge state for the oligosaccharide ions corresponded to one charge per sulfate group. For example, an abundant [M–4H]4 ion was observed for the tetrasulfated CS octamers with [M–3H]3 and [M–5H]5 ions in much lower abundances. Ions that correspond to losses of SO3 were in very low abundances, and it was possible to analyze up to CS tetradecasaccharides. The same conditions were used to obtain negative ESI of disulfated Lewisx trisaccharides, which results in abundant [M(Na)–H] and [M– 2H]2 ions with no losses of SO3 (Ii et al., 1995). Heparin oligosaccharides generated by heparin-lyase digestion have been analyzed with negative ESI as ammonium salts (Chai et al., 1998). Gel filtration-purified oligosaccharides were infused in organic solvent with or without 0.5 mM ammonium bicarbonate. In the absence of ammonium ions, the mass spectra of heparin oligosaccharides were quite complex due to multiple adduction with sodium ions. In the presence of ammonium ions, the profiles were much less complex, due to a marked reduction in the abundances of ions that correspond to sodium adducts of the oligosaccharides. It was possible to acquire data on heparin fractions up to a decasaccharide size and of CS oligosaccharides up to tetradecamer size (Beeson et al., 1998; Chai, Beeson, & Lawson, 2002). Negative-ion ESI mass spectra were acquired on chromatographically purified oligosaccharides derived from acharan sulfate, a glycosaminoglycan from the giant African snail that is related to heparin and HS (Kim et al., 1998). The spectra were acquired from 50% acetonitrile, 0.05% ammonium hydroxide, and showed abundant ions that correspond to [M–2H]2 and [M(Na)–H]2 for the disulfated tetrasaccharide fraction. The degree of sodium adduction was observed to decrease as the concentration of the analyte was decreased, and to increase with the size of the analyte, and data were acquired on up to octasaccharides. The same conditions have been used to analyze synthetic heparin oligosaccharides (Yu et al., 2000) and those derived from dermatan sulfate (Yang et al., 2000). Nano-ESI MS (Wilm & Mann, 1994) has been applied to the analysis of synthetic heparins, using a modification of the above conditions (Pope et al., 2001). The oligosaccharides were sprayed from a solution of 0.2% ammonium hydroxide, 0.2% imidazole, and 50% acetonitrile from pulled glass capillaries, using a titanium wire to provide & electrical contact. The results showed that abundant deprotonated ions are produced; those ions that corresponded to sodium, potassium, or imidazole adduction had a lower abundance. Although it is clear that sulfated oligosaccharides may be ionized without in-source fragmentation with negative ESI, the use of basic peptides is an alternative method. Polysulfated carbohydrates such as sucrose octasulfate were mixed with a basic peptide in 1:5 (carbohydrate:peptide) molar ratios, and negative-ion ESI mass spectra were acquired from 50% acetonitrile solutions (Siegel et al., 1997). The results for sucrose octasulfate in the absence of basic peptide showed a complex pattern of ions that resulted from varying degrees of sodiation and desulfation. In the presence of basic peptide, abundant ions that corresponded to [M þ peptide-4H]4 and [M þ peptide– 3H]3 ions were observed with natriated ions in very low abundance and no in-source fragmentation of sulfate groups. A number of peptides that contained a high percentage of lysine and arginine residues were found to form complexes, the choice of which was found empirically for each polysulfated carbohydrate tested. Recently, the analysis of sucrose octasulfate (SOS) has been accomplished in the positive ESI mode through the use of counterions in the spray solvent (Gunay et al., 2003). The results show that, although a clear [M(8Na) þ Na]þ ion is observed in the positive mode with no fragmentation, extensive fragmentation was observed for the [M(7Na)– H] ion in the negative mode. Note that, using this nomenclature, the protons displaced by the cation in parentheses is understood, see Section II.B. Pairing of SOS with tetramethylammonium cation resulted in the detection of [M(8(CH3)4N) þ (CH3)4N]þ, in which all eight sulfate groups are deprotonated and paired with a cation, and an additional cation provides the net positive charge. In the negative mode, an ion that corresponds to [M(7(CH3)4N)– H] was detected in addition to abundant fragment ions. Similar trends were observed for SOS paired with tetrapropylammonium and tetramethylphosphonium cations. The best results for ionization of SOS were obtained in 10 mM tetraethylammonium hydroxide, resulting in the observation of a single ion that corresponded to [M(8R) þ R]þ in the positive mode and [M(7R)–H] in the negative mode, where R ¼ tetraethylammonium cation. The sensitivity was 10-fold higher in the positive mode. Those results demonstrate that sulfate groups may be stabilized with cations to allow their detection as singly charged ions with ESI. Although it is not clear at this time why more fragmentation is observed in the negative mode, it will be important to study the fragmentation patterns that result from CID of those ions. The use of ESI to analyze of polysulfated oligosaccharide is recommended because it results in little or no insource fragmentation—provided that a low desolvation 205 & ZAIA energy is used. MALDI is considerably more energetic, and is, therefore, applicable to molecules for which the sulfate groups are relatively stable: negatively charged monosulfated oligosaccharides or sodium-adducted disulfated oligosaccharides (Wheeler & Harvey, 2001). 4. On-Line Separation Systems for Sulfated Carbohydrates An interface has been optimized to allow direct capillary electrophoresis measurements on oligosaccharides produced from enzymatic digestion of heparins (Duteil et al., 1999). Separations were performed using either ammonia/ ammonium acetate pH 9.2 or acetic acid/ammonium acetate pH 3.5; both are compatible with ESI. The two conditions resulted on a reversal of the elution order for a series of 8 heparin disaccharides. Capillary electrophoresis was coupled with negative ESI by using a sheath liquid of 2 mM triethylamine, 3 mM ammonium formate in 50% acetonitrile, resulting in the observation of abundant deprotonated protonated molecule ions, no in-source fragmentation, and low-abundance triethylamine adducts. On-line CE/MS data were acquired for heparin disaccharide and tetrasaccharide mixtures from a heparin-lyase digestion. Pressure-assisted capillary electrophoresis has been interfaced with an ESI ion trap mass spectrometer, using a sheath liquid of 5 mM formic acid pH 3.20 in 2-propanol (Ruiz-Calero et al., 2001). The capillary electrophoresis electrolyte consisted of 30 mM formic acid, pH 3.2, and the separation was assisted with 0.5 psi of nitrogen gas. Using those conditions, it was possible to resolve the eight heparin disaccharides in a 13-min electrophoretic separation, and it was possible to analyze disaccharides produced from enzymatic depolymerization of heparins. Off-line CE has been used to separate oligosaccharides produced from the chondroitinase digestion of chondroitin sulfate samples for ESI MS and MS/MS analysis (Zamfir et al., 2002). On-line LC/MS has been used to analyze sulfated N-linked oligosaccharides, using a graphitized carbon column (Kawasaki et al., 2001). The oligosaccharides were eluted with a water/acetonitrile gradient in the presence of 5 mM ammonium acetate, and were detected with negative or positive ESI. An ion-pairing reversed-phase chromatographic method has recently been described for on-line LC/ MS of heparins as well as other acidic carbohydrates (Kuberan et al., 2002). Dibutylamine is used in the mobile phase in the presence of acetic acid, where dibutyammonium ions pair with the negatively charged carbohydrates to allow fractionation on a C-18 column. A mixture of deprotonated and dibutylammonium-adducted ions were observed with these conditions to analyze a heparin pentasaccharide. Significantly, no sulfate losses were observed. That technique seems appropriate for on-line LC/MS 206 determination of sulfated oligosaccharide masses, and is appropriate for miniaturization to maximize sensitivity. It should be noted that this method is most appropriate when an instrument can be dedicated to HLGAG analysis due to difficulties in removing the ion-pairing agent. Size-exclusion chromatography (SEC) has been used on-line with ESI MS to desalt and separate mixtures of sulfated GAG oligosaccharides (Zaia & Costello, 2001). A commercial column (Amersham Biosciences Superdex Peptide) is available that separates di-, tetra-, hexa-, and octa-saccharides in a 20-min time-period. The method is quite convenient because contaminating salts are effectively removed; at the same time, the complexity of the oligosaccharide mixture that enters the instrument at any moment is limited. Solvent conditions of 0.1 M ammonium acetate, 10% methanol have been found to work for all types of sulfated GAG oligosaccharides. C. Tandem MS of Sulfated Oligosaccharides 1. Lessons from CID of Sulfated Peptides High-energy tandem mass spectrometric analysis of tyrosine-sulfated peptides as [M–H] ions with FAB ionization results in abundant product ions from peptidebackbone cleavage and an ion that corresponds to [M– SO3 –H] in moderate abundance (Gibson & Cohen, 1990). A sulfated an ion is observed that definitively places the sulfated tyrosine group in the context of the peptide sequence. It was also reported that losses of SO3 from the precursor are relatively more abundant than losses of HPO3 from peptides that contain phosphotyrosine. Using positive FAB ionization, ions produced from the loss of all sulfate groups form the base peaks in CID mass spectra generated from peptides that contain multiple tyrosine-sulfate residues (Yagami, Kitagawa, & Futaki, 1995). In the negative FAB mode, a trisulfated peptide produced a ladder of ions that corresponded to [M–H], [M–SO3 –H], and [M– 2(SO3)–H] with the absence of a [M–3(SO3)–H] ion. Those results are consistent with the conclusion that protonated sulfate groups are significantly more labile than deprotonated sulfate groups. Positively charged ions generated from phosphopeptides undergo minimal losses of HPO3 during CID on a Q-oTOF mass spectrometer with ESI (Nemeth-Cawley, Karnik, & Rouse, 2001). Under the same conditions, sulfated peptides undergo losses of SO3; the number of losses correspond to the number of sulfate groups that modify the peptide. By increasing the collision energy, it was possible to remove all sulfate groups and to induce fragmentation of the peptide backbone. The method is useful to differentiate isobaric phosphate and sulfate groups based on CID fragmentation. Carbohydrates are known to undergo backbone fragmentation at lower collision energies than peptides, and the challenge is to find MASS SPECTROMETRY OF OLIGOSACCHARIDES conditions where such ions are formed in preference over losses of SO3 so that as much structural information as possible can be produced from the tandem mass spectra. 2. Tandem MS of Mono- and Di-Sulfated Oligosaccharides From the discussion of ionization of sulfated oligosaccharides (see Section VI.B), the analysis of those molecules is best carried out with negative ESI. Using that technique, sulfated N-linked oligosaccharides are likely to ionize with one charge per sulfate group, a condition where those groups are least labile. Using negative ESI, CID mass spectra of monosulfated N-linked oligosaccharides show product ions that always contain the sulfate group. For a biantennary core fucosylated lactosamine structure, the product-ion pattern was consistent with sulfation of one of the two GlcNAc residues, as shown in Figure 32 (Geyer & et al., 2001). The results show a relatively complete series of Bm and Yn ions. Although the B5 ion is the most abundant in the spectrum, other cleavages to the reducing side of GlcNAc are absent. This result is consistent with the proton-mediated weakening of the HexNAc glycosidic bond for protonated ions. The absence of mobile protons results in the stabilization of those linkages, and allow other cleavages to occur in the MS2 stage. Sulfoglycolipids have been analyzed in negative- and positive-mode MS. Positive-mode product-ion mass spectra were not useful because the abundant losses of SO3 from the precursor ion resulted in the loss of information on the position of sulfation. In the negative mode, however, all fragment ions contained the SO3 group and were thus useful for structural determination (Kushi, Handa, & Ishizuka, 1985; Egge & Peter-Katalinc, 1987). Useful fragmentation was also obtained from CID of [M–H] ions generated from disulfated glycosphingolipids. FIGURE 32. Negative-ion nano-ESI-IT-MS spectra of a sulfated N-linked glycan present on neural cell adhesion molecule from newborn mouse brain. (A) MS2 profile of the [M–H] ion at m/z 1944.0; (B) MS3 profile of the Y5b ion at m/z 1781.7 (Geyer et al., 2001). 