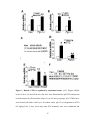
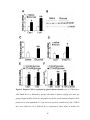
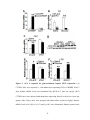
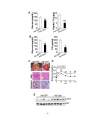
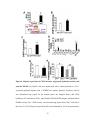
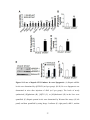
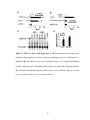
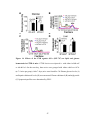
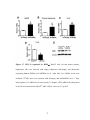
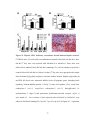
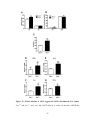
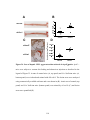
Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Expression vector wikipedia , lookup
Lipid signaling wikipedia , lookup
Fatty acid synthesis wikipedia , lookup
Biochemistry wikipedia , lookup
Endocannabinoid system wikipedia , lookup
Wilson's disease wikipedia , lookup
Specialized pro-resolving mediators wikipedia , lookup
Fatty acid metabolism wikipedia , lookup
CARBOXYLESTERASE 1 PLAYS A PROTECTIVE ROLE AGAINST METABOLIC DISEASE A dissertation submitted to Kent State University in partial fulfillment of the requirements for the degree of Doctor of Philosophy By Jiesi Xu May 2016 © Copyright All rights reserved Except for previously published materials Dissertation written by Jiesi Xu B.S., Jilin University 2008 Ph.D., Kent State University 2016 Approved by ___________________________________ , Chair, Doctoral Dissertation Committee Dr. Yanqiao Zhang, M.D., Associate Professor, NEOMED ___________________________________ , Member, Doctoral Dissertation Committee Dr. John Y.L. Chiang Ph.D., Distinguished Professor, NEOMED ___________________________________, Member, Doctoral Dissertation Committee Dr. Colleen M. Novak, Ph.D., Associate Professor, Kent State University ___________________________________, Member, Doctoral Dissertation Committee Dr. Min You, Ph.D., Professor, NEOMED ___________________________________, Member, Doctoral Dissertation Committee Dr. Eric M. Mintz, Ph.D., Professor, Associate dean, Kent State University Accepted by ___________________________________ , Director, School of Biomedical Sciences Dr. Ernest Freeman, Ph.D. ___________________________________ , Dean, College of Arts and Sciences Dr. James L. Blank, Ph.D ii TABLE OF CONTENTS TABLE OF CONTENTS LIST OF FIGURES ............................................................................................................v LIST OF ABBREVIATIONS .........................................................................................viii ACKNOWLEDGEMENTS ...............................................................................................x ABSTRACT ......................................................................................................................xi CHAPTER 1 THE ROLE OF CARBOXYLESTERASE 1 IN NON-ALCOHOLIC FATTY LIVER DISEASE AND CARBOHYDRATE METABOLISM………………...1 1.1 INTRODUCTION……………........................................................................ 1 1.2 METHODS…………........................................................................................7 1.3 RESULTS .......................................................................................................14 1.3.1 Hepatic Carboxylesterase1 is Induced by Glucose and Regulates Postprandial Glucose Levels................................................................................. 14 1.3.2 Hepatic Carboxylesterase 1 is Essential for Normal and Farnesoid X Receptor-Controlled Lipid Homeostasis................................................................25 1.4 DISCUSSION……………………..................................................................50 CHPTER 2 THE ROLE OF CARBOXYLESTERASE 1 IN ALCOHOLIC LIVER DISEASE………………………………...…................................................................... 56 2.1 INTRODUCTION.......................................................................................... 56 2.2 METHODS..................................................................................................... 59 2.3 RESULTS....................................................................................................... 67 iii 2.4 DISCUSSION ................................................................................................ 96 CHAPTER 3 THE ROLE OF CARBOXYLESTERASE 1 IN ATHEROSCLEROSIS…………………………............................................................100 3.1 INTRODUCTION.........................................................................................100 3.2 METHODS....................................................................................................103 3.3 RESULTS......................................................................................................105 3.4 DISCUSSION ...............................................................................................111 CHAPTER 4 CONCLUSION…….……………............................................................113 REFERENCES…………………………………………………………………………116 iv LIST OF FIGURES Figure 1. Hepatic CES1 is regulated by nutritional status……………………………….15 Figure 2. Hepatic CES1 is regulated by glucose but not insulin………………………...18 Figure 3. ACL is required for glucose-induced hepatic CES1 expression………………21 Figure 4. ACL is required for glucose-mediated acetylation of histones (H3, H4) in the CES1 chromatin………………………………………………………….…….22 Figure 5. CES1 regulates postprandial levels…………………………………………....24 Figure 6. Hepatic expression of CES1 lowers hepatic triglyceride levels and improves glucose homeostasis………………………………………………………...26,27 Figure 7. Hepatic expression of CES1 selectively regulates gene expression and has no effect on lipogenesis or VLDL secretion………………………………………30 Figure 8. Hepatic expression of CES1 increases triglyceride hydrolase activity and activates PPARα……………………………………………………………….32 Figure 9. Loss of hepatic CES1 causes fatty liver and increased plasma cholesterol level…………………………………………………………………………….35 Figure 10. Loss of hepatic CES1 induces de novo lipogenesis………………………….37 Figure 11. Hepatic CES1 is regulated by FXR…………………………………………..40 Figure 12. CES1 is a direct FXR target gene…………………………………………….43 Figure 13. Essential roles of hepatic CES1 in FXR-regulated lipid homeostasis………..45 Figure 14. Effects of the FXR agonist OCA (INT-747) on lipid and glucose homeostasis in C57BL/6 