207 & ZAIA 3. Precursor-Ion and Neutral-Loss Scans for Sulfated Glycoconjugates The sphingolipid sulfatide consists of a 3-sulfogaltosyl group linked to a ceramide nucleus. Product-ion tandem mass spectra of these species with negative ESI showed the formation of structurally significant product ions in the absence of losses of SO3. In addition, an ion at m/z 97, [HSO4], was observed in all sulfatide product-ion spectra (Marbois et al., 2000). Precursor-ion scans for m/z 97 were used to identify ions produced from brain extracts that contained sulfate and that were likely to correspond to sulfatide (Hsu, Bohrer, & Turk, 1998). Precursor-ion scans have been used to identify and quantify glycosphingolipids from kidney tissue (Sandhoff et al., 2002). Significantly, the m/z 97 ion characterizes O-sulfate groups and is absent for sulfone groups (giving rise to .SO3, m/z 80). Quantification was linear over more than two orders of magnitude, a range that was 70-times greater than that for densitometric scanning. Precursor-ion scanning has been used in the structural analysis of sulfated N-linked oligosaccharides (Kawasaki et al., 2001). In that work, a complex mixture of N-linked oligosaccharide alditols released from erythropoietin was analyzed with negative-ion triple-quadrupole ESI LC/MS with graphitized carbon chromatography. Sulfated oligosaccharides were identified from mass chromatograms of m/z 1035 (sulfated triantennary, trisialylated, 3-), m/z 1254 (sulfated tetraantennary tetrasialylated, 3-), m/z 1281 (disulfated tetraantennary, tetrasialylated, 3-), and other sulfated N-linked ions. The results provide useful information on the presence of sulfated species in the mixtures, and show the value of negative-ion ESI for the analysis of sulfated oligosaccharides. Because the sulfate groups are charged, they are stable and can be analyzed with confidence. Sulfated peptides generated by the tryptic digestion of recombinant factor VIII produce abundant doubly charged ions in positive-ESI reversed-phase LC/MS analyses (Severs et al., 1999). Detected m/z values for several ions were consistent with modification with up to three sulfate groups. Scans for precursor ions that lose SO3 (80 Da) were used to identify peptides that eluted from the column that are modified with sulfate. Because the ions were doubly charged, this identification entailed scanning for a 40 u neutral loss. 4. Determination of Positional Sulfation Isomers in GAG Disaccharides Negative-ion FAB CID mass-analyzed ion kinetic-energy spectra (MIKES) have been used to analyze monosulfated disaccharides generated from CS (Lamb et al., 1992). As summarized in Table 8, the results show that [M–H] ion 208 generated from those molecules undergo fragmentation to produce abundant ions from glycosidic-bond and crossring cleavages with no ions that correspond to losses of SO3. The residue location of the sulfate group could easily be determined by the m/z values of the ions produced from glycosidic-bond cleavage. Mass spectrometric differentiation of positional sulfation isomers was demonstrated for [M–H] ions of isomeric disaccharides that differ only in the position of sulfation on the GalNAc residues, DUA(1,3)GalNAc4S and DUA(1,3)GalNAc6S. The MIKES spectra showed that the 4-sulfated isomer produced an abundant ion at m/z 300 that corresponded to Y1, and that an abundant ion at m/z 282 that corresponded to Z1 was produced from the 6-sulfated disaccharide. The product-ion spectra are very distinct in that the Z1 ion is absent for the 4-sulfated isomer and the Y1 ion is absent for the 6-sulfated isomer. MIKES spectra of [M–H] ions generated from DUA2S(1,3)GalNAc and DUA2S(1,4)GlcNAc were very similar. Spectra of DUA(1,3)GalNAc6S and DUA(1,4)GlcNAc6S were quite distinct, the latter produced an abundant Y1 ion in addition to cross-ring cleavage ions that are absent in spectra of the former. It is significant that MIKES spectra of the [M(Na)–H] ions generated from those disaccharides produced different product-ion patterns. DUA(1,3)GalNAc4S and DUA(1,3)GalNAc6S both produced abundant Z1 and Z1(Na) ions, and the two isomers differ in the extent to which cross-ring fragmentation occurs. Positive ions generated from sulfated GAG disaccharides with FAB have also been analyzed with tandem MS (Ii et al., 1994). As summarized in Table 9, for [M(2Na) þ H]þ ions, it was not possible to distinguish DUA(1,3)GalNAc4S from DUA(1,3)GalNAc6S based on the product-ion pattern. The differences between DUA2S(1,3)GalNAc and the other two isomers was not very distinct due to abundant losses of SO3 from B1 and C1 ions. Those three compounds were easily distinguished, however, from tandem mass spectra generated from [M(3Na) þ H]þ ions. An ion that corresponded to Z1(2Na) was much more abundant in the spectrum of DUA(1,3)GalNAc6S than in DUA(1,3)GalNAc4S, and abundant B1(3Na) and C1(3Na) ions distinguished the DUA2S(1,3)GalNAc isomer. Abundant losses of 98 u (H2SO4) were observed from the latter ions. The disulfated isomers DUA2S(1,3)GalNAc4S and DUA2S(1,3)GalNAc6S could not be distinguished from CID of [M(3Na) þ H]þ ions, and losses of SO3 from the Y1(3Na) made it difficult to distinguish the DUA(1,3)GalNAc4S,6S isomer (see Table 9). All three disulfated isomers, however, were easily distinguished by CID of the [M(4Na) þ H]þ ions. CID product-ion spectra have proven to be generally useful to determine the number of sulfate groups that modify heparin disaccharides (Ii et al., 1995b). In MASS SPECTROMETRY OF OLIGOSACCHARIDES & TABLE 8. Summary of product ions generated from monosulfated GAG disaccharides that are useful for distinguishing isomers a Ions analyzed by MIKES. Spectra of the isomeric compounds were not distinguishable. c Ions analyzed by CID MS/MS. d Ion contains sulfate and is generated by ring cleavage. e Ions produced from subsequent fragmentation of Z1. f [Na2HSO4]þ. g [Na2HSO4]þ. b particular, it is possible to differentiate DUA2S(1,4)GlcN from DUA(1,4)GlcNS and DUA(1,4)GlcN6S by CID of [M(2Na) þ H]þ ions. As Table 9 shows, the information obtained depends on the number of sodium adducts in the precursor ion. In general, the B1 and C1 ions are observed in low relative abundance, but are useful to determine whether the DUroA residue is sulfated. Cross-ring cleavage ions 0,2 A2 and 2,4A2 are often abundant in the positive-ion CID tandem mass spectra of disulfated heparin disaccharides, are useful to assign the position of sulfation on the GlcNAc residue, and may be useful in the case of a rare GlcNS3S or GlcNAc3S sulfation pattern. 5. Tandem Mass Spectrometric Quantification of GAG Disaccharides A stereochemical differentiation of hexosamine monosaccharide residues has been demonstrated with ESI QIT MS (Desaire & Leary, 1999b). In that work, it was demonstrated that cobalt–diaminopropane complexes of the hexosamine monosaccharides glucosamine, galactosamine, and mannosamine produced unique product-ion spectra. The relative abundances of ions produced from pure standards were characterized, and were used to quantify two- and three-component mixtures. Using that 209 method, an adaptation of the superposition principle of quantifying oil distillates (Roboz, 1968), a system of equations is developed, one for each mixture component. That method has been applied to the quantification of CS disaccharides from mixtures derived from a chondroitinase digestion (Desaire & Leary, 2000). DUroAGalNAc4S (DDi-4S) produces an abundant Y1 ion at m/z 300, and DUroAGalNAc6S (DDi-6S) produces an abundant Z1 ion at m/z 282 (Linhardt et al., 1992). Using ESI QIT, DUroA2SGalNAc (DDiUA2S) was observed to produce an ion at m/z 237 that corresponded to B1 (Desaire, Sirich, & Leary, 2001). That disaccharide was used as an internal standard for the quantification of mixtures of DDi-4S and DDi-6S produced from the digestion of CS chains with chondroitinase enzymes. For that purpose, CID production mass spectra were first collected individually for the three pure disaccharides under the same instrumental conditions. The CID product-ion spectrum of a 1:1:1 DDiUA2S:DDi-4S:DDi-6S standard mixture was acquired, and the per cent total ion abundances for m/z 237, 282, and 300 were determined. The abundance of each product ion was expressed as contributions from each of the three standard disaccharides, as shown in Table 10 and Figure 33. Normalization factors (A, B, C) were calculated by substituting the observed percent total ion abundances for m/z 237, 282, and 300 in the equation. Internal standard DDiUA2S was added to unknown mixtures of DDi-4S and DDi-6S, the CID product-ion tandem mass spectra were acquired, and the percent total ion abundances for m/z 237, TABLE 10. Contributions of the three distinguishing product ions for analysis of CS disaccharides, m/z 237, 282, and 300 [reprinted with permission from Desaire, Sirich, & Leary (2001), Copyright 2001 American Chemical Society] b a Loss of SO3 and a neutral fragment, corresponding to 0,2X0 (C4H7NO2), from the precursor ion. [OHCCH2OSO3]. ZAIA TABLE 9. Summary of product ions generated from disulfated GAG disaccharides that are useful for distinguishing isomers & a The contribution is an average of four measurements. 210 MASS SPECTROMETRY OF OLIGOSACCHARIDES FIGURE 33. System of equations used to quantify disaccharides. Reprinted with permission from Desaire, Sirich, & Leary (2001). Copyright 2001 American Chemical Society. 282, and 300 were substituted in the same system of equations. The values calculated for A, B, and C were normalized with those values calculated from a 1:1:1 mixture to produce the ratios of the three disaccharides present in the mixture. That method was validated with a series of standard mixtures, and used to quantify disaccharides produced from enzyme digests (Desaire, Sirich, & Leary, 2001). Negative-ion FAB tandem mass spectrometry has been used to structurally analyze CS tetrasaccharide tri- and tetrasulfated monoclonal antibody antigenic determinants as potassium salts (Ii et al., 1995a). That work has been extended to negative-ionization ESI with QIT MS (Desaire & Leary, 2000; Desaire, Sirich, & Leary, 2001), triplequadrupole (Zaia & Costello, 2001), QoTOF (Zaia, McClellan, & Costello, 2001), and FT (McClellan et al., 2002) instruments to show that the tandem mass spectra can be used to determine sulfation ratios in mixtures produced by chondroitin lyases without any derivatization or purification. 6. Tandem Mass Spectrometric Analysis of GAG Oligosaccharides It has been shown that CID MS/MS of alkali-adducted chondroitin sulfate tetramers ionized by negative FAB results in abundant glycosidic bond cleavages with no losses of SO3 (Ii et al., 1995a). Heparin disaccharides that contain three sulfate groups can be ionized as singly charged negative ions, and fragmented by CID with abundant glycosidic-bond and cross-ring cleavages with minimal SO3 losses, provided that at least two sodium adducts are present in the negative singly charged molecular ion (Ii et al., 1995b). Singly charged trisulfated & disaccharides can also be dissociated in the positive ionization mode with tetra-sodium adducts with low to moderate losses of sulfate from the precursor ion (Li et al., 1995). Those results, and data on sulfated peptides (Yagami et al., 2000), support the conclusion that sulfate groups are least labile when charged, and that dissociation of negatively charged ions results in the greatest number of charged sulfate groups with the fewest metal-cation adducts. The fact that ESI results in more highly charged ions than observed for FAB ionization or MALDI has important implications for the analysis of highly sulfated carbohydrates. Heparin oligosaccharides generated by heparinlyase digestion can be ionized with negative ESI as multiply charged deprotonated ions from solutions that contain ammonium salts (Chai et al., 1998; Desaire & Leary, 2000; McClellan et al., 2002) without SO3 losses provided that mild desolvation conditions were used. Abundant sodium-adducted ions are observed with negative ESI in the absence of ammonium ions (Chai et al., 1998; Kim et al., 1998). Dissociation of singly charged monosulfated (Ruiz-Calero et al., 2001) or doubly charged disulfated heparin disaccharides (Pope et al., 2001) resulted in abundant ions from glycosidic-bond and crossring cleavages. Dissociation of a doubly charged, trisulfated, heparin disaccharide results in an abundant loss of one SO3 molecule from the precursor, in addition to glycosidic and cross-ring cleavages (Ruiz-Calero et al., 2001). In summary, SO3 losses are minimized and glycosidic-bond and cross-ring cleavages are maximized, when the negative charge on the precursor ion equals the number of sulfate groups. a. CS Oligosaccharides. Negative ESI of CS oligosaccharides results in a mixture of charge states, the most abundant corresponds to one charge per sulfate group (Zaia, McClellan, & Costello, 2001). To ionize GAGs, it is best to use 30% organic solutions modified with either ammonium acetate or ammonium hydroxide at a low percent (0.1%). Although the exact charge-state distribution will depend to some extent on the instrument-source design, the most abundant ion will contain one charge per