mice……………………………………………………………….47 Figure 15. Effects of the FXR agonist OCA on lipid homeostasis in ob/ob mice……….49 v Figure 16. CES1 and HNF4 expressions are reduced in patients with alcoholic steatohepatitis and in mouse and mouse primary hepatocytes treated with ethanol………………………………………………………………………….68 Figure 17. CES1 is regulated by HNF4………………………………………………...71 Figure 18. CES1 is a direct target of HNF4……………………………………………72 Figure 19. Over-expression of hepatic CES1 protects against alcohol-induced triglyceride accumulation in AML12 cells………………………………………………….74 Figure 20. Hepatic CES1 deficiency alters plasma lipid levels in response to alcohol challenge……………………………………………………………………….75 Figure 21. Hepatic CES1 deficiency exacerbates alcohol-induced hepatic steatosis……77 Figure 22. Hepatic CES1 deficiency exacerbates alcohol-induced liver inflammation…………………………………………………………………...80 Figure 23. Global deletion of CES1 does not exacerbate alcohol-induced hepatic steatosis...………………………………………………………………………83 Figure 24. Global deletion of CES1 exacerbates alcohol-induced liver inflammation….85 Figure 25. Global deletion of CES1 increases MCD diet-induced liver inflammation………………………………………………………..………….88 Figure 26. CES1 deficiency does not significantly change fibrogenic gene expressions……………………………………………………………………..88 Figure 27. Global deletion of CES1 does not exacerbate MCD-diet induced fibrosis….............................................................................................................89 Figure 28. Global deletion of CES1 increases hepatic acetaldehyde level and oxidative stress……………………………………………………………………………92 vi Figure 29. Global deletion of CES1 does not change mRNA levels of genes involved in fatty acid metabolism…………………………………………………………..95 Figure 30. Macrophage cholesterol efflux……………………………………………...102 Figure 31. Global deletion of CES1 results in increased lipid accumulation in macrophages………………………………………………………………….105 Figure 32. Loss of hepatic CES1 increases lipid contents in ApoE mice……………106 Figure 33. Loss of hepatic CES1 shows atherosclerotic lipid profile…………………..108 Figure 34. Loss of hepatic CES1 aggravates atherosclerosis in ApoE mice…………110 vii LIST OF ABBREVIATIONS ABCA1 ATP binding cassette sub-family A1 ABCG5 ATP binding cassette sub-family G5 ACC acetyl-CoA carboxylase ACL ATP citrate lyase AKT protein kinase B ALD alcoholic liver disease APOB apolipoprotein B CD36 cluster of differentiation 36 CES1 carboxylesterase 1 ChIP chromatin immunoprecipitation CPT carnitine palmitoyltransferase DGAT diacylglycerol O-acyltransferase EMSA electrophoretic mobility shift assay FAS fatty acid synthase FFA free fatty acid FPLC fast protein liquid chromatography FXR farnesoid X receptor GCK glucose kinase G6Pase glucose-6 phosphatase HDL high density lipoprotein HMGCS HMG-CoA synthase viii HMGCR HMG-CoA reductase HNF4 hepatocyte nuclear factor 4 IL-1 interleukin 1 IL-6 interleukin 6 LDL low density lipoprotein L-PK liver pyruvate kinase MCP monocyte chemoattractant protein MDA malondialdehyde MTP microsomal triglyceride transfer protein NAFLD non-alcoholic fatty liver disease PPAR peroxisome proliferator-activated receptor PDK pyruvate dehydrogenase kinase PEPCK phosphoenolpyruvate carboxykinase PGC1 peroxisome proliferator-activated receptor coactivator 1 ROS reactive oxygen species SREBP1C sterol regulatory binding protein 1c TC total cholesterol TNF tumor necrosis factor TG triglyceride VLDL very low density lipoprotein ix ACKNOWLEDGEMENTS I would like to express my gratitude to all the people who gave me help and support throughout the course of this work. I would especially like to thank my advisor Dr. Yanqiao Zhang for his invaluable guidance, caring and immense knowledge. I would also like to thank him for pushing me father than I thought I could go and providing financial support for my research. I wish to express my sincere thank to Dr. John Chiang, Dr. Colleen Novak and Dr. Min You for giving me insightful suggestions regarding my research. Their guidance and encouragements help my research from various perspectives. I would like to acknowledge and thank Dr. Novak for the help she has given with CLAMS analysis. My special thanks go to Dr. Yang Xu and Dr. Preeti Pathak for their selfless, enormous help. I am so grateful to Dr. Xu for teaching me the techniques that are important for completing this study; and to Dr. Pathak for lending me her shoulder to cry on. This dissertation is dedicated to my mother who is always there for me and experiences all the peaks and valleys with me during this journey. Even so far away, I feel her love and care every minute in my life. x ABSTRACT Jiesi Xu Department of Biomedical Sciences Kent State University, 2016 Advisor Yanqiao Zhang Carboxylesterase 1 (CES1) is a phase I drug metabolizing enzyme which is shown to hydrolyze ester and amide-containing drugs and prodrugs. It also hydrolyzes triglyceride and cholesteryl ester. The present studies show that glucose induces CES1 expression, and that the induction of CES1 expression, in turn, reduces plasma glucose level, likely through increasing insulin sensitivity. We then demonstrate that overexpression of hepatic CES1 reduces hepatic triglyceride level and promotes fatty acid oxidation. In contrast, loss of hepatic CES1 induces hepatic steatosis by increasing sterol regulatory-element binding protein (SREBP) processing and lipogenesis. We also find that CES1 is a farnesoid X receptor (FXR) target gene, and that activation of FXR reduces hepatic and plasma triglyceride levels through, at least in part, inducing CES1. These results suggest that CES1 plays a protective role against non-alcoholic fatty liver disease (NAFLD). We further show that CES1 is inhibited by alcohol, and is a hepatocyte nuclear factor 4 (HNF4) direct target. Using liver specific Ces1 deficient and Ces1 mice together with chronic-binge alcohol feeding, we find that CES1 deficiency exacerbates alcohol-induced liver inflammation. CES1 deficiency also results in increased hepatic acetaldehyde level, elevated reactive oxygen species (ROS) level and enhanced lipid peroxidation, suggesting that CES1 plays a protective role against xi alcoholic liver disease (ALD). Lastly, we