sulfate group. For oligosaccharides produced by chondroitinase enzymes that act by eliminative mechanism to produce a D-unsaturated residue at the nonreducing terminus, the charge state with all sulfates charged corresponds to m/z 458. Thus, a mixture of those oligosaccharides will result in an overlapping isotopic cluster at that value. Therefore, it is necessary to either separate the oligosaccharides prior to MS analysis, or to choose a different, non-degenerate, charge state. CS oligosaccharides generated by testicular hyaluronidase, an enzyme that acts by a hydrolytic mechanism, contain a saturated non-reducing terminus, and do not produce degenerate m/z values. That enzyme, however, contains transglycosylase activity, and 211 & ZAIA its use for sequence analysis of biological samples is not advised. Tandem mass spectrometric analysis of D-unsaturated CS oligosaccharides for which all sulfates are charged produces a product-ion pattern as shown in Figure 34. Note that the abundance of the precursor ion was kept high to minimize the extent to which multiple bond fragmentation occurs. The product-ions correspond to Bm and Yn cleavages to odd-numbered glycosidic bonds, with no cleavage to even-numbered bonds. Inspection of CID product-ion mass spectra of mixtures of 4- and 6-sulfated CS octamers demonstrates that the abundance of the Bm ions depends on the position of GalNAc sulfation; see Figure 35. To gain further information on how product-ion abundances vary according to the position of sulfation, CID mass spectra were obtained on various 4- and 6- FIGURE 35. Bar-graph representations of percent total ion abundances for Bm ions produced from [M–4H]4 ions CSA/CSC octasaccharides mixtures, ratios as given: (a) tetrasulfated octamer; (b) sodium borodeuteride-reduced tetrasulfated octamer. For symbol definitions, see Figure 34. Reprinted with permission from Zaia, McClellan, & Costello (2001). Copyright 2001 American Chemical Society. FIGURE 34. Negative-ion ESI CID product-ion spectra acquired at collision energy 17.5 V, (a) CSA tetrasulfated octamer [M–4H]4 and (b) CSC tetrasulfated octamer [M–4H]4. The symbolic structure and product-ion cleavages are shown in (c). Reprinted with permission from Zaia, McClellan, & Costello (2001). Copyright 2001 American Chemical Society. 212 sulfated CS oligosaccharide mixture percentages. As shown in Figure 35, the abundance of Bm ions diminishes with the percent of 4-sulfated oligosaccharides in the mixture; those data are consistent with the abundances of those ions that indicate the sulfation position of adjacent GalNAc residues (Zaia, McClellan, & Costello, 2001). Evidently, the glycosidic bonds for 4-sulfated GalNAc oligosaccharides are more labile than are those that are 6-sulfated, resulting in more-abundant Bm ions. Therefore, it is expected that the relative abundances of those product ions will be useful in determining the position of sulfation of adjacent GalNAc residues. To prove this point, it will be necessary to synthesize CS oligosaccharides with mixed 4- and 6-sulfation in defined positions. The CS oligosaccharide product-ion pattern varies markedly with charge state, as summarized in Figure 36 (McClellan et al., 2002). For CS hexamers, D(UroA)3 (GalNAc)3(SO3)3, CID of the 2 charge state results in Bm and Yn ions from cleavage of only the even-numbered glycosidic bonds. Those results are consistent with the generation of information that is complementary to CID of the 3 charge state for hexamers. The CID product-ion MASS SPECTROMETRY OF OLIGOSACCHARIDES & FIGURE 36. The effect of charge state on the product-ion profile generated from CS oligosaccharides. The top spectrum shows the CID profile obtained from SORI CID of the [M–3H]3 ion generated from a CS hexamer, and the bottom shows that for the [M–2H]2 ion. The fragmentation diagrams are shown to the right of each spectrum (McClellan et al., 2002). profile for hexamer 1 ions results in even-numbered Bm and Yn ions, with intramolecular sulfate transfer observed. It appears that the ions with a 1 charge state may adopt a compact gas-phase conformation that allows intramolecular sulfate transfer to occur. The 2 and 3 charge states are likely to adopt an extended conformation, thus separating potential acceptor sites for an intramolecular sulfate transfer. The observed product-ion pattern for ions in which all sulfate groups are charged (top spectrum in Fig. 36) in which fragmentation occurs to the reducing side of GlcA residues is in contrast to the fragmentation observed for deprotonated HexNAc-containing oligosaccharides, in which abundant fragmentation is observed to the reducing side of HexNAc residues (see Section II.C.2). Protonated and alkali-cationized ions produced from oligosaccharides have, in fact, been observed to undergo facile cleavages to the reducing side of HexNAc residues in the positive mode for native and peralkylated ions, see Sections II.C.1, II.C.3, and II.D. The cases with low-energy CID, where facile fragmentation of HexNAc residue is not observed, are those in which a transition metal is tightly associated with the reducing end to yield charge-remote fragmentation (Konig & Leary, 1998). When CS oligosaccharide ions in which one of the sulfate groups is protonated undergo CID (bottom spectrum in Fig. 36), abundant fragmentation to the reducing side of GalNAc residues is observed that corresponds to even-numbered Bm and Yn ions (McClellan et al., 2002). Those observations may be explained based on the availability of mobile protons for ions in which one of the sulfate groups is not charged. During the multiple-collision low-energy CID process, it is likely that the –SO3H proton migrates to a GalNAc nitrogen atom, and is, therefore, in close proximity to the glycosidic bond on the reducing side, destabilizing it to the point that it is the most labile bond in the ion, resulting in preferential production of even-numbered Bm and Yn ions. This result is consistent with fragmentation of fully charged CS oligosaccharides (all sulfate groups are deprotonated and therefore charged) being a charge-remote process despite the low-energy fragmentation regime. This process is analogous to what is seen when transition metals are coordinated to an oligosaccharide, immobilizing charge and resulting in fragmentation directed by the metal center (Fura & Leary, 1993; Konig & Leary, 1998), where HexNAc fragmentation is suppressed. Care must be taken in setting the collision voltages for tandem mass spectrometric analysis of CS oligosaccharides to avoid overfragmentation of the molecules (Zamfir et al., 2002). The multiply charged ions generated from GAG oligosaccharides are unique among the classes of biomolecules because of the high density of charge per residue. This charge density dramatically changes the CID properties such that bonds are destabilized to approximately the same degree as the molecular mass increases. Thus, the diminishing yield of CID product ions that is observed for peptides is not expected to limit the fragmentation of GAGs to so great a degree. 213 & ZAIA b. Heparin/HS Oligosaccharides. Negative ESI has been used to ionize heparin oligosaccharides for tandem mass spectrometric analysis, and has been found to be useful in quantification (Pope et al., 2001). Tandem mass spectrometric analysis has also been shown to produce abundant glycosidic bond cleavages despite the high degree of sulfation of heparins (Zaia & Costello, 2003). CID production mass spectra generated from the [M–6H]6 ion of an octasulfated pentamer results in abundant ions from glycosidic bond cleavage—as shown in Figure 37 (Zaia & Costello, 2003). The data demonstrate that abundant glycosidic bond cleavages can be obtained, provided that most of the sulfate groups are charged. Repulsions from such charges are likely to destabilize glycosidic bond, and to make their rupture energetically favorable with respect to losses of SO3 from the precursor ion. Glycosidicbond cleavage is also favorable for [M(3Ca)–4H]4 ions generated from the same oligosaccharide, as shown in Figure 38 (Zaia & Costello, 2003). Note that the nomenclature used assumes the protons displaced by the metal cations, and shows only the loss of protons that confer charge on the ion (see Section V.B). More information is produced from the [M(3Ca)–4H]4 CID mass spectrum than from that of the [M–6H]6 ion in the form of glycosidic-bond and cross-ring cleavages. The use of ESI MS for the analysis of highly sulfated oligosaccharides is, therefore, indicated from the point of view of obtaining ions without in-source fragmentation, and of obtaining tandem mass spectra that indicate patterns of bond cleavages that indicate sequence. FIGURE 37. Product-ion tandem mass spectrum of the [M–6H]6 ion generated from an octasulfated synthetic heparin pentamer, using negative ESI QIT MS. Reprinted with permission from Zaia & Costello (2003). Copyright 2003 American Chemical Society. 214 FIGURE 38. Negative-ion ESI tandem mass spectrum of a calcium- complexed synthetic heparin pentamer [M(3Ca)–4H]4, acquired on a Q-oTOF instrument. Reprinted with permission from Zaia & Costello (2003). Copyright 2003 American Chemical Society. D. Conclusions The most important factor for the mass spectrometric analysis of sulfated oligosaccharides is the charge environment of the sulfate groups. Sulfate groups are quite labile when protonated, but become substantially less so when either deprotonated or paired with a metal cation. The decrease in lability is such that losses of SO3 from the precursor ion, either in-source or during CID, become less energetically favorable than cleavage of glycosidic bonds. When ions formed correspond to one negative charge per sulfate group, abundant intact protonated molecule ions result, and abundant glycosidic bond cleavages are observed in CID mass spectra. ESI results in the observation of such charge states, and is, therefore, the choice for analysis of sulfated oligosaccharides. For sulfated N- and O-linked oligosaccharides, the CID product ions all contain the sulfate group, facilitating the identification of the modified residue. Although chondroitin sulfate and keratan sulfate GAGs contain more sulfate groups per monosaccharide unit, the most abundant ions formed with ESI correspond to one charge per sulfate. Because oligosaccharides derived from those GAGs produce abundant glycosidic-bond product ions with losses of SO3 in very low abundances, the product-ion patterns are readily interpreted in terms of the number of sulfate groups on each monosaccharide residue. The abundance of the MASS SPECTROMETRY OF OLIGOSACCHARIDES glycosidic-bond product ions also indicate the position of sulfation (GalNAc 4- or 6-) for CS oligosaccharides. ESI MS is also useful for the analysis of the HLGAGs, a class that contains the highest degree of sulfation of any biomolecule. Provided that gentle desolvation conditions are used in the source, intact deprotonated protonated molecule ions can be observed with no losses of SO3. It is not, however, possible to produce ions in which all sulfate groups are deprotonated, due to charge-charge repulsion. The high density of charge on these ions appears to destabilize glycosidic bonds, and to make their rupture energetically favorable with respect to losses of SO3. Therefore deprotonated HLGAG ions produce both types of fragments that result from CID. Interpretation of those production profiles entails calculating neutral-mass equivalents for each product ion, and identifying pairs that sum to the mass of the neutral intact molecule. Such pairs will be produced without losses of SO3 and will be useful for determining structure. It is necessary to obtain CID MS on all observed charge states, and to combine the useful structural data. An alternative is to use metal cations to stabilize sulfate groups without increasing the ionic charge density. Divalent calcium ions have been shown to be useful for that purpose, producing abundant glycosidic bond as well as some crossring cleavages. That approach seems likely to be the most generally useful because complete or nearly complete fragmentation data are obtained in a single CID spectrum. VII. OVERALL CONCLUSIONS Although the analysis of complex oligosaccharides remains a challenge, recent research has outlined clear approaches with modern mass spectrometry. Mixtures released from a glycoprotein or other biological source may be effectively profiled, provided that steps are taken to make the oligosaccharides chemically equivalent, thereby insuring equal ionization responses. Such steps include chromatographic separation, peralkylation, and methyl esterification. Peralkylated ions produce the most informative tandem mass spectra with regard to the branching structure, by virtue of the unique mass created by bond scission. For analysis of sample quantities below a few micrograms, however, permethylation is not recommended due to sample losses. Many researchers are thus investigating mass spectrometric analysis of native or minimally derivatized samples. Although protonated native ions produce informative product-ion mass spectra, that precursorion form is not recommended because of the possibility of internal-residue rearrangements. At this time, direct interpretation of branched oligosaccharide tandem mass spectra does not seem feasible. Rather, cleavages to labile residues in branced oligosaccharides (Neu5Ac, Fuc and HexNAc) expose core & structures; subsequent stages of fragmentation of those structures provide useful structural fingerprints. Such data will be most useful when compared to a library of tandem mass spectra generated from known oligosaccharide structures. In that way, oligosaccharide substructures released by gas-phase degradation in the mass spectrometer may be identified by a comparision with library spectra. Although the theoretical basis for that approach has been described in a series of publications (Konig & Leary, 1998; Sheeley & Reinhold, 1998; Viseux, de Hoffmann, & Domon, 1998; Weiskopf, Vouros, & Harvey, 1998; Tseng, Hedrick, & Lebrilla, 1999), more development is needed. Specifically, bioinformatics tools must be developed to allow the facile comparison of the tandem mass spectrometric data from unknown and known structures. This process will entail a collaboration among glycoscience research groups to produce the libraries and to conduct detailed studies of the elucidation of unknowns to establish the validity of the gas-phase degradation and library-searching techniques. The public availability of such libraries is likely to result in a dramatic progress in the ability to correlate the expression of specific carbohydrate structures with biological functions. VIII. ABBREVIATIONS CID CS dHex ECD ESI FAB FT Fuc GAG Gal GalNAc Glc GlcA GlcNAc Hex HexA DHexA HexNAc HS HLGAG IRMPD KS MALDI Man Neu5Ac NeuGc collision-induced dissociation chondroitin sulfate deoxyhexose residue electron capture dissociation electrospray ionization fast atom bombardment Fourier transform fucose residue glycosaminoglycan galactose residue N-acetylgalactosamine residue glucose residue glucuronic acid residue N-acetylglucosamine hexose residue hexuronic acid residue 4,5-unsaturated hexuronic acid residue N-acetylhexosamine residue heparan sulfate heparin-like glycosaminoglycan infrared multiphoton dissociation keratan sulfate matrix assisted laser desorption/ionization mannose residue N-acetylneuraminic acid residue N-glycolyl neuraminic acid 215 & ZAIA PSD SDS–PAGE SORI TOF QIT post-source decay sodium dodecyl sulfate–polyacrylamide gel electrophoresis selective off-resonance irradiation time-of-flight quadrupole ion trap REFERENCES Adams J. 1990. Charge-remote fragmentations—Analytical applications and fundamental-studies. Mass Spectrom Rev 9:141–186. Alving K, Paulsen H, Peter-Katalinic J. 1999. Characterization of O-glycosylation sites in MUC2 glycopeptides by nanoelectrospray QTOF mass spectrometry. J Mass Spectrom 34: 395–407. Asam MR, Glish GL. 1997. Tandem mass spectrometry of alkali cationized polysaccharides in a quadrupole ion trap. J Am Soc Mass Spectrom 8:987–995. Bahr U, Pfenninger A, Karas M, Stahl B. 1997. High-sensitivity analysis of neutral underivatized oligosaccharides by nanoelectrospray mass spectrometry. Anal Chem 69:4530–4535. Baldwin MA, Stahl N, Reinders LG, Gibson BW, Prusiner SB, Burlingame AL. 1990. Permethylation and tandem mass spectrometry of oligosaccharides having free hexosamine: Analysis of glycoinisitol phospholipid anchor glycan from the scrapie prion protein. Anal Biochem 191:174–182. Barr JR, Anumula KR, Vettese MB, Taylor PB, Carr SA. 1991. Structural classification of carbohydrates in glycoproteins by mass spectrometry and high-performance anion-exchange chromatography. Anal Biochem 192:181–192. Bateman RH, Green MR, Scott G, Clayton E. 1995. A combined magnetic sector-time-of-flight mass-spectrometer for structural determination studies by tandem mass-spectrometry. Rapid Commun Mass Spectrom 9:1227–1233. Bateman K, White R, Yaguchi M, Thibault P. 1998. Characterization of protein glycoforms by capillary-zone electrophoresis nanoelectrospray mass spectrometry. J Chromatogr A 794:327–344. Beeson JG, Chai W, Rogerson SJ, Lawson AM, Brown GV. 1998. Inhibition of binding of malaria-infected erythrocytes by a tetradecasaccharide fraction from chondroitin sulfate A. Infect Immun 66:3397–3402. Biemann K, Martin SA. 1987. Mass spectrometric determination of the amino acid sequence of peptides and proteins. Mass Spectrom Rev 6:1–76. Bordoli RS, Howes K, Vickers RG, Bateman RH, Harvey DJ. 1994. Matrix-assisted laser-desorption mass-spectrometry on a magnetic-sector. Instrument fitted with an array detector. Rapid Commun Mass Spectrom 8:585–589. Bourne EJ, Stacey M, Tatlow JC, Tedder JM. 1949. Studies on trifluoroacetic acid. Part I. Trifluoroacetic anhydride as a promotor of ester formation between hydroxy-compounds and carboxylic acids. J Chem Soc 2976–2979. 216 Broberg S, Broberg A, Duus JO. 2000. Matrix-assisted laser desorption/ionization time-of-flight mass spectrometry of oligosaccharides derivatized by reductive amination and N,N-dimethylation. Rapid Commun Mass Spectrom 14: 1801–1805. Brüll LP, Heerma W, ThomasOates J, Haverkamp J, Kovacik V, Kovac P. 1997. Loss of internal 1 ! 6 substituted monosaccharide residues from underivatized and per-O-methylated trisaccharides. J Am Soc Mass Spectrom 8:43–49. Brüll LP, Kovacik V, Thomas-Oates JE, Heerma W, Haverkamp J. 1998. Sodium-cationized oligosaccharides do not appear to undergo ‘internal residue loss’ rearrangement processes on tandem mass spectrometry. Rapid Commun Mass Spectrom 12:1520–1532. Burlingame AL, Boyd RK, Gaskell SJ. 1994. Mass spectrometry. Anal Chem 66:634R–683R. Cancilla MT, Penn SG, Lebrilla CB. 1998. Alkaline degradation of oligosaccharides coupled with matrix-assisted laser desorption/ionization Fourier transform mass spectrometry: A method for sequencing oligosaccharides. Analytical Chemistry 70:663–672. Cancilla MT, Penn SG, Carroll JA, Lebrilla CB. 1996. Coordination of alkali metals to oligosaccharides dictates fragmentation behavior in matrix assisted laser desorption ionization Fourier transform mass spectrometry. J Am Chem Soc 118:6736–6745. Cancilla MT, Wong AW, Voss LR, Lebrilla CB. 1999. Fragmentation reactions in the mass spectrometry analysis of neutral oligosaccharides. Anal Chem 71:3206–3218. Carr SA, Huddleston MJ, Bean MF. 1993. Selective identification and differentiation of N- and O-linked oligosaccharides in glycoproteins by liquid chromatography-mass spectrometry. Protein Sci 2:183–196. Carr SA, Reinhold VN. 1984. Structural characterization of sulfated glycosaminoglycans by fast atom bombardment mass spectrometry: Application to chondroitin sulfate. J Carbohydr Chem 3:381–401. Carr SA, Roberts GD. 1986. Carbohydrate mapping by mass spectrometry: A novel method for identifying attachment sites of Asn-linked sugars in glycoproteins. Anal Biochem 157:396–406. Carr SA, Reinhold VN, Green BN, Hass JR. 1985. Enhancement of structural information in FAB ionized carbohydrate samples by neutral gas collision. Biomed Mass Spectrom 12:288–295. Carr SA, Hemling ME, Folena-Wasserman G, Sweet RW, Anumula K, Barr JR, Huddleston MJ, Taylor P. 1989. Protein and carbohydrate structural analysis of a recombinant soluble CD4 receptor by mass spectrometry. J Biol Chem 264:21286–21295. Carr SA, Barr JR, Roberts GD, Anumula KR, Taylor PB. 1990. Identification of attachment sites and structural classes of asparagine- linked carbohydrates in glycoproteins. Methods Enzymol 193:501–518. Chai W, Beeson JG, Lawson AM. 2002. The structural motif in chondroitin sulfate for adhesion of Plasmodium falciparum- MASS SPECTROMETRY OF OLIGOSACCHARIDES infected erythrocytes comprises disaccharide units of 4-Osulfated and non-sulfated N-acetylgalactosamine linked to glucuronic acid. J Biol Chem 277:22438–22446. Chai W, Kogelberg H, Lawson AM. 1996. Generation and structural characterization of a range of unmodified chondroitin sulfate oligosaccharide fragments. Anal Biochem 237:88–102. Chai W, Rosankiewicz JR, Lawson AM. 1995. TLC-LSIMS of neoglycolipids of glycosaminoglycan disaccharides and of oxymercuration cleavage products of heparin fragments that contain unsaturated uronic acid. Carbohydr Res 269:111– 124. Chai W, Luo J, Lim CK, Lawson AM. 1998. Characterization of heparin oligosaccharide mixtures as ammonium salts using electrospray mass spectrometry. Anal Chem 70:2060– 2066. Chance DL, Mawhinney TP. 1996. Disulfated oligosaccharides derived from tracheobronchial mucous glycoproteins of a patient suffering from cystic fibrosis. Carbohydr Res 295: 157–177. Charlwood J, Tolson D, Dwek M, Camilleri P. 1999. A detailed analysis of neutral and acidic carbohydrates in human milk. Anal Biochem 273:261–277. Chen P, Werner-Zwanziger U, Wiesler D, Pagel M, Novotny MV. 1999. Mass spectrometric analysis of benzoylated sialooligosaccharides and differentiation of terminal alpha 2 ! 3 and alpha 2 ! 6 sialogalactosylated linkages at subpicomole levels. Anal Chem 71:4969–4973. Ciucanu I, Kerek F. 1984. A simple and rapid method for the permethylation of carbohydrates. Carbohydr Res 131:209– 217. Claeys M, VandenHeuvel H, Chen S, Derrick PJ, Mellon FA, Price KR. 1996. Comparison of high- and low-energy collision-induced. Dissociation tandem mass spectrometry in the analysis of glycoalkaloids and their aglycons. J Am Soc Mass Spectrom 7:173–181. Colangelo J, Licon V, Benen J, Visser J, Bergmann C, Orlando R. 1999. Characterization of the N-linked glycosylation site of recombinant pectate lyase. Rapid Commun Mass Spectrom 13:2382–2387. Cole RB. 1997. Electrospray ionization mass spectrometry: Fundamentals, Instrumentation and Applications. New York, NY: John Wiley, Inc. Conboy JJ, Henion JD. 1992. The determination of glycopeptides by liquid-chromatography mass-spectrometry with collision-induced dissociation. J Am Soc Mass Spectrom 3:804– 814. Costello CE, Juhasz P, Perreault H. 1994. New mass spectral approaches to ganglioside structure determinations. Prog Brain Res 101:45–61. Costello CE, Vath JE. 1990. Tandem mass spectrometry of glycolipids. Methods Enzymol 193:738–768. Dage JL, Ackermann BL, Halsall HB. 1998. Site localization of sialyl Lewis(x) antigen on alpha1-acid glycoprotein by high performance liquid chromatography-electrospray mass spectrometry. Glycobiology 8:755–760. & Daniel PF. 1987. Separation of benzoylated oligosaccharides by reversed-phase high-pressure liquid chromatography: Application to high-mannose type oligosaccharides. Methods Enzymol 138:94–116. Delaney J, Vouros P. 2001. Liquid chromatography ion trap mass spectrometric analysis of oligosaccharides using permethylated derivatives. Rapid Commun Mass Spectrom 15:325– 334. Dell A. 1990. Preparation and desorption mass spectrometry of permethyl and peracetyl derivatives of oligosaccharides. Methods Enzymol 193:647–660. Dell A, Morris HR, Egge H, von Nicolai H, Strecker G. 1983a. Fast-atom bombardment mass spectrometry for carbohydrate structure determination. Carbohydr Res 115:41–52. Dell A, Oates JE, Morris HR, Egge H. 1983b. Structure determination of carbohydrates and glycosphingolipids by fast atom bombardment mass spectrometry. Int J Mass Spectrom Ion Phys 46:415–418. Dell A, Rogers M, Thomas-Oates J, Huckerby T, Sanderson P, Nieduszynski I. 1988. Fast-atom-bombardment mass-spectrometric strategies for sequencing sulfated oligosaccharides. Carbohydr Res 179:7–19. Dell A, Reason AJ, Khoo KH, Panico M, McDowell RA, Morris HR. 1994. Mass spectrometry of carbohydrate-containing biopolymers. Methods Enzymol 230:108–132. Derappe C, Bauvy C, Lemonnier M, Lhermitte M, Platzer N, Egge H, Peter-Katalinc J, van Halbeek H, Kamerling JP, Vliegenthart JF. 1986. Determination of the structure of a fucose-containing trisaccharide monophosphate isolated from human pregnancy urine. Carbohydr Res 150:273– 284. Desaire H, Sirich TL, Leary JA. 2001. Evidence of block and randomly sequenced chondroitin polysaccharides: sequential enzymatic digestion and quantification using ion trap tandem mass spectrometry. Anal Chem 73:3513–3520. Desaire H, Leary JA. 1999a. Differentiation of diastereomeric Nacetylhexosamine monosaccharides using ion trap tandem mass spectrometry. Anal Chem 71:1997–2002. Desaire H, Leary JA. 1999b. Multicomponent quantification of diastereomeric hexosamine monosaccharides using ion trap tandem mass spectrometry. Anal Chem 71:4142–4147. Desaire H, Leary J. 2000. Detection and quantification of the sulfated disaccharides in chondoritin sulfate by electrospray tandem mass spectrometry. J Am Soc Mass Spectrom 11:916–920. Domon B, Müller DR, Richter WJ. 1989. Identification of interglycosidic linkages and sugar constituents of larger glycosides by tandem mass spectrometry. Organic Mass Spectrom 24:357–359. Domon B, Müller DR, Richter WJ. 1990. High performance tandem mass spectrometry for sequence, branching and interglycosidic linkage analysis of peracetylated oligosaccharides. Biomed Environ Mass Spectrom 19:390–392. Domon B, Costello CE. 1988a. Structure elucidation of glycosphingolipids and gangliosides using high-performance tandem mass spectrometry. Biochemistry 27:1534–1543. 