demonstrate that loss of hepatic CES1 aggravates western diet-induced atherosclerosis in ApoE mice. These exciting results lead us to conclude that CES1 plays an essential role in lipid and carbohydrate metabolism, and that it also protects against NAFLD, ALD and atherosclerosis. Targeting CES1 may be a plausible strategy for treating metabolic disease. xii CHAPTER 1: THE ROLE OF CARBOXYLESTERASE 1 IN NON-ALCOHOLIC FATTY LIVER DISEASE AND CARBOHYDRATE METABOLISM 1.1 INTRODUCTION Nonalcoholic fatty liver disease (NAFLD), one of the most common liver diseases worldwide, encompasses a spectrum of liver disorders ranging from simple hepatic steatosis, nonalcoholic steatohepatitis (NASH), to cirrhosis. NAFLD is often associated with insulin resistance, obesity, type 2 diabetes and dyslipidemia, and is often a hepatic manifestation of metabolic disorders [1]. The worldwide prevalence of NAFLD and its subtype nonalcoholic steatohepatitis (NASH) range from 6.3% to 33% and from 3% to 5% in the general population, respectively [2]. Non alcoholic fatty liver (NAFL) is defined as the presence of hepatic steatosis with no evidence of hepatocellular injury in the form of ballooning of the hepatocytes. In contrast, NASH is defined as the presence of hepatic steatosis and inflammation with hepatocyte injury (ballooning) with or without fibrosis. Liver fibrosis is characterized by over-stimulation of liver stellate cells and excessive accumulation of extracellular matrix proteins, thus results in portal hypertension, cirrhosis and liver failure. Cirrhosis, featured by the development of broad collagen bands that form nodules, is irreversible and predicted to become the most common indication for liver transplantation. The estimated risk of progression to cirrhosis in patients with NASH and fibrosis is approximately 20%, compared with only 4% risk of progression in 1 patients with simple hepatic steatosis [3]. Our knowledge of the pathogenesis of NAFLD has greatly advanced. Risks factors including dietary fat, genetic predisposition, gut microbiome and metabolic disorders such as type 2 diabetes, obesity and cardiovascular diseases can often contribute to the development of NAFLD. Hepatic steatosis is a hallmark in NAFLD and is often associated with disrupted glucose and lipid homeostasis. In human body, approximately 60% of the triglyceride accumulated in the liver is derived from plasma non-esterified fatty acid (NEFA) pool, 25% derived from hepatic de novo lipogenesis and 15% derived from dietary intake [3]. Adipose tissue is the largest supplier of fatty acids and generates multiple signals that alter lipid and glucose metabolism. Free fatty acids (FFAs), released from adipose tissue, can be used for triglyceride synthesis or broken down in response to energy demands. Peripheral insulin resistance, a predisposing factor for hepatic steatosis, enhances adipocyte lipolysis and increases the efflux of free fatty acids to the plasma NEFA pool [3,4]. Although hepatic de novo synthesis only accounts for 20% of the lipid pool in the liver, alteration of de novo synthetic gene expressions are often present in patients with metabolic diseases. Hyperinsulinemia and hyperglycemia induce sterol regulatory element-binding protein 1c (SREBP-1c) and carbohydrate response element binding protein (ChREBP) in the liver, subsequently activate lipogenic genes and increase hepatic de novo lipogenesis [2]. The mechanism underlying the disease progression from simple hepatic steatosis to NASH, and to more advanced fibrosis and cirrhosis remains elusive. Proinflammatory cytokines in addition to hepatic steatosis imposes a second “hit” to incite hepatic injury, through sensitizing kupffer cells and triggering apoptosis and necrosis. Oxidative stress, 2 mitochondria dysfunction, endoplasmic reticulum stress, lipotoxicity and apoptotic pathways advance liver damage toward fibrosis and cirrhosis [4]. It is reported that less than 4% of individuals with steatosis progress to cirrhosis, as compared with 20% of individuals with NASH [3]. Currently, no pharmacological therapy is proven to be effective to alter the nature history of NAFLD. Thus, weight loss is often recommended to patients to improve liver biochemistries and histology [5]. The pharmaceutical intervention for NAFLD is directed toward correction of its risk factors. Insulin-sensitizing drugs including thiazolidinediones, rosiglitazone, pioglitazone, and metformin are shown to improve insulin sensitivity, serum alanine transaminase (ALT), and histologic features in the livers of NASH patients without diabetes [6,7,8]. Vitamin E decreases oxidative stress, which provides a rationale for its use not only in patients with NASH but also those with liver fibrosis [9]. Pioglitazone combined with vitamin E is more effective to reverse histologic abnormalities than vitamin E alone [5]. Anti-inflammatory (corticosteroids), and antioxidants (silymarin, phosphatidylcholine, and s-adenosyl-L-methionine) are shown to attenuate experimental liver fibrosis [10]. In addition, pentoxifylline (phosphodiesterase inhibitor), amiloride (Na+/H+ pump inhibitor), S-farnesylthiosalicylic acid (Ras antagonist), thiazolidinediones (peroxisome proliferator-activated receptor ligands), and Renin-angiotensin inhibitors are shown to have antifibrotic effect [10]. Patients with end-stage liver disease are subject to liver transplantation. However, transplantation per se does not correct some metabolic syndrome. Thus, weight management and correction of hyperglycemia and hyperlipidemia become crucial goals for managing NAFLD. 