217 & ZAIA Domon B, Costello CE. 1988b. A systematic nomenclature for carbohydrate fragmentations in FAB-MS/MS spectra of glycoconjugates. Glycoconjugate J 5:397–409. Duffin KL, Welply JK, Huang E, Henion JD. 1992. Characterization of N-linked oligosaccharides by electrospray and tandem mass spectrometry. Anal Chem 64:1440–1448. Dufresne CP, Wood TD, Hendrickson CL. 1998. High-resolution electrospray ionization Fourier transform mass spectrometry with infrared multiphoton dissociation of glucokinase from bacillus stearothermophilus. J Am Soc Mass Spectrom 9:1222–1225. Duteil S, Gareil P, Girault S, Mallet A, Feve C, Siret L. 1999. Identification of heparin oligosaccharides by direct coupling of capillary electrophoresis/ionspray-mass spectrometry. Rapid Commun Mass Spectrom 13:1889–1898. Egge H, Dell A, Von Nicolai H. 1983. Fucose containing oligosaccharides from human milk. I. Separation and identification of new constituents. Arch Biochem Biophys 224: 235–253. Egge H, Peter-Katalinc J. 1987. Fast atom bombardment mass spectrometry for structural elucidation of glycoconjugates. Mass Spectrom Rev 6:331–393. Ernst B, Müller DR, Richter WJ. 1997. False sugar sequence ions in electrospray tandem mass spectrometry of underivatized sialyl-Lewis-type oligosaccharides. Int J Mass Spectrom Ion Proc 160:283–290. Ernst S, Rhomberg AJ, Biemann K, Sasisekharan R. 1998. Direct evidence for a predominantly exolytic processive mechanism for depolymerization of heparin-like glycosaminoglycans by heparinase I. Proc Natl Acad Sci USA 95:4182– 4187. Ethier M, Saba JA, Ens W, Standing KG, Perreault H. 2002. Automated structural assignment of derivatized complex Nlinked oligosaccharides from tandem mass spectra. Rapid Commun Mass Spectrom 16:1743–1754. Fenn JB, Mann M, Meng CK, Wong SF. 1990. Electrospray ionization—Principles and practice. Mass Spectrom Rev 9:37–70. Flora JW, Hannis JC, Muddiman DC. 2001. High-mass accuracy of product ions produced by SORI-CID using a dual electrospray ionization source coupled with FTICR mass spectrometry. Anal Chem 73:1247–1251. Fournet B, Dhalluin JM, Strecker G, Montreuil J, Bosso C, Defaye J. 1980. Gas-liquid chromatography and mass spectrometry of oligosaccharides obtained by partial acetolysis of glycans of glycoproteins. Anal Biochem 108:35–56. Fura A, Leary JA. 1993. Differentiation of Ca2þ-coordinated and Mg2þ-coordinated branched trisaccharide isomers—An electrospray-ionization and tandem mass-spectrometry study. Anal Chem 65:2805–2811. Gallego RG, Blanco JLJ, Thijssen-van Zuylen C, Gotfredsen CH, Voshol H, Duus JO, Schachner M, Vliegenthart JFG. 2001. Epitope diversity of N-glycans from bovine peripheral myelin glycoprotein P0 revealed by mass spectrometry and nano probe magic angle spinning H-1 NMR spectroscopy. J Biol Chem 276:30834–30844. 218 Gangl ET, Annan MM, Spooner N, Vouros P. 2001. Reduction of signal suppression effects in ESI-MS using a nanosplitting device. Anal Chem 73:5635–5644. Garozzo D, Giuffrida M, Impallomeni G, Ballistreri A, Montaudo G. 1990. Determination of linkage position and identification of the reducing end in linear oligosaccharides by negative-ion fast atom bombardment mass-spectrometry. Anal Chem 62:279–286. Gaucher SP, Morrow J, Leary JA. 2000. STAT: A saccharide topology analysis tool used in combination with tandem mass spectrometry. Anal Chem 72:2331–2336. Gaucher SP, Pedersen SF, Leary JA. 1999. Stereospecific synthesis and characterization of aminoglycoside ligands from diethylenetriamine. J Org Chem 64:4012–4015. Gaucher SP, Leary JA. 1998. Stereochemical differentiation of mannose, glucose, galactose, and talose using zinc(II) diethylenetriamine and ESI-ion trap mass spectrometry. Anal Chem 70:3009–3014. Gaucher SP, Cancilla MT, Phillips NJ, Gibson BW, Leary JA. 2000. Mass spectral characterization of lipooligosaccharides from Haemophilus influenzae 2019. Biochemistry 39: 12406–12414. Gauthier JW, Trautman TR, Jacobson DB. 1991. Sustained offresonance irradiation for collision-activated dissociation involving Fourier transform mass spectrometry. Collisionactivated dissosiation technique that emulates infrared multiphoton dissociation. Analytica Chimica Acta 246: 211–225. Geyer R, Geyer H. 1994. Saccharide linkage analysis using methylation and other techniques. Methods Enzymol 230: 86–108. Geyer H, Bahr U, Liedtke S, Schachner M, Geyer R. 2001. Core structures of polysialylated glycans present in neural cell adhesion molecule from newborn mouse brain. Eur J Biochem 268:6587–6599. Gibson BW, Cohen P. 1990. Liquid secondary ion mass spectrometry of phosphorylated and sulfated peptides and proteins. Methods Enzymol 193:480–501. Gillece-Castro BL, Burlingame AL. 1990. Oligosaccharide characterization with high-energy collision-induced dissociation mass spectrometry. Methods Enzymol 193:689– 712. Guan S, Marshall AG, Wahl MC. 1994. MS/MS with high detection efficiency and mass resolving power for product ions in Fourier transform ion cyclotron resonance mass spectrometry. Anal Chem 66:1363–1367. Guerrini M, Agulles T, Bisio A, Hricovini M, Lay L, Naggi A, Poletti L, Sturiale L, Torri G, Casu B. 2002. Minimal heparin/heparan sulfate sequences for binding to fibroblast growth factor-1. Biochem Biophys Res Commun 292:222– 230. Gunay NS, Tadano-Aritomi K, Toida T, Ishizuka I, Linhardt R. 2003. Evaluation of counterions for electrospray ionization mass spectral analysis of a highly sulfated carbohydrate, sucrose octasulfate. Anal Chem 75:3226– 3231. MASS SPECTROMETRY OF OLIGOSACCHARIDES Gunnarson A. 1987. N- and O-alkylation of glycoconjugates and polysaccharides by solid base dimethyl sulphoxide/alkyl iodide. Glycoconj J 4:239–245. Hakomori S-I. 1964. Rapid permethylation of glycolipid, and polysaccharide catalyzed by methylsulfinyl carbanion in dimethyl sulfoxide. J Biochem 55:205–208. Hallgren P, Lundblad A. 1977. Structural analysis of nine oligosaccharides isolated from the urine of a blood group O, nonsecretor, woman during pregnancy and lactation. J Biol Chem 252:1014–1022. Handa S, Nakamura K. 1984. Modification of sialic acid carboxyl group of ganglioside. J Biochem (Tokyo) 95:1323–1329. Hanisch FG, Jovanovic M, Peter-Katalinic J. 2001. Glycoprotein identification and localization of O-glycosylation sites by mass spectrometric analysis of deglycosylated/alkylaminylated peptide fragments. Anal Biochem 290:47–59. Hanrahan S, Charlwood J, Tyldesley R, Langridge J, Bordoli R, Bateman R, Camilleri P. 2001. Facile sequencing of oligosaccharides by matrix-assisted laser desorption/ionisation on a hybrid quadrupole orthogonal acceleration time-offlight mass spectrometer. Rapid Commun Mass Spectrom 15:1141–1151. Harvey DJ. 1993. Quantitative aspects of the matrix-assisted laser-desorption mass-spectrometry of complex oligosaccharides. Rapid Commun Mass Spectrom 7:614–619. Harvey DJ. 2000a. Collision-induced fragmentation of underivatized N-linked carbohydrates ionized by electrospray. J Mass Spectrom 35:1178–1190. Harvey DJ. 2000b. Electrospray mass spectrometry and fragmentation of N-linked carbohydrates derivatized at the reducing terminus. J Am Soc Mass Spectrom 11:900–915. Harvey DJ. 2000c. N-(2-diethylamino)ethyl-4-aminobenzamide derivative for high sensitivity mass spectrometric detection and structure determination of N-linked carbohydrates. Rapid Commun Mass Spectrom 14:862–871. Harvey DJ. 2001. Ionization and collision-induced fragmentation of N-linked and related carbohydrates using divalent cations. J Am Soc Mass Spectrom 12:926–937. Harvey DJ, Bateman RH, Green MR. 1997. High-energy collision-induced fragmentation of complex oligosaccharides ionized by matrix-assisted laser desorption/ionization mass spectrometry. J Mass Spectrom 32:167–187. Harvey DJ, Naven TJP, Küster B, Bateman RH, Green MR, Critchley G. 1995. Comparison of fragmentation modes for the structural determination of complex oligosaccharides ionized by matrix-assisted laser. Rapid Commun Mass Spectrom 9:1556–1561. Harvey DJ, Bateman RH, Bordoli RS, Tyldesley R. 2000. Ionisation and fragmentation of complex glycans with a quadrupole time-of-flight mass spectrometer fitted with a matrix-assisted laser desorption/ionisation ion source. Rapid Commun Mass Spectrom 14:2135–2142. Harvey DJ, Mattu TS, Wormald MR, Royle L, Dwek RA, Rudd PM. 2002. ‘‘Internal residue loss’’: Rearrangements occurring during the fragmentation of carbohydrates derivatized at the reducing terminus. Anal Chem 74:734–740. & Heck AJR, Derrick PJ. 1997. Ultrahigh mass accuracy in isotopeselective collision-induced dissociation using correlated sweep excitation and sustained off-resonance irradiation— A Fourier transform ion cyclotron resonance mass spectrometry case study on the [M þ 2H](2þ) ion of Bradykinin. Anal Chem 69:3603–3607. Hettich RL, Stemmler EA. 1996. Investigation of oligonucleotide fragmentation with matrix-assisted laser desorption/ionization Fourier-transform mass spectrometry and sustained offresonance irradiation. Rapid Commun Mass Spectrom 10: 321–327. Hillenkamp F, Karas M, Beavis RC, Chait BT. 1991. Matrixassisted laser desorption/ionization mass spectrometry of biopolymers. Anal Chem 63:1193A–1203A. Hirayama K, Yuji R, Yamada N, Kato K, Arata Y, Shimada I. 1998. Complete and rapid peptide and glycopeptide mapping of mouse monoclonal antibody by LC/MS/MS using ion trap mass spectrometry. Anal Chem 70:2718–2725. Hofmeister GE, Zhou Z, Leary JA. 1991. Linkage position determination in lithium-cationized disaccharides—Tandem mass-spectrometry and semiempirical calculations. J Am Chem Soc 113:5964–5970. Hofstadler SA, Wahl JH, Bakhtiar R, Anderson GA, Bruce JE, Smith RD. 1994. Capillary electrophoresis Fourier transform ion cyclotron resonance mass spectrometry with sustained off-resonance irradiation for the characterization of protein and peptide mixtures. J Am Soc Mass Spectrom 5:894–899. Honda S, Akao E, Suzuki S, Okuda M, Kakehi K, Nakamura J. 1989. High-performance liquid-chromatography of reducing carbohydrates as strongly ultraviolet-absorbing and electrochemically sensitive 1-phenyl-3-methyl-5-pyrazolone derivatives. Anal Biochem 180:351–357. Hsu FF, Bohrer A, Turk J. 1998. Electrospray ionization tandem mass spectrometric analysis of sulfatide. Determination of fragmentation patterns and characterization of molecular species expressed in brain and in pancreatic islets. Biochim Biophys Acta 1392:202–216. Huang Y, Pasa-Tolic L, Guan S, Marshall AG. 1994. Collisioninduced dissociation for mass spectrometric analysis of biopolymers: High-resolution Fourier transform ion cyclotron resonance MS4. Anal Chem 66:4385–4389. Huberty MC, Vath JE, Yu W, Martin SA. 1993. Site-specific carbohydrate identification in recombinant proteins using MALD-TOF MS. Anal Chem 65:2791–2800. Huddleston MJ, Bean MF, Carr SA. 1993. Collisional fragmentation of glycopeptides by electrospray ionization LC/MS and LC/MS/MS: Methods for selective detection of glycopeptides in protein digests. Anal Chem 65:877–884. Hui JP, Lanthier P, White TC, McHugh SG, Yaguchi M, Roy R, Thibault P. 2001. Characterization of cellobiohydrolase I (Cel7A) glycoforms from extracts of Trichoderma reesei using capillary isoelectric focusing and electrospray mass spectrometry. J Chromatogr B Biomed Sci Appl 752:349– 368. Håkansson K, Cooper HJ, Emmett MR, Costello CE, Marshall AG, Nilsson CL. 2001. Electron capture dissociation and 219 & ZAIA Infrared multiphoton dissociation MS/MS of an N-glycosylated tryptic peptide to yield complementary sequence information. Anal Chem 73:4530–4536. Ii T, Okuda S, Hirano T, Ohashi M. 1994. Tandem mass spectrometry for characterization of unsaturated disaccharides from chondroitin sulfate, dermatan sulfate and hyaluronan. Glycoconj J 11:123–132. Ii T, Ohashi Y, Nunomura S, Ogawa T, Nagai Y. 1995. Fast atom bombardment and electrospray ionization tandem mass spectrometry of sulfated Lewis(x) trisaccharides. J Biochem (Tokyo) 118:526–533. Ii T, Kubota M, Hirano T, Ohashi M, Yoshida K, Suzuki S. 1995a. FAB CID-MS/MS characterization of tetrasaccharide triand tetrasulfate derived from the antigenic determinant recognized by the anti-chondroitin sulfate monoclonal antibody MO-225. Glycoconj J 12:282–289. Ii T, Kubota M, Okuda S, Hirano T, Ohashi M. 1995b. Negativeion fast atom bombardment tandem mass spectrometry for characterization of sulfated unsaturated disaccharides from heparin and heparan sulfate. Glycoconj J 12:162– 172. Itoh S, Kawasaki N, Ohta M, Hayakawa T. 2002. Structural analysis of a glycoprotein by liquid chromatography-mass spectrometry and liquid chromatography with tandem mass spectrometry—Application to recombinant human thrombomodulin. J Chromatogr A 978:141–152. Jacobs A, Dahlman O. 2001. Enhancement of the quality of MALDI mass spectra of highly acidic oligosaccharides by using a nafion-coated probe. Anal Chem 73:405–410. Jacoby ES, Kicman AT, Laidler P, Iles RK. 2000. Determination of the glycoforms of human chorionic gonadotropin betacore fragment by matrix-assisted laser desorption/ionization time-of-flight mass spectrometry. Clin Chem 46:1796– 1803. Johnson JV, Yost RA. 1985. Tandem mass spectrometry for trace analysis. Anal Chem 57:758A–768A. Johnson JV, Yost RA, Kelley PE, Bradford DC. 1990. Tandem-inspace and tandem-in-time mass spectrometry: Triple quadrupoles and quadrupole ion traps. Anal Chem 62:2162– 2172. Juhasz P, Biemann K. 1995. Utility of non-covalent complexes in the matrix-assisted laser desorption ionization mass spectrometry of heparin-derived oligosaccharides. Carbohydr Res 270:131–147. Kakehi K, Kinoshita M, Kawakami D, Tanaka J, Sei K, Endo K, Oda Y, Iwaki M, Masuko T. 2001. Capillary electrophoresis of sialic acid-containing glycoprotein. Effect of the heterogeneity of carbohydrate chains on glycoform separation using an alpha(1)-acid glycoprotein as a model. Anal Chem 73:2640–2647. Karas M, Bahr U, Dulcks T. 2000. Nano-electrospray ionization mass spectrometry: Addressing analytical problems beyond routine. Fresenius J Anal Chem 366:669–676. Karlsson H, Carlstedt I, Hansson GC. 1989. The use of gas chromatography and gas chromatography-mass spectrometry for the characterization of permethylated oligosac- 220 charides with molecular mass up to 2300. Anal Biochem 182:438–446. Karlsson NG, Karlsson H, Hansson GC. 1995. Strategy for the investigation of O-linked oligosaccharides from mucins based on the separation into neutral, sialic acid- and sulfatecontaining species. Glycoconj J 12:69–76. Karlsson NG, Karlsson H, Hansson GC. 1996. Sulphated mucin oligosaccharides from porcine small intestine analysed by four-sector tandem mass spectrometry. J Mass Spectrom 31:560–572. Kawasaki N, Haishima Y, Ohta M, Itoh S, Hyuga M, Hyuga S, Hayakawa T. 2001. Structural analysis of sulfated N-linked oligosaccharides in erythropoietin. Glycobiology 11:1043– 1049. Kelleher NL, Taylor SV, Grannis D, Kinsland C, Chiu HJ, Begley TP, McLafferty FW. 1998. Efficient sequence analysis of the six gene products (7–74 kDa) from the Escherichia coli thiamin biosynthetic operon by tandem high-resolution mass spectrometry. Protein Sci 7:1796– 1801. Kelleher NL, Zubarev RA, Bush K, Furie B, Furie BC, McLafferty FW, Walsh CT. 1999. Localization of labile posttranslational modifications by electron capture dissociation: The case of gamma-carboxyglutamic acid. Anal Chem 71:4250–4253. Khoo KH, Morris HR, McDowell RA, Dell A, Maccarana M, Lindahl U. 1993. FABMS/derivatisation strategies for the analysis of heparin-derived oligosaccharides. Carbohydr Res 244:205–223. Kim YS, Ahn MY, Wu SJ, Kim DH, Toida T, Teesch LM, Park Y, Yu G, Lin J, Linhardt RJ. 1998. Determination of the structure of oligosaccharides prepared from acharan sulfate. Glycobiology 8:869–877. Kinoshita A, Yamada S, Haslam SM, Morris HR, Dell A, Sugahara K. 1997. Novel tetrasaccharides isolated from squid cartilage chondroitin sulfate E contain unusual sulfated disaccharide units GlcA(3-O-sulfate)beta1-3GalNAc(6-O-sulfate) or GlcA(3-O-sulfate)beta1-3GalNAc. J Biol Chem 272:19656–19665. Kinoshita A, Yamada S, Haslam SM, Morris HR, Dell A, Sugahara K. 2001. Isolation and structural determination of novel sulfated hexasaccharides from squid cartilage chondroitin sulfate E that exhibits neuroregulatory activities. Biochemistry 40:12654–12665. Kjeldsen F, Haselmann KF, Budnik BA, Sorensen ES, Zubarev RA. 2003. Complete characterization of posttranslational modification sites in the bovine milk protein PP3 by tandem mass spectrometry with electron capture dissociation as the last stage. Anal Chem 75:2355–2361. Kohler M, Leary JA. 1995. LC/MS/MS of carbohydrates with postcolumn addition of metal chlorides using a triaxial electrospray probe. Anal Chem 67:3501–3508. Konig S, Leary JA. 1998. Evidence for linkage position determination in cobalt coordinated pentasaccharides using ion trap mass spectrometry. J Am Soc Mass Spectrom 9:1125– 1134. MASS SPECTROMETRY OF OLIGOSACCHARIDES Kovacik V, Hirsch J, Kovac P, Heerma W, Thomasoates J, Haverkamp J. 1995. Oligosaccharide characterization using collision-induced dissociation fast-atom-bombardment mass-spectrometry—Evidence for internal monosaccharide residue loss. J Mass Spectrom 30:949–958. Kragten EA, Bergwerff AA, vanOostrum J, Müller DR, Richter WJ. 1995. Site-specific analysis of the N-glycans on murine polymeric immunoglobulin a using liquid chromatography electrospray mass spectrometry. J Mass Spectrom 30:1679– 1686. Kruger NA, Zubarev RA, Horn DM, McLafferty FW. 1999. Electron capture dissociation of multiply charged peptide cations. Int J Mass Spectrom 185/186/187:787–793. Krutchinsky AN, Loboda AV, Spicer VL, Dworschak R, Ens W, Standing KG. 1998. Orthogonal injection of matrix-assisted laser desorption/ionization ions into a time-of-flight spectrometer through a collisional damping interface. Rapid Commun Mass Spectrom 12:508–518. Kuberan B, Lech M, Zhang L, Wu ZL, Beeler DL, Rosenberg R. 2002. Analysis of heparan sulfate oligosaccharides with ion pair-reverse phase capillary high performance liquid chromatography—Microelectrospray ionization time-offlight mass Spectrometry. J Am Chem Soc 124:8707– 8718. Kushi Y, Handa S, Ishizuka I. 1985. Secondary ion mass spectrometry for sulfoglycolipids: application of negative ion detection. J Biochem (Tokyo) 97:419–428. Kwan CP, Venkataraman G, Shriver Z, Raman R, Liu D, Qi Y, Varticovski L, Sasisekharan R. 2001. Probing fibroblast growth factor dimerization and role of heparin-like glycosaminoglycans in modulating dimerization and signaling. J Biol Chem 276:23421–23429. Körner R, Limberg G, Mikkelsen JD, Roepstorff P. 1998. Characterization of enzymatic pectin digests by matrixassisted laser desorption/ionization mass spectrometry. J Mass Spectrom 33:836–842. Körner R, Limberg G, Christensen TM, Mikkelsen JD, Roepstorff P. 1999. Sequencing of partially methylesterified oligogalacturonates by tandem mass spectrometry and its use to determine pectinase specificities. Anal Chem 71:1421–1427. Küster B, Mann M. 1999. 18O-labeling of N-glycosylation sites to improve the identification of gel-separated glycoproteins using peptide mass mapping and database searching. Anal Chem 71:1431–1440. Küster B, Wheeler SF, Hunter AP, Dwek RA, Harvey DJ. 1997. Sequencing of N-linked oligosaccharides directly from protein gels: In-gel deglycosylation followed by matrixassisted laser desorption/ionization mass spectrometry and normal-phase high-performance liquid chromatography. Anal Biochem 250:82–101. Küster B, Hunter AP, Wheeler SF, Dwek RA, Harvey DJ. 1998. Structural determination of N-linked carbohydrates by matrix-assisted laser desorption/ionization-mass spectrometry following enzymatic release within sodium dodecyl sulphate-polyacrylamide electrophoresis gels: Application & to species-specific glycosylation of alpha1-acid glycoprotein. Electrophoresis 19:1950–1959. Laiko VV, Baldwin MA, Burlingame AL. 2000. Atmospheric pressure matrix assisted laser desorption/ionization mass spectrometry. Anal Chem 72:652–657. Laiko VV, Moyer SC, Cotter RJ. 2000. Atmospheric pressure MALDI/ion trap mass spectrometry. Anal Chem 72:5239– 5243. Laine RA. 1989. Tandem mass spectrometry of oligosaccharides. Methods Enzymol 179:157–164. Laine RA, Pamidimukkala KM, French AD, Hall RW, Abbas SA, Jain RK, Matta KL. 1988. Linkage position in oligosaccharides by fast atom bombardment ionization, collisionactivated dissociation, tandem mass-spectrometry and molecular modeling—L-Fucosylp-(alpha-1-]3)-D-N-acetylD -glucosamylp-(beta-1-]3)- D -galactosylp-(beta-1-OMethyl), L-fucosylp-(alpha-1-]4)-D-N-acetyl-D-glucosaminylp-(beta-1-]3)-D-galactosylp-(beta-1-O-methyl), L-fucosylp-(alpha-1-]6)-D-N-acetyl-D-glucosaminylp-(beta-1-]3)D-galactosylp-(beta-1-O-methyl)-. J Am Chem Soc 110: 6931–6939. Laine RA, Yoon E, Mahier TJ, Abbas SA, de Lappe B, Jain R, Matta K. 1991. Non-reducing terminal linkage position determination in intact and permethylated synthetic oligosaccharides having a penultimate amino sugar: Fast atom bombardment ionization, collisional-induced dissociation and tandem mass spectrometry. Biol Mass Spectrom 20: 505–514. Lamb DJ, Wang HM, Mallis LM, Linhardt RJ. 1992. Negative ion fast-atom bombardment tandem mass spectrometry to determine sulfate and linkage position in glycosaminoglycan derived disaccharides. J Am Soc Mass Spectrom 3:797– 803. Leavell MD, Leary JA. 2001. Stabilization and linkage analysis of metal-ligated sialic acid containing oligosaccharides. J Am Soc Mass Spectrom 12:528–536. Lemoine J, Chirat F, Domon B. 1996. Structural analysis of derivatized oligosaccharides using post-source decay matrixassisted laser desorption ionization mass spectrometry. J Mass Spectrom 31:908–912. Lemoine J, Strecker G, Leroy Y, Fournet B, Ricart G. 1991. Collisional-activation tandem mass spectrometry of sodium adduct ions of methylated oligosaccharides: sequence analysis and discrimination between alpha-NeuAc-(2–3) and alpha-NeuAc-(2–6) linkages. Carbohydr Res 221:209– 217. Lemoine J, Fournet B, Despeyroux D, Jennings KR, Rosenberg R, Dehoffmann E. 1993. Collision-induced dissociation of alkali-metal cationized and permethylated oligosaccharides—Influence of the collision energy and of the collision gas for the assignment of linkage position. J Am Soc Mass Spectrom 4:197–203. Li T, Kubota M, Okuda S, Hirano T, Ohashi M. 1995. Positive-ion fast atom bombardment tandem mass spectrometry for characterization of sulfated disaccharides from heparin and heparansulfate. Eur Mass Spectrom 1:11–22. 221 & ZAIA Lindberg B, Lonngren J. 1978. Methylation analysis of complex carbohydrates: general procedure and application for sequence analysis. Methods Enzymol 50:3–33. Linhardt RJ, Wang HM, Loganthan D, Lamb DJ, Mallis LM. 1992. Analysis of glycosaminoglycan-derived oligosaccharides using fast atom bombardment mass spectrometry. Carbohydr Res 225:137–145. Little DP, McLafferty FW. 1995. Sequencing 50-Mer DNAs using electrospray tandem mass spectrometry and complementary fragmentation methods. J Am Chem Soc 117: 6783–6784. Little DP, Speir JP, Senko MW, Oconnor PB, McLafferty FW. 1994. Infrared multiphoton dissociation of large multiply charged ions for biomolecule sequencing. Anal Chem 66:2809–2815. Little DP, Aaserud DJ, Valaskovic GA, McLafferty FW. 1996. Sequence information from 42–108-Mer DNAs (complete for a 50-Mer) by tandem mass spectrometry. J Am Chem Soc 118:9352–9359. Liu J, Shriver Z, Pope M, Thorp SC, Duncan MB, Copeland RJ, Raska CS, Yoshida K, Eisenberg RJ, Cohen G, Linhardt RJ, Sasisekharan R. 2002. Characterization of a heparan sulfate octasaccharide that binds to herpes simplex viral type 1 glycoprotein D. J Biol Chem 277:33456–33467. Loboda AV, Krutchinsky AN, Bromirski M, Ens W, Standing KG. 2000. A tandem quadrupole/time-of-flight mass spectrometer with a matrix-assisted laser desorption/ionization source: Design and performance. Rapid Commun Mass Spectrom 14:1047–1057. LoGuidice JM, Herz H, Lamblin G, Plancke Y, Roussel P, Lhermitte M. 1997. Structures of sulfated oligosaccharides isolated from the respiratory mucins of a non-secretor (O, Le(aþb–)) patient suffering from chronic bronchitis. Glycoconj J 14:113–125. Macek B, Hofsteenge J, Peter-Katalinic J. 2001. Direct determination of glycosylation sites in O-fucosylated glycopeptides using nano-electrospray quadrupole timeof-flight mass spectrometry. Rapid Commun Mass Spectrom 15:771–777. Maier CS, Yan XG, Harder ME, Schimerlik MI, Deinzer ML, Pasa-Tolic L, Smith RD. 2000. Electrospray ionization Fourier transform ion cyclotron resonance mass spectrometric analysis of the recombinant human macrophage colony stimulating factor beta and derivatives. J Am Soc Mass Spectrom 11:237–243. Mallis LM, Wang HM, Loganathan D, Linhardt RJ. 1989. Sequence analysis of highly sulfated, heparin-derived oligosaccharides using fast atom bombardment mass spectrometry. Anal Chem 61:1453–1458. Mann M, Wilm M. 1994. Error-tolerant identification of peptides in sequence databases by peptide sequence tags. Anal Chem 66:4390–4399. Marbois BN, Faull KF, Fluharty AL, Raval-Fernandes S, Rome LH. 2000. Analysis of sulfatide from rat cerebellum and multiple sclerosis white matter by negative ion electrospray mass spectrometry. Biochim Biophys Acta 1484:59–70. 