3 Farnesoid X receptor (FXR) is a key element in regulation of bile acid synthesis, glucose and lipid metabolism, energy homeostasis and inflammation pathways. It is a member of the nuclear receptor superfamily and is known to be activated by bile acids [11]. The beneficial effects of activated FXR have been well documented [12,13,14,15]. It lowers hepatic and plasma triglyceride level and plasma cholesterol level, and improves insulin sensitivity [14,15]. These beneficial effects of activation of FXR provide rationales for its use in treating NAFLD. Currently, INT747, a FXR agonist, is undergoing clinical trial for patients with NASH. The hepatoprotective mechanism of FXR agonism lies in two hypotheses: 1) activation of hepatic FXR reduces bile acid synthesis, increases hepatic content of cholesterol, and reduces SREBP1 activity and triglyceride synthesis [16] 2) activation of intestinal FXR increases fibroblast growth factor (FGF)15 secretion, which in turn improves hepatic steatosis and insulin sensitivity [17,18]. In addition, FXR agonism decreases gluconeogenesis, increases glycogen synthesis and increases insulin sensitivity, which leads to decreases in the hepatic content of lipids and overall circulating glucose [19]. Interestingly, disruption intestinal FXR increases glucagon-like peptide-1 (GLP-1), which leads to increased insulin sensitivity [20]. Activation of FXR also reduces diet-induced weight gain, and enhances energy expenditure and browning of white adipose tissue [18,21]. More and more FXR target genes have been reported. Recent studies demonstrate that the hepatoprotective effect of FXR results from activating its target genes, including small heterodimer partner (SHP) [16], PPAR, and Apolipoprotein CII (ApoC-II) [23], and FGF15 [18]. To date, the tissue specific function of FXR in regulation of lipid and glucose metabolism is still obscure. Our group finds that INT747 enhances reverse cholesterol transport through 4 activation of hepatic FXR but not intestinal FXR and subsequent inhibition of cholesterol 7-hydroxylase (Cyp7A1), reduction of bile acids pool size and reduction of intestinal cholesterol absorption. In 2014, Gonzalez, et.al. reported that intestine specific Fxr disruption decreases hepatic triglyceride accumulation, through lowering circulating ceramides, which in turn downregulate hepatic SREBP1C and subsequently decrease de novo lipogenesis [24]. Carboxylesterase 1 (CES1) is a drug-metabolizing enzyme that is highly expressed in the liver but also to a lesser extent in the intestine, macrophages and other tissues [25]. It is a member of mammalian carboxylesterases family which is localized in the endoplasmic reticulum and catalyzes the hydrolysis of a variety of ester and amidecontaining chemicals, drugs (including prodrugs), and endogenous compounds (including acyl-glycerols, long-chain acyl-carnitine and long-chain acyl-coenzyme A ester), to their respective free acids [26]. Drug-metabolizing enzymes, present predominantly in the liver, are involved in detoxification and biotransformation of both endogenous and exogenous compounds to hydrophilic products to facilitate their elimination [27]. CES enzymes catalyzes phase I drug-metabolizing reactions for angiotensin-converting enzyme inhibitors (temocapril, cilazapril, quinapril, and imidapril) [28,29,30], anti-tumor drugs (CPT-11 and Capecitabine) [31,32,33,34,35,36] and narcotics (cocaine, heroin and meperidine) [37,38]. The gene structures of CES family reveal that the murine CES gene is located on the minus strand of chromosome 8 at 8C5 in a cluster of six CES genes that span 260.6kb in total, and that CES families share several common binding sites for transcription factors in the promoter region, suggesting that orthologous CES genes have evolutionally conserved transcriptional regulatory patterns [39]. 5 The physiological functions of CESs have been well documented. So far, CESs families have been shown to be involved in metabolizing a number of drugs and prodrugs such as anti-coagulant [40,41], anti-cancer [33,35,42,43,44], anti-virus[45,46,47] and anti-hypertensive[48,49,50] drugs. CESs possesses triglyceride (TG) and cholesteryl ester (CE) hydrolase activity, and acyl-coenzyme A: cholesterol acyltransferase activity [51,52,53]. Ablation of hepatic CES3 reduces very low-density lipoprotein (VLDL) assembly, decreases plasma TG and cholesterol (CHOL) levels without affecting glucose tolerance [54]. However, global ablation of CES3/TGH leads to improved systemic glucose clearance and insulin sensitivity, decreased pancreatic islet size and decreased expression of hepatic gluconeogenesis-related genes [55]. CES1 is robustly expressed in human THP-1 monocytes/macrophages, and loss of macrophagic CES1 enhances retention of intracellular cholesteryl esters and leads to a “foamy” phenotype, suggesting that CES1 is involved in the development of atherosclerosis [56]. Over-expression of human CES1 in the macrophage leads to increased CE hydrolysis [51], elevated mobilization of cytoplasmic CE [57], enhanced free cholesterol (FC) efflux [58] and attenuation of atherosclerosis in Ldlrmice [51]. The TG and CE hydrolase property of CES1 (ESx) makes it an important target in treating NAFLD [52]. In 2012, Lehner et. al. reported that Ces1mice develop obesity, fatty liver, hyperinsulinemia, and increased cholymicron production [59,60]. In this chapter, the role of CES1 in regulating NAFLD and carbohydrate metabolism is delineated. 6 1.2 METHODS Mice, Diets, and Ligands. C57BL/6 mice, leptin deficient (ob/ob) mice, leptin receptor deficient (db/db) mice, and Fxr mice were purchased from the Jackson Laboratories (Bar Harbor, ME). High fat/high cholesterol (HFHC) diet (40% kcal from fat, 1.5% cholesterol) was purchased from Research Diets (cat #D12108, New Brunswick, NJ). For FXR ligands treatment, FXR agonists GW4064 (30mg/kg, twice a day) and OCA (INT-747) (30mg/kg/d) were administered to Fxr wild type and Fxrmice by gavage. For glucose treatment, C57BL/6 mice were fasted for 16 h, and 40% glucose (8g/kg) was administered twice with 3 h interval through oral gavage. Unless otherwise stated, male mice were used and all mice were fasted for 5-6 hours prior to euthanization. All the animal studies have been approved by the Institutional Animal Care and Use Committee at Northeast Ohio Medical University. Real-Time PCR. RNA was isolated using TRIzol Reagent (Invitrogen, Carlsbad, CA). Messenger RNA (mRNA) levels were determined by quantitative reverse-transcription polymerase chain reaction (qRT-PCR) on a 7500 real-time PCR machine from Applied Biosystems (Foster City, CA) by using SYBR Green supermix (Roche, Indianapolis, IN). Results were calculated using Ct values and normalized to 36B4 mRNA level. Lipid and lipoprotein analysis. 