222 March RE. 1998. Quadrupole ion trap mass spectrometry: Theory, simulation, recent developments and applications. Rapid Commun Mass Spectrom 12:1543–1554. March RE. 2000. Quadrupole ion trap mass spectrometry: Aview at the turn of the century. Int J Mass Spectrom 200:285–312. March RE, Hughes HE. 1989. Quadrupole storage mass spectrometry. New York, NY: John Wiley & Sons, Inc. March RE, Londry FA. 1995. Theory of quadrupole mass spectrometry. In: March RE, Todd JFJ, editors. Practical aspects of ion trap mass spectrometry. Vol. 1. New York, NY: CRC Press, Inc. pp 25–48. Marshall AG, Wang T-CL, Ricca TL. 1985. Tailored excitation for Fourier transform ion cyclotron resonance mass spectrometry. J Am Chem Soc 107:7893–7897. Mawhinney TP, Chance DL. 1994. Structural elucidation by fastatom-bombardment mass-spectrometry of multisulfated oligosaccharides isolated from human respiratory mucous glycoproteins. J Carbohydr Chem 13:825–840. McClellan JM, Costello CE, O’Connor PB, Zaia J. 2002. Influence of charge state on product ion mass spectra and the determination of 4S/6S sulfation sequence of chondroitin sulfate oligosaccharides. Anal Chem 74:3760–3771. McNeil M, Darvill AG, Aman P, Franzen LE, Albersheim P. 1982. Structural analysis of complex carbohydrates using high-performance liquid chromatography, gas chromatography, and mass spectrometry. Methods Enzymol 83:3–45. Medzihradszky KF, Gillece-Castro BL, Settineri CA, Townsend RR, Masiarz FR, Burlingame AL. 1990. Structure determination of O-linked glycopeptides by tandem mass spectrometry. Biomed Environ Mass Spectrom 19:777–781. Medzihradszky K, Gillece-Castro B, Townsend R, Burlingame A, Hardy M. 1996. Structural elucidation of O-linked glycopeptides by high energy collision-induced dissociation. J Am Soc Mass Spectrom 7:319–328. Medzihradszky KF, Campbell JM, Baldwin MA, Falick AM, Juhasz P, Vestal ML, Burlingame AL. 2000. The characteristics of peptide collision-induced dissociation using a highperformance MALDI-TOF/TOF tandem mass spectrometer. Anal Chem 72:552–558. Mendonca S, Cole RB, Zhu JH, Cai Y, French AD, Johnson GP, Laine RA. 2003. Incremented alkyl derivatives enhance collision induced glycosidic bond cleavage in mass spectrometry of disaccharides. J Am Soc Mass Spectrom 14:63–78. Meng CK, Mann M, Fenn JB. 1988. Of protons or proteins. Z Phys D 10:361–368. Mirgorodskaya E, Roepstorff P, Zubarev RA. 1999. Localization of O-glycosylation sites in peptides by electron capture dissociation in a Fourier transform mass spectrometer. Anal Chem 71:4431–4436. Mizuno Y, Sasagawa T, Dohmae N, Takio K. 1999. An automated interpretation of MALDI/TOF postsource decay spectra of oligosaccharides. 1. Automated peak assignment. Anal Chem 71:4764–4771. Morris HR, Paxton T, Dell A, Langhorne J, Berg M, Bordoli RS, Hoyes J, Bateman RH. 1996. High sensitivity collisionallyactivated decomposition tandem mass spectrometry on a MASS SPECTROMETRY OF OLIGOSACCHARIDES novel quadrupole/orthogonal-acceleration time-of-flight mass spectrometer. Rapid Commun Mass Spectrom 10: 889–896. Morris HR, Paxton T, Panico M, McDowell R, Dell A. 1997. A novel geometry mass spectrometer, the Q-TOF, for lowfemtomole/attomole-range biopolymer sequencing. J Protein Chem 16:469–479. Mortz E, Sareneva T, Julkunen I, Roepstorff P. 1996. Does matrix-assisted laser desorption ionization mass spectrometry allow analysis of carbohydrate heterogeneity in glycoproteins? A study of natural human interferon-gamma. J Mass Spectrom 31:1109–1118. Moyer SC, Cotter RJ. 2002. Atmospheric pressure MALDI. Anal Chem 74:468A–476A. Moyer SC, Marzilli LA, Woods A, Laiko VV, Doroshenko VM, Cotter RJ. 2002. Atmospheric pressure matrix-assisted laser desorption/ionization (AP MALDI) on a quadrupole ion trap mass spectrometer. Int J Mass Spectrom 226:133– 150. Mueller DR, Domon BM, Blum W, Raschdorf F, Richter WJ. 1988. Direct stereochemical assignment of sugar subunits in naturally occurring glycosides by low energy collision induced dissociation. Application to papulacandin antibiotics. Biomed Environ Mass Spectrom 15:441–446. Müller DR, Domon BM, Blum W, Raschdorf F, Richter WJ. 1988. Direct stereochemical assignment of sugar subunits in naturally occurring glycosides by low energy collision induced dissociation. Application to papulacandin antibiotics. Biomed Environ Mass Spectrom 15:441–446. Nemeth JF, Hochgesang GP, Jr, Marnett LJ, Caprioli RM, Hochensang GP, Jr. 2001. Characterization of the glycosylation sites in cyclooxygenase-2 using mass spectrometry. Biochemistry 40:3109–3116. Nemeth-Cawley JF, Karnik S, Rouse JC. 2001. Analysis of sulfated peptides using positive electrospray ionization tandem mass spectrometry. J Mass Spectrom 36:1301–1311. Nilsson B, Zopf D. 1982. Gas chromatography and mass spectrometry of hexosamine-containing oligosaccharide alditols as their permethylated, N-trifluoroacetyl derivatives. Methods Enzymol 83:46–58. Nilsson B, Zopf D. 1983. Oligosaccharides released from glycolipids by trifluoroacetolysis can be analyzed by gas chromatography-mass spectrometry. Arch Biochem Biophys 222:628–648. O’Connor PB, Mirgorodskaya E, Costello CE. 2002. High pressure matrix-assisted laser desorption/ionization Fourier transform mass spectrometry for minimization of ganglioside fragmentation. J Am Soc Mass Spectrom 13:402– 407. O’Connor PB, Costello CE. 2001. A high pressure matrixassisted laser desorption/ionization Fourier transform mass spectrometry ion source for thermal stabilization of labile biomolecules. Rapid Commun Mass Spectrom 15:1862– 1868. Ohta M, Kawasaki N, Hyuga S, Hyuga M, Hayakawa T. 2001. Selective glycopeptide mapping of erythropoietin by on-line & high-performance liquid chromatography-electrospray ionization mass spectrometry. J Chromatogr A 910:1–11. Okafo G, Langridge J, North S, Organ A, West A, Morris M, Camilleri P. 1997. High-performance liquid chromatographic analysis of complex N-linked glycans derivatized with 2-aminoacridone. Anal Chem 69:4985–4993. Okamoto M, Takahashi K, Doi T, Takimoto Y. 1997. Highsensitivity detection and postsource decay of 2-aminopyridine- derivatized oligosaccharides with matrix-assisted laser desorption/ionization mass spectrometry. Anal Chem 69:2919–2926. Orlando R, Bush CA, Fenselau C. 1990. Structural-analysis of oligosaccharides by tandem mass-spectrometry—Collisional activation of sodium adduct ions. Biomed Environ Mass Spectrom 19:747–754. Overberg A, Karas M, Bahr U, Kaufmann R, Hillenkamp F. 1990. Matrix-assisted infrared-laser (2.94 mm) desorption/ionization mass spectrometry of large biomolecules. Rapid Commun Mass Spectrom 4:293–296. Papac DI, Wong A, Jones AJ. 1996. Analysis of acidic oligosaccharides and glycopeptides by matrix-assisted laser desorption/ionization time-of-flight mass spectrometry. Anal Chem 68:3215–3223. Papayannopoulos IA. 1995. The interpretation of collisioninduced dissociation tandem mass spectra of peptides. Mass Spectrom Rev 14:49–73. Penn SG, Cancilla MT, Lebrilla CB. 2000. Fragmentation behavior of multiple-metal-coordinated acidic oligosaccharides studied by matrix-assisted laser desorption ionization Fourier transform mass spectrometry. Int J Mass Spectrom 196:259–269. Perrault H, Costello CE. 1994. Liquid secondary ionization, tandem and matrix-assisted laser desorption/ionization time-of-flight mass spectrometric characterization of glycosphingolipid derivatives. Org Mass Spectrom 29:720–735. Perreault H, Costello CE. 1996. Differentiation of cerebroside isomers and study of fragmentation by liquid secondary ion mass spectrometry and mass spectrometry mass spectrometry of selected derivatives. Canadian J Chem-Revue Canadienne De Chimie 74:1682–1695. Perreault H, Hronowski XL, Koul O, Street J, McCluer RH, Costello CE. 1997. High sensitivity mass spectral characterization of glycosphingolipids from bovine erythrocytes, mouse kidney and fetal calf brain. Int J Mass Spectrom 169:351–370. Peter-Katalinic J. 1994. Analysis of glycoconjugates by fast atom bombardment mass spectrometry and related MS techniques. Mass Spectrom Rev 13:77–98. Pitt JJ, Gorman JJ. 1996. Matrix-assisted laser desorption/ ionization time-of-flight mass spectrometry of sialylated glycopeptides and proteins using 2,6-dihydroxyacetophenone as a matrix. Rapid Commun Mass Spectrom 10:1786– 1788. Pope RM, Raska CS, Thorp SC, Liu J. 2001. Analysis of heparan sulfate oligosaccharides by nano-electrospray ionization mass spectrometry. Glycobiology 11:505–513. 223 & ZAIA Poulter L, Burlingame AL. 1990. Desorption mass spectrometry of oligosaccharides coupled with hydrophobic chromophores. Methods Enzymol 193:661–689. Powell AK, Harvey DJ. 1996. Stabilization of sialic acids in Nlinked oligosaccharides and gangliosides for analysis by positive ion matrix-assisted laser desorption/ionization mass spectrometry. Rapid Commun Mass Spectrom 10:1027– 1032. Prakobphol A, Thomsson KA, Hansson GC, Rosen SD, Singer MS, Phillips NJ, Medzihradszky KF, Burlingame AL, Leffler H, Fisher SJ. 1998. Human low-molecular-weight salivary mucin expresses the sialyl lewisx determinant and has L-selectin ligand activity. Biochemistry 37:4916–4927. Rademaker GJ, Pergantis SA, Blok-Tip L, Langridge JI, Kleen A, Thomas-Oates JE. 1998. Mass spectrometric determination of the sites of O-glycan attachment with low picomolar sensitivity. Anal Biochem 257:149–160. Reinhold BB, Chan S-Y, Reinhold VN. 1994. Profiling glycosphingolipid structural detail—Periodate-oxidation, electrospray, collision-induced dissociation and tandem massspectrometry. Org Mass Spectrom 29:736–7469. Reinhold VN, Reinhold BB, Costello CE. 1995. Carbohydrate molecular weight profiling, sequence, linkage, and branching data: ES-MS and CID. Anal Chem 67:1772–1784. Reinhold VN, Sheeley DM. 1998. Detailed characterization of carbohydrate linkage and sequence in an ion trap mass spectrometer: Glycosphingolipids. Anal Biochem 259: 28–33. Reinhold VN, Carr SA, Green BN, Petitou M, Choay J, Sinay P. 1987. Structural characterization of sulfated glycosaminoglycans by fast atom bombardment mass spectrometry: application to heparin fragments prepared by chemical synthesis. Carbohydr Res 161:305–313. Rhomberg AJ, Ernst S, Sasisekharan R, Biemann K. 1998. Mass spectrometric and capillary electrophoretic investigation of the enzymatic degradation of heparin-like glycosaminoglycans. Proc Natl Acad Sci USA 95:4176–4181. Rhomberg AJ, Shriver Z, Biemann K, Sasisekharan R. 1998. Mass spectrometric evidence for the enzymatic mechanism of the depolymerization of heparin-like glycosaminoglycans by heparinase II. Proc Natl Acad Sci USA 95:12232– 12237. Ritchie MA, Gill AC, Deery MJ, Lilley K. 2002. Precursor ion scanning for detection and structural characterization of heterogeneous glycopeptide mixtures. J Am Soc Mass Spectrom 13:1065–1077. Roboz J. 1968. Introduction to mass spectrometry: Instrumentation and Techniques. New York, NY: Interscience. Ruiz-Calero V, Moyano E, Puignou L, Galceran MT. 2001. Pressure-assisted capillary electrophoresis-electrospray ion trap mass spectrometry for the analysis of heparin depolymerised disaccharides. J Chromatogr A 914:277– 291. Saba JA, Shen XD, Jamieson JC, Perreault H. 1999. Effect of 1phenyl-3-methyl-5-pyrazolone labeling on the fragmentation behavior of asialo and sialylated N-linked glycans under 224 electrospray ionization conditions. Rapid Commun Mass Spectrom 13:704–711. Sandhoff R, Hepbildikler ST, Jennemann R, Geyer R, Gieselmann V, Proia RL, Wiegandt H, Grone HJ. 2002. Kidney sulfatides in mouse models of inherited glycosphingolipid disorders: determination by nano-electrospray ionization tandem mass spectrometry. J Biol Chem 277:20386–20398. Sato Y, Suzuki M, Nirasawa T, Suzuki A, Endo T. 2000. Microsequencing of glycans using 2-aminobenzamide and MALDI-TOF mass spectrometry: Occurrence of unique linkage-dependent fragmentation. Anal Chem 72:1207– 1216. Schiller J, Arnhold J, Benard S, Reichl S, Arnold K. 1999. Cartilage degradation by hyaluronate lyase and chondroitin ABC lyase: a MALDI-TOF mass spectrometric study. Carbohydr Res 318:116–122. Senko MW, Speir JP, McLafferty FW. 1994. Collisional activation of large multiply-charged ions using Fourier transform mass spectrometry. Anal Chem 66:2801–2808. Severs JC, Carnine M, Eguizabal H, Mock KK. 1999. Characterization of tyrosine sulfate residues in antihemophilic recombinant factor VIII by liquid chromatography electrospray ionization tandem mass spectrometry and amino acid analysis. Rapid Commun Mass Spectrom 13: 1016–1023. Sheeley DM, Reinhold VN. 1998. Structural characterization of carbohydrate sequence, linkage, and branching in a quadrupole ion trap mass spectrometer: Neutral oligosaccharides and N-linked glycans. Anal Chem 70:3053–3059. Shevchenko A, Chernushevich I, Ens W, Standing KG, Thomson B, Wilm M, Mann M. 1997. Rapid ‘de novo’ peptide sequencing by a combination of nanoelectrospray, isotopic labeling and a quadrupole/time-of-flight mass spectrometer. Rapid Commun Mass Spectrom 11:1015–1024. Shi SD, Hemling ME, Carr SA, Horn DM, Lindh I, McLafferty FW. 2001. Phosphopeptide/phosphoprotein mapping by electron capture dissociation mass spectrometry. Anal Chem 73:19–22. Shriver Z, Raman R, Venkataraman G, Drummond K, Turnbull J, Toida T, Linhardt R, Biemann K, Sasisekharan R. 2000. Sequencing of 3-O-sulfate containing heparin decasaccharides with a partial antithrombin III binding site. Proc Natl Acad Sci USA 97:10359–10364. Shriver Z, Sundaram M, Venkataraman G, Fareed J, Linhardt R, Biemann K, Sasisekharan R. 2000. Cleavage of the antithrombin III binding site in heparin by heparinases and its implication in the generation of low molecular weight heparin. Proc Natl Acad Sci USA 97:10365–10370. Sible EM, Brimmer SP, Leary JA. 1997. Interaction of first row transition metals with alpha 1-3, alpha 1-6 mannotriose and conserved trimannosyl core oligosaccharides: A comparative electrospray ionization study of doubly and singly charged complexes. J Am Soc Mass Spectrom 8:32–42. Siegel MM, Tabei K, Kagan MZ, Vlahov IR, Hileman RE, Linhardt R. 1997. Polysulfated carbohydrates analyzed as ion-paired complexes with basic peptides and proteins using MASS SPECTROMETRY OF