7 Approximately 100 mg liver was homogenized in methanol and lipids were extracted in chloroform/methanol (2:1 v/v) as described [61]. Hepatic triglyceride and cholesterol levels were then quantified using Infinity reagents from Thermo Scientific (Waltham, MA). Hepatic fatty acid profile was quantified using gas chromatography (GC)-mass spectrometry at the Mouse Metabolic Phenotyping Center (MMPC) of Case Western Reserve University (Cleveland, OH). Hepatic total free fatty acids and free cholesterol were quantified using kits from BioVision (Milpitas, CA). Plasma lipid and glucose levels were also determined using Infinity reagents. Briefly, after 100 mL plasma was injected, lipoproteins were run at 0.5 mL/min in a buffer containing 0.15 M NaCl, 0.01 M Na2HPO4, 0.1 mM EDTA, pH 7.5, and separated on a Superose 6 10/300 GL column (GE Healthcare) using the BioLogic DuoFlow QuadTec 10 System (Bio-Rad, Hercules, CA). A 500-mL sample per fraction was collected. Adenovirus. Ad-Ces1-GFP was constructed by cloning mouse Ces1 cDNA into pAd-shuttleIRES-hrGFP vector (Stratagene, CA) as described previously [62]. To generate adenovirus expressing small hairpin RNA against Ces1 (Ad-shCes1), oligonucleotides were designed using BLOCK-iT™ RNAi Designer (Invitrogen, CA), annealed, and ligated to pEnter/U6 vector (Invitrogen, CA). Adenovirus was then generated following the instructions provided by Invitrogen. Three different shRNA oligonucleotides against murine Ces1 were designed. The sequences that produced the most inhibitory effect on endogenous Ces1 expression 8 are: 5’- GCTGATTCCAGCAGCTATTGACGAATCAATAGCTGCTGGAATCAGC-3’ (top strand) and 5’-GCTGATTCCAGCAGCTATTGATTCGTCAATAGCTGCTGGAATCA GC-3’ (bottom strand). Adenoviruses expressing ChREBP and shAcl have been described previously [63,64]. All the adenoviruses were grown in 293A cells and purified by cesium chloride gradient centrifugation. About 1-2x109 plaque formation units (pfu) of adenoviruses were transfused into each mouse intravenously. Unless otherwise stated, 7 days post infection, mice were fasted for 5-6 h and then euthanized. Primary hepatocyte isolation. Mouse primary hepatocytes were isolated as described [65,66]. Mice were anaesthetized by intraperitoneal injection of 50 mg/kg pentobarbital. The portal vein was cannulated with a 23-gauge plastic cannula. Mouse livers were perfused with Hank’s Balanced Salt Solution (HBSS, cat#14170-112, Thermo Fisher Scientific) with 0.19g/L EDTA. Simultaneously, the inferior vena cava was cut open. Subsequently, livers were perfused with HBSS, calcium, magnesium buffer (cat#14025092, Thermo Fisher Scientific) with 0.8mg/mL Collagenase from Clostridium histolyticum type IV (Sigma, St. Louis, MO). Primary hepatocytes were released and collected in a 50 mL centrifuge tube. After centrifugation at 50g for 5 minutes and washing with DMEM, cells were cultured in 6-well plate pre-coated with 0.1% gelatin in a 2 mL of DMEM+10% FBS. Mutagenesis and Transient Transfection Assays. The mutant pGL3 promoter-luciferase construct was generated using a QuickChange Site-directed Mutagenesis kit from Agilent (Santa Clara, CA). HepG2 cells 9 were plated in a 24-well plate and cultured in DMEM containing 10% FBS. Transient transfections were performed in triplicate as described [67]. Briefly, pGL3-Ces1 luciferase reporter constructs were transfected into HepG2 cells together with plasmids expressing FXR or RXR, followed by treatment with either vehicle or GW4064 (1 M). After 36 h, luciferase activities were determined and normalized to -galactosidase activity. For glucose-induced CES1 expression, pGL3-Ces1 luciferase reporter constructs were transfected into HepG2 cells, followed by treatment with either 5.5 mM glucose or 27.5 mM glucose. To determine the effect of over-expression of CES1 on PPAR activity, a CES1-expression plasmid was co-transfected with the 3xPPRE-luc plasmid, and luciferase activity was determined as described above. ChIP assay: Chromatin immunoprecipitation (ChIP) assays were performed as described previously [68] and following the manufacturer’s instructions (Millipore, Bilerica, MA). 200 mg liver for each sample was used for ChIP assays. Antibodies against acetyl-H3K9, acetyl-H4 and FXR (Cell Signaling Technology, Danvers, MA) were used to immunoprecipitate chromatin. Non-immune IgG was used as a measure of nonspecific background in immunoprecipitation. Chromatin purified from 10% sonicated tissue lysate was used as “input”. Real-time PCR was performed to test the chromatin enrichment on CES1 promoter region. ChIP assays were performed in triplicates. Electrophoretic Mobility Shift Assay (EMSA). 10 Oligonucleotides containing the putative FXR response element (FXRE; DR-5; 218 bp) were annealed, and EMSA and competition studies were performed as previously described [68]. The wild-type oligonucleotide sequence used for EMSA was ATG TAA GAT GTT CCT TGG TTA GTT TAT GGA CCT CTG TTA TCT GAG AGC TGT CCA ATG G (top strand). The mutant oligonucleotide sequence was ATG TAA GAT GTT CCT TAA TTA GTT TAT GCA AAT CTG TTA TCT GAG AGC TGT CCA ATG G (top strand) (mutation sites are underlined). Western Blot Assay. Western blot assays were performed using whole liver lysates [62] or nuclear lysates of the liver samples as described previously.[69] CES1 antibody was purchased from Abcam (Cambridge, MA, USA). -actin antibody was from Novus Biologicals (Littleton, CO). p-AKT(ser473) and AKT were from Millipore (Billerica, MA). SREBP-1 antibody was from Novus Biologicals (Littleton, CO). SREBP-2 antibody was from Cayman Chemicals (Ann Arbor, MI). Histone antibody was from Cell Signaling (Beverly, MA). VLDL Secretion. C57BL/6J mice were injected intravenously with specific adenoviruses. On day 6, these mice were fasted overnight, followed by intravenous injection of Tyloxapol (500 mg/kg). Blood was taken at indicated time points and plasma TG levels were determined. VLDL secretion rate was determined as described [70]. 