OLIGOSACCHARIDES electrospray negative ionization mass spectrometry. J Mass Spectrom 32:760–772. Solouki T, Pasa-Tolic L, Jackson GS, Guan S, Marshall AG. 1996. High-resolution multistage MS, MS2, and MS3 matrix-assisted laser desorption/ionization FT-ICR mass spectra of peptides from a single laser shot. Anal Chem 68:3718–3725. Solouki T, Reinhold BB, Costello CE, O’Malley M, Guan S, Marshall AG. 1998. Electrospray ionization and matrixassisted laser desorption/ionization Fourier transform ion cyclotron resonance mass spectrometry of permethylated oligosaccharides. Anal Chem 70:857–864. Spengler B, Dolce JW, Cotter RJ. 1990. Infrared-laser desorption mass-spectrometry of oligosaccharides—Fragmentation mechanisms and isomer analysis. Anal Chem 62:1731– 1737. Spengler B, Kirsch D, Kaufmann R. 1991. Metastable decay of peptides and proteins in matrix-assisted laser-desorption mass-spectrometry. Rapid Commun Mass Spectrom 5:198– 202. Spengler B, Kirsch D, Kaufmann R, Lemoine J. 1994. Structureanalysis of branched oligosaccharides using post-source decay in matrix-assisted laser-desorption ionization massspectrometry. Org Mass Spectrom 29:782–787. Steen H, Küster B, Mann M. 2001. Quadrupole time-of-flight versus triple-quadrupole mass spectrometry for the determination of phosphopeptides by precursor ion scanning. J Mass Spectrom 36:782–790. Stensballe A, Jensen ON, Olsen JV, Haselmann KF, Zubarev RA. 2000. Electron capture dissociation of singly and multiply phosphorylated peptides. Rapid Commun Mass Spectrom 14:1793–1800. Strecker G, Wieruszeski J-M, Martel C, Montreuil J. 1987. Determination of the structure of sulfated tetra- and pentaoligosaccharides obtained by alkaline borohydride degradation of hen ovomucin. A fast atom bombardmentmass spectrometric study. Glycoconj J 4:329–337. Sturiale L, Naggi A, Torri G. 2001. MALDI mass spectrometry as a tool for characterizing glycosaminoglycan oligosaccharides and their interaction with proteins. Semin Thromb Hemost 27:465–472. Sugahara K, Takemura Y, Sugiura M, Kohno Y, Yoshida K, Takeda K, Khoo KH, Morris HR, Dell A. 1994. Chondroitinase ABC-resistant sulfated trisaccharides isolated from digests of chondroitin/dermatan sulfate chains. Carbohydr Res 255:165–182. Sugahara K, Tanaka Y, Yamada S, Seno N, Kitagawa H, Haslam SM, Morris HR, Dell A. 1996. Novel sulfated oligosaccharides containing 3-O-sulfated glucuronic acid from king crab cartilage chondroitin sulfate K. Unexpected degradation by chondroitinase ABC. J Biol Chem 271: 26745–26754. Tadano-Aritomi K, Kubo H, Ireland P, Okuda M, Kasama T, Handa S, Ishizuka I. 1995. Structural analysis of monoand bis-sulfated glycosphingolipids by negative liquid secondary ion mass spectrometry with high- and low- & energy collision-induced dissociation. Carbohydr Res 273: 41–52. Taguchi T, Iwasaki M, Muto Y, Kitajima K, Inoue S, Khoo KH, Morris HR, Dell A, Inoue Y. 1996. Occurrence and structural analysis of highly sulfated multiantennary Nlinked glycan chains derived from a fertilization-associated carbohydrate-rich glycoprotein in unfertilized eggs of Tribolodon hakonensis. Eur J Biochem 238:357–367. Takagaki K, Kojima K, Majima M, Nakamura T, Kato I, Endo M. 1992. Ion-spray mass spectrometric analysis of glycosaminoglycan oligosaccharides. Glycoconj J 9:174–179. Takao T, Tambara Y, Nakamura A, Yoshino K, Fukuda H, Fukuda M, Shimonishi Y. 1996. Sensitive analysis of oligosaccharides derivatized with 4-aminobenzoic acid 2-(diethylamino)ethyl ester by matrix-assisted laser desorption/ionization mass spectrometry. Rapid Commun Mass Spectrom 10: 637–640. Talbo G, Roepstorff P. 1993. Determination of sulfated peptides via prompt fragmentation by UV matrix-assisted laser desorption/ionization mass spectrometry. Rapid Commun Mass Spectrom 7:201–204. Tang XJ, Ens W, Standing KG, Westmore JB. 1988. Daughter ion mass spectra from cationized molecules of small oligopeptides in a reflecting time-of-flight mass spectrometer. Anal Chem 60:1791–1799. Tarelli E, Byers HL, Wilson M, Roberts G, Homer KA, Beighton D. 2000. Detecting mannosidase activities using ribonuclease B and matrix-assisted laser desorption/ionizationtime of flight mass spectrometry. Anal Biochem 282:165– 172. Tetaert D, Briand G, Soudan B, Richet C, Demeyer D, Boersma A, Degand P. 1994. Analysis by electrospray mass spectrometry of glycopeptides from the in vitro O-glycosylation reaction using human mucin motif peptide. Anal Biochem 222:409–416. Todd JFJ. 1991. Ion trap mass spectrometer–Past, present, and future(?). Mass Spectrom Rev 10:3–52. Tseng K, Hedrick JL, Lebrilla CB. 1999. Catalog-library approach for the rapid and sensitive structural elucidation of oligosaccharides. Anal Chem 71:3747–3754. Tseng K, Lindsay LL, Penn S, Hedrick JL, Lebrilla CB. 1997. Characterization of neutral oligosaccharide-alditols from Xenopus laevis egg jelly coats by matrix-assisted laser desorption Fourier transform mass spectrometry. Anal Biochem 250:18–28. Venkataraman G, Shriver Z, Davis JC, Sasisekharan R. 1999a. Fibroblast growth factors 1 and 2 are distinct in oligomerization in the presence of heparin-like glycosaminoglycans. Proc Natl Acad Sci USA 96:1892–1897. Venkataraman G, Shriver Z, Raman R, Sasisekharan R. 1999b. Sequencing complex polysaccharides. Science 286:537– 542. Viseux N, Costello CE, Domon B. 1999. Post-source decay mass spectrometry: optimized calibration procedure and structural characterization of permethylated oligosaccharides. J Mass Spectrom 34:364–376. 225 & ZAIA Viseux N, de Hoffmann E, Domon B. 1997. Structural analysis of permethylated oligosaccharides by electrospray tandem mass spectrometry. Anal Chem 69:3193–3198. Viseux N, de Hoffmann E, Domon B. 1998. Structural assignment of permethylated oligosaccharide subunits using sequential tandem mass spectrometry. Anal Chem 70:4951– 4959. Viseux N, Hronowski X, Delaney J, Domon B. 2001. Qualitative and quantitative analysis of the glycosylation pattern of recombinant proteins. Anal Chem 73:4755–4762. Wang WT, Matsuura F, Sweeley CC. 1983. Chromatography of permethylated oligosaccharide alditols. Anal Biochem 134: 398–405. Wang WT, LeDonne NC, Jr, Ackerman B, Sweeley CC. 1984. Structural characterization of oligosaccharides by highperformance liquid chromatography, fast-atom bombardment-mass spectrometry, and exoglycosidase digestion. Anal Biochem 141:366–381. Weiskopf AS, Vouros P, Harvey DJ. 1997. Characterization of oligosaccharide composition and structure by quadrupole ion trap mass spectrometry. Rapid Commun Mass Spectrom 11:1493–1504. Weiskopf AS, Vouros P, Harvey DJ. 1998. Electrospray ionization-ion trap mass spectrometry for structural analysis of complex N-linked glycoprotein oligosaccharides. Anal Chem 70:4441–4447. Wheeler SF, Harvey DJ. 2000. Negative ion mass spectrometry of sialylated carbohydrates: discrimination of N-acetylneuraminic acid linkages by MALDI-TOF and ESI-TOF mass spectrometry. Anal Chem 72:5027–5039. Wheeler SF, Harvey DJ. 2001. Extension of the in-gel release method for structural analysis of neutral and sialylated Nlinked glycans to the analysis of sulfated glycans: Application to the glycans from bovine thyroid-stimulating hormone. Anal Biochem 296:96–100. Wilm M, Neubauer G, Mann M. 1996. Parent ion scans of unseparated peptide mixtures. Anal Chem 68:527–533. Wilm MS, Mann M. 1994. Electrospray and Taylor-cone theory, Dole’s beam of macromolecules at last? Int J Mass Spectrom Ion Processes 136:167–180. Wolfender JL, Chu F, Ball H, Wolfender F, Fainzilber M, Baldwin MA, Burlingame AL. 1999. Identification of tyrosine sulfation in Conus pennaceus conotoxins alpha-PnIA and alpha-PnIB: Further investigation of labile sulfo- and phosphopeptides by electrospray, matrix-assisted laser desorption/ionization (MALDI) and atmospheric pressure MALDI mass spectrometry. J Mass Spectrom 34:447–454. Wong AW, Wang H, Lebrilla CB. 2000. Selection of anionic dopant for quantifying desialylation reactions with MALDIFTMS. Anal Chem 72:1419–1425. Wong AW, Cancilla MT, Voss LR, Lebrilla CB. 1999. Anion dopant for oligosaccharides in matrix-assisted laser desorption/ionization mass spectrometry. Anal Chem 71:205– 211. Wu QY, Vanorden S, Cheng XH, Bakhtiar R, Smith RD. 1995. Characterization of cytochrome-C variants with high- 226 resolution FTICR mass-spectrometry—Correlation of fragmentation and structure. Anal Chem 67:2498–2509. Yagami T, Kitagawa K, Futaki S. 1995. Liquid secondary-ion mass spectrometry of peptides containing multiple tyrosineO-sulfates. Rapid Commun Mass Spectrom 9:1335–1341. Yagami T, Kitagawa K, Aida C, Fujiwara H, Futaki S. 2000. Stabilization of a tyrosine O-sulfate residue by a cationic functional group: Formation of a conjugate acid-base pair. J Pept Res 56:239–249. Yamada S, Yoshida K, Sugiura M, Sugahara K, Khoo KH, Morris HR, Dell A. 1993. Structural studies on the bacterial lyase-resistant tetrasaccharides derived from the antithrombin III-binding site of porcine intestinal heparin. J Biol Chem 268:4780–4787. Yamagaki T, Nakanishi H. 1999. A new technique distinguishing alpha2-3 sialyl linkage from alpha2-6 linkage in sialyllactoses and sialyl-N-acetyllactosamines by post-source decay fragmentation method of MALDI-TOF mass spectrometry. Glycoconj J 16:385–389. Yamagaki T, Nakanishi H. 2000. Post-source decay fragmentation analyses of linkage isomers of Lewis-type oligosaccharides in curved-field reflection matrix-assisted laser desorption/ionization time-of-flight mass spectrometry: Combined in-source decay/post-source decay experiments and relative ion abundance analysis. J Mass Spectrom 35: 1300–1307. Yang Y, Bergmann C, Benen J, Orlando R. 1997. Identification of the glycosylation site and glycan structures of recombinant endopolygalacturonase II by mass spectrometry. Rapid Commun Mass Spectrom 11:1257–1262. Yang HO, Gunay NS, Toida T, Kuberan B, Yu G, Kim YS, Linhardt RJ. 2000. Preparation and structural determination of dermatan sulfate-derived oligosaccharides. Glycobiology 10:1033–1039. Yergey AL, Coorssen JR, Backlund PS, Jr, Blank PS, Humphrey GA, Zimmerberg J, Campbell JM, Vestal ML. 2002. De novo sequencing of peptides using MALDI/TOF-TOF. J Am Soc Mass Spectrom 13:784–791. Yu G, LeBrun L, Gunay NS, Hoppensteadt D, Walenga JM, Fareed J, Linhardt RJ. 2000. Heparinase I acts on a synthetic heparin pentasaccharide corresponding to the antithrombin III binding site. Thromb Res 100:549–556. Zaia J, McClellan JE, Costello CE. 2001. Tandem mass spectrometric determination of the 4S/6S sulfation sequence in chondroitin sulfate oligosaccharides. Anal Chem 73:6030– 6039. Zaia J, Costello CE. 2001. Compositional analysis of glycosaminoglycans by electrospray mass spectrometry. Anal Chem 73:233–239. Zaia J, Costello CE. 2003. Tandem mass spectrometry of sulfated heparin-like glycosaminoglycan oligosaccharides. Anal Chem 75:2445–2455. Zaia J, Boynton RE, McIntosh A, Marshak DR, Olsson H, Heinegård D, Barry FP. 1997. Post-translational modifications in cartilage oligomeric matrix protein. Characterization of the N-linked oligosaccharides by matrix-assisted MASS SPECTROMETRY OF OLIGOSACCHARIDES laser desorption ionization time-of-flight mass spectrometry. J Biol Chem 272:14120–14126. Zaia J, Liu B, Boynton R, Barry F. 2000. Structural analysis of cartilage proteoglycans and glycoproteins using matrixassisted laser desorption/ionization time-of-flight mass spectrometry. Anal Biochem 277:94–103. Zaia J, Boynton R, Heinegård D, Barry F. 2001. Posttranslational modifications to human bone sialoprotein determined by mass spectrometry. Biochemistry 40:12983–12991. Zamfir A, Seidler DG, Kresse H, Peter-Katalinc J. 2002. Structural characterization of chondroitin/dermatan sulfate oligosaccharides from bovine aorta by capillary electrophoresis and electrospray ionization quadrupole time-offlight tandem mass spectrometry. Rapid Commun Mass Spectrom 16:2015–2024. Zamze S, Wing DR, Wormald MR, Hunter AP, Dwek RA, Harvey DJ. 2001. A family of novel, acidic N-glycans in Bowes melanoma tissue plasminogen activator have L2/HNK-1bearing antennae, many with sulfation of the fucosylated chitobiose core. Eur J Biochem 268:4063–4078. & Zeng R, Xu Q, Shao XX, Wang KY, Xia QC. 1999. Characterization and analysis of a novel glycoprotein from snake venom using liquid chromatography-electrospray mass spectrometry and Edman degradation. Eur J Biochem 266:352–358. Zhou ZR, Ogden S, Leary JA. 1990. Linkage position determination in oligosaccharides—MS/MS study of lithiumcationized carbohydrates. J Org Chem 55:5444–5446. Zhu X, Borchers C, Bienstock RJ, Tomer KB. 2000. Mass spectrometric characterization of the glycosylation pattern of HIV-gp120 expressed in CHO cells. Biochemistry 39:11194–11204. Zubarev RA, Kelleher NL, McLafferty FW. 1998. Electron capture dissociation of multiply charged protein cations. A nonergodic process. J Am Chem Soc 120:3265–3266. Zubarev RA, Kruger NA, Fridriksson EK, Lewis MA, Horn DM, Carpenter BK, McLafferty FW. 1999. Electron capture dissociation of gaseous multiply-charged proteins is favored at disulfide bonds and other sites of high hydrogen atom affinity. J Am Chem Soc 121:2857–2862. 227