11 Hepatic Lipogenesis. Mice were fasted for 4 h and then injected intraperitoneally with 2H2O (20-30 l/g). After 4 h, liver and plasma were snap-frozen in liquid nitrogen. The newly synthesized palmitate, triglycerides, and cholesterol were measured by mass spectrometry at MMPC of Case Western Reserve University. TGH Activity Assay. Cells were lysed in lysis buffer containing 50mM Tris, pH 7.4, 0.25M Sucrose, and 1mM EDTA, 1 mM dithiothreitol, 50 mM NaF and protease inhibitor cocktail (Sigma). After spun at 14,000 x g, the supernatant was used for TGH activity assay. For animals, liver was harvested after a 6-h fast. Liver was homogenized in lysis buffer and then spun at 800 x g to remove cell debris. The supernatant was then centrifuged at 100,000 × g for 1 h. The cytosolic portion and microsome portion were used for TGH assay separately. In brief, 100 l liver extracts (100 μg of protein) were incubated at 37 oC with 100 l substrates containing 0.15 mM cold triolein, 0.32 M [3H]triolein, 10 M egg yolk lecithin, 100 M sodium taurocholate, 1 mM dithiothreitol, and 50 mM potassium phosphate (pH 7.4). After 1 h, the reaction was stopped by 3.75 ml methanol:chloroform:heptane (10:9:7) and 1 ml of 0.1 M potassium carbonate/0.1M boric acid. After centrifuge at 800 x g, 1 ml top phase is used for counting radioactivity using a liquid scintillation counter. Fatty Acid Oxidation. 12 AML12 cells were cultured in DMEM containing 10% FBS in 12-well dishes and infected with either Ad-GFP or Ad-Ces1. After 48 h, the media were removed and washed with 1XPBS. The cells were then cultured in DMEM containing 0.5% fatty acidfree BSA, 0.5 Ci [3H]palmitate and 500 M cold palmitate. Fatty acid oxidation was performed as described [71]. Briefly, after incubation for 3 h, the supernatant was collected. 0.1 ml supernatant was added to a round-bottomed Eppendorf tube containing 0.9 ml of charcoal slurry. The samples were left at room temperature for 30 min with intermittent shaking (at least once every 5 minutes), and then centrifuged at 13,000 rpm for 15 minutes. 0.2 ml of the supernatant was carefully taken out and added to a scintillation vial containing 2.8 ml of scintillation liquid and the radioactivity was determined on a liquid scintillation counter. Statistical Method: The data were analyzed statistically using unpaired Student’s t-test (two-tailed) and ANOVA (for more than two groups), followed by a post hoc Newman-Keuls test. The data were expressed as mean±SE. Only p<0.05 was considered statistically significant. 13 1.3 RESULTS 1.3.1 Hepatic Carboxylesterase 1 is Induced by Glucose and Regulates Postprandial Glucose Levels. Hepatic CES1 is regulated by nutritional status. To determine whether nutritional status affects CES1 expression, we first determined hepatic CES1 expression in diabetic mice. Our data indicated that hepatic Ces1 mRNA (Figure 1A, B) and protein (Figure 1C) levels were significantly induced in both type 2 diabetic ob/ob mice and db/db mice. In streptozotocin (STZ)-treated mice, a type 1 diabetic mouse model, hepatic Ces1 mRNA levels were induced by >7 fold (Figure 1D). Peroxisome proliferator-activated receptor gamma coactivator-1 (PGC-1) and phosphoenolpyruvate carboxykinase (PEPCK) served as positive controls. To investigate whether hepatic CES1 expression is affected by a Western diet feeding, we fed C57BL/6 mice a high fat/high cholesterol (HFHC) diet; the data show that HFHC diet feeding did not change hepatic Ces1 expression (Figure 1E). ATP-binding cassette (ABC) transporter A1 (ABCA1) and ABC transporter G5 (ABCG5) served as positive controls. Finally, we investigated the effect of fasting on hepatic CES1 expression. The data of Figure 1F show that fasting for 8 or 24 hours caused a reduction in hepatic CES1 protein levels. Overall, these data indicated that hepatic CES1 is regulated by nutritional status and glucose may induce hepatic CES1 expression. 14 Figure 1. Hepatic CES1 is regulated by nutritional status. (A-C) Hepatic mRNA levels in ob/ob (A) and db/db mice (B) mice were determined by qRT-PCR and protein levels determined by Western blot assays (C) (n=4-6 mice per group). (D) C57BL/6 mice were treated with either vehicle (0.1 M sodium citrate, pH 4.5) or streptozotocin (STZ) (50 mg/kg/d) for 5 days. Seven days after STZ treatment, mice were euthanized and 15 hepatic mRNA levels were quantified (n=5 mice per group). (E) Wild-type mice were fed a chow or high fat/high cholesterol (HFHC) diet (21% fat, 1.5% cholesterol) for 3 weeks and hepatic mRNA levels were determined (n=8 mice per group). (F) C57BL/6 mice were fed a chow diet, or fasted for 3, 8, 24 h, or fasted for 24 h followed by refed for 24 h (n=5 mice per group). Hepatic protein levels were determined. Pgc-1, peroxisome proliferator-activated receptor gamma coactivator-1. Abca1, ATP-binding (ABC) transporter A1. Abcg5, ABC transporter G5. carboxykinase. *p<0.05, **p<0.01 16 cassette Pepck, phosphoenolpyruvate Hepatic CES1 is regulated by glucose but not insulin To test our hypothesis that glucose induces hepatic CES1 expression, saline or glucose (8g/kg) were administered to C57BL/6 mice twice with 3 hours interval via oral gavage. Mice were sacrificed 3 hours after second oral gavage and hepatic CES1 expression was determined. Hepatic Ces1 mRNA level (Figure 2A) and protein levels (Figure 2B, C) were induced by ~2 fold in response to glucose stimulation. In contrast, insulin did not induce hepatic CES1 expression (Figure 2D). In mouse primary hepatocytes, high glucose (27.5 mM) induced hepatic Ces1 mRNA expression (Figure 2E), suggesting that glucose can directly regulate CES1 expression. To test whether glucose can directly regulate Ces1 promoter activity, Transient transfection assays and luciferase reporter assays were performed using a serial of luciferase reporter constructs with 5’-deletions. The data show that glucose stimulated Ces1 promoter activity through a region between 75 bp and 150 bp upstream of the transcription start site (Figure 2F). Collectively, the data of Figure 2 demonstrate that glucose induces CES1 expression both in vivo and in vitro. 17 Figure 2. Hepatic CES1 is regulated by glucose but not insulin. (A-C) C57BL/6 mice were fasted for 16 h, followed by gavage with saline or glucose (8g/kg) (n=6 mice per group). Hepatic mRNA levels (A) and protein levels (B) were determined. Hepatic CES1 protein levels were quantified (C). L-pk serves as a positive control in (A). (D) C57BL/6 mice were fasted for 16 h, followed by i.p. injection of either saline or insulin (0.8 18 units/kg) (n=5 mice per group). After 3 h, mice were euthanized. Srebp-1c serves as a positive control. (E) Mouse primary hepatocytes were isolated and cultured in dulbecco’s modified eagle medium (DMEM) plus 10% fetal bovine serum (FBS) overnight, followed by serum-free fasting for 8 h. Cells were then treated with either normal (5.5 mM) or high (27.5 mM) glucose for additional 24 h prior to quantification of mRNA levels. Fas serves as a positive control. (F) CES1 promoter-luciferase constructs were transfected into HepG2 cells, then treated with 5.5 mM or 27.5 mM glucose. After 36 h, luciferase activity was determined. Srebp-1c, sterol response element binding protein-1c. L-pk, liver type pyruvate kinase. Fas, fatty acid synthase. RLU, relative luciferase units. *p<0.05, **p<0.01 19 ACL is required for glucose-induced hepatic CES1 expression ChREBP is suggested to be the principal mediator of glucose metabolism in the liver [72,73,74]. Absence of ChREBP leads to the failure of glucose to induce hepatic glycolytic gene (L-PK) and lipogenic genes (ACC and FAS) [75]. However, overexpression of ChREBP in the liver had no effect on hepatic Ces1 expression (Figure 3A), suggesting that glucose induces hepatic CES1 expression independent of ChREBP. It has been shown that ACL is required for glucose-mediated histone acetylation and gene activation [76,77]. Thus, we investigated the role of ACL in glucose-mediated hepatic CES1 expression. Adenovirus-mediated expression of Acl shRNA reduced hepatic Acl mRNA levels by >85% (Fig. 3B). Interestingly, glucose induced hepatic CES1 mRNA and protein expression in the control mice but not in Acl-deficient mice (Figure 3C-E). Liver-type pyruvate kinase (L-PK), a gene responsive to glucose stimulation, served as a positive control (Figure 3F). Hence, the data of Figure 3 indicate that ACL is required for glucose to induce hepatic CES1 expression. 20 Figure 3. ACL is required for glucose-induced hepatic CES1 expression. (A) C57BL/6 mice were injected i.v. with adenovirus expressing GFP or ChREBP. After 5 days, hepatic mRNA levels were determined by qPCR (n=7 mice per group). (B-F) C57BL/6 mice were injected with adenovirus expressing shLacZ or shAcl (n=6 mice per group). After 5 days, mice were gavaged with either saline or glucose (8g/kg). Hepatic mRNA levels of Acl (B), Ces1 (C) and L-pk (F) were determined. Hepatic protein levels 21 were determined by Western blot assays (D) and CES1 protein levels quantified (E). *p<0.05, **p<0.01 ACL is required for glucose-mediated acetylation of histones (H3, H4) in the Ces1 chromatin Several lines of evidence have shown that glucose may regulate gene expression via epigenetic modifications [76,78,79,80]. ACL converts glucose-derived citrate to acetyl-CoA, which subsequently serves as substrate for histone acetyltransferase for acetylation of H3 and H4 tails. Our data showed that glucose increased the acetylation of histone 3 and histone 4 in the Ces1 chromatin and these effects were abolished in Acldeficient mice (Figure 4A, B), indicating that ACL is required for glucose-mediated acetylation of histones (H3 and H4) in the Ces1 chromatin. Figure 4. ACL is required for glucose-mediated acetylation of histones (H3, H4) in the CES1 chromatin. (A, B) C57BL/6 mice were treated with glucose as described in Fig. 3. Liver lysates were used for ChIP assay to determine acetylation of histone 3 (AcH3) (A) and histone 4 (AcH4) (B). *p<0.05, **p<0.01 22 CES1 regulates postprandial glucose levels and insulin sensitivity Postprandial blood glucose levels are tightly controlled to avoid any unwanted side effect of glucose. The finding that glucose induces CES1 expression and that CES1 regulates glucose metabolism [1] suggest that hepatic CES1 may regulate postprandial blood glucose levels. To test this hypothesis, C57BL/6 mice were injected with AdshLacZ and Ad-shCes1. Five days after adenovirus injection, mice were fasted for 16 h prior to gavage with saline or glucose (8 g/kg). Blood glucose levels were measured 1 h after gavage. For saline treatment, Ces1-deficient mice had similar blood glucose levels compared with the control mice (Figure 5A). For glucose treatment, however, Ces1deficient mice had significantly higher blood glucose levels compared to the control mice (Figure 5A). In the liver, expression of Ces1 shRNA resulted in elevated levels of hepatic triglycerides (TG) (Figure 5B) and free fatty acids (FFAs) (Figure 5C). Consistent with the latter data, hepatic Ces1 knockdown reduced the ratio of phospho-AKT (p-AKT) to total AKT (Figure 5D and 5E). Finally, knockdown of hepatic Ces1 reduced hepatic Ces1 mRNA levels by>90% (Figure. 5F), and increased hepatic PEPCK and glucose 6phosphatase (G6Pase) expression in saline- but not glucose-treated mice (Figure 5G and 5H). These latter data suggest that hepatic CES1 deficiency may cause hepatic insulin resistance and that the increase in postprandial glucose levels may not be a result of uncontrolled hepatic glucose production. Together, the data of Figure 5 demonstrate that hepatic CES1 plays an important role in regulating postprandial glucose levels. 23 Figure 5. CES1 regulates postprandial levels. (A–H) C57BL/6 mice were injected with Ad-shLacZ or Ad-shCes1. After 5 days, mice were fasted for 16 h followed by gavage with saline or glucose (8 g/kg) (n = 6 mice per group). Blood glucose levels were measured 1 h after gavage using a glucometer (A). Mice were then sacrificed 3 hours after gavage. Hepatic triglyceride (TG) (B) and free fatty acid (FFA) (C) levels were analyzed. Hepatic protein levels were assessed by Western blot assays (D) and then the ratio of p-AKT to total AKT was quantified (E). Hepatic mRNA levels of Ces1 (F), PEPCK (G) and G6Pase (H) were determined by qRT-PCR. (I) Reciprocal regulation between plasma glucose and hepatic CES1. Elevated plasma glucose induces hepatic CES1, which in turn helps lower plasma glucose levels likely via increasing peripheral insulin sensitivity. AKT, protein kinase B. *p<0.05 **p<0.01. 24 1.3.2 Hepatic Carboxylesterase 1 is Essential for Normal and Farnesoid X ReceptorControlled Lipid Homeostasis. Over-expression of hepatic CES1 reduces hepatic triglyceride and improves glucose homeostasis. We initially examined whether hepatic CES1 plays a role in regulation of hepatic lipid homeostasis. Adenovirus-mediated over-expression of hepatic Ces1 (Ad-Ces1-GFP) had no effect on plasma TG and TC levels (data not shown), but significantly lowered plasma glucose and hepatic TG levels with reduction of ~30% and 60%, respectively (Figure 6A). Consistently, over-expression of hepatic Ces1 in ob/ob mice reduced plasma glucose and hepatic TG levels (Figure 6B). H&E and Oil red O staining revealed that over-expression of hepatic Ces1 reduced lipid accumulation in liver, which is also evidenced by the appearance of more reddish liver (Figure 6C). The lowered glucose level led us to determine the role of hepatic CES1 in glucose metabolism. Hepatic expression of Ces1 improved glucose tolerance in glucose tolerance test (Figure 6D). In addition, p-AKT/AKT ratio was markedly elevated, suggesting that hepatic Ces1 enhances insulin signaling (Figure 6E). Collectively, over-expression of hepatic Ces1 reduces hepatic lipid accumulation and improves glucose homeostasis. 25 26 Figure 6. Hepatic expression of CES1 lowers hepatic triglyceride levels and improves glucose homeostasis. (A) C57BL/6 mice were i.v. injected with either AdGFP or Ad-Ces1 (n=7–8 mice per group). After 7 days, mice were fasted for 5 h. Plasma glucose (left panel) and hepatic TG (right panel) levels were determined. (B–F) ob/ob mice were i.v. injected with Ad-GFP or Ad-Ces1 (n=5 mice per group). After 7 days and a 5-h fast, mice were euthanized. Plasma glucose (B, left panel) and hepatic TG levels (B, right panel) were determined. Representative liver images are shown in (C, top panel) and representative H&E staining (C, middle panel) or oil red O staining (C, bottom panel) of the liver sections are shown in (C). Glucose tolerance test (GTT) was performed after a 16 h fast (D). Western blot assays were performed using liver lystates (E) and protein levels quantified using Image J software (F). 27 Over-expression of hepatic CES1 does not affect fatty acid lipogenesis or very lowdensity lipoprotein (VLDL) secretion. To determine the mechanism by which CES1 reduces lipid accumulation in the liver, genes involved in cholesterol, fatty acids and TG synthesis, and fatty acid oxidation (FAO) were analyzed. Figure 7A showed that mRNA levels of genes in cholesterol synthesis (Srebp-2, HMG-CoA reductase (Hmgcr) and HMG-CoA synthase (Hmgcs)) were reduced; genes controlling fatty acid and TG synthesis (Srebp-1c, acetyl-CoA carboxylesterase 1 (Acc1), fatty acid synthase (Fas), acyl-CoA: diacylglycerol acyltransferase 1 (Dgat1) and Dgat2) were not significantly changed, except that mRNA level of Acc2, a gene which represses fatty acid oxidation (FAO) was decreased; Ppar target genes (fatty acid translocase (Cd36), pyruvate dehydrogenase kinase 4 (Pdk4), angiopoietin-like protein 4 (Angptl4), carnitine palmitoyltransferase 1b (Cpt1b)) were markedly increased. Interestingly, genes involved in gluconeogensis (phosphoenolpyruvate carboxykinase (Pepck) and glucose-6-phosphate (Gpase6) were not significantly changed in wild-type mice injected with Ad-Ces1 (Figure 7A left panel), but significantly reduced in ob/ob mice injected with Ad-Ces1 (Figure 7A right panel). This result is in agreement with enhanced insulin signaling in ob/ob mice injected with Ad-Ces1 (Figure 6E). Glucokinase (Gck), which facilitates glucose uptake by phosphorylating glucose to glucose-6-phosphate, is transcriptionally regulated by insulin [81]. Hepatic expression of Ces1 increased Gck mRNA level by >2 fold (Figure 7A right panel). Next, we examined de novo lipogenesis by injection of mice with 2H2O. Consistent with the gene expression data (Figure 7A), over-expression of hepatic Ces1 28 did not significantly change hepatic de novo biosynthesis of palmitate (Figure 7B) or TG (Figure 7C), but reduced hepatic cholesterol biosynthesis (Figure 7D). It has been shown that Ces3 is involved in very low-density lipoprotein (VLDL) assembly in the liver and affects plasma TG and CHOL levels [54]. Accordingly, we determined whether Ces1 had similar effect. Interestingly, over-expression of hepatic Ces1 had no effect on the protein levels of microsomal triglyceride transfer protein (MTP) or apolipoprotein B (ApoB) (Figure 7E), or VLDL secretion rate (Figure 7F). This result is in line with unchanged plasma lipid levels in hepatic expression of Ces1 mice. Thus, hepatic expression of CES1 did not affect TG biosynthesis or VLDL secretion. 29 Figure 7. Hepatic expression of CES1 selectively regulates gene expression and has no effect on lipogenesis or VLDL secretion. (A) Hepatic mRNA levels in wild-type (left panel) or ob/ob mice (right panel) were determined by qRT-PCR. (B–D) De novo lipogenesis was determined in mice after injection of 2H2O (n=5 mice per group). The levels of newly synthesized [2H]palmitate (B), [2H]TG (C) or [2H]cholesterol (D) in the liver were quantified. (E) Hepatic protein level was assessed by Western blot assays. (F) VLDL secretion rate was determined (n=6 mice per group). 30 Over-expression of hepatic CES1 increases hepatic TG hydrolysis and stimulates fatty acid oxidation. The increased PPAR target gene expressions indicated that Ces1 overexpression may induce PPAR activity. To test this hypothesis, we co-transfected a Ces1-expressing plasmid and a luciferase-reporter plasmid containing 3 copies of PPAR responsive element (PPRE). PPAR activity was markedly induced by over-expression of CES1 (Figure 8A top penal). Free fatty acids (FFAs) are endogenous ligands for PPAR [82]. The increased PPAR activity is indicative of increased FFA level as a result of TG hydrolysis (Figure 8A bottom penal). This result suggests that CES1 has TG hydrolysis activity. In addition, over-expression of Ces1 significantly increased the release of [3H]FFAs from [3H]triolein in both COS-7 cells (Figure 8B) and liver (Figure 8C), indicating that CES1 is capable of hydrolyzing TG. Further study showed that hepatic total fatty acids and a number of long-chain fatty acids, such as C14:0, C16:0, C16:1, and C18:2, were significantly reduced by hepatic expression of Ces1 (Figure 8D). Hepatic FFA level was also reduced by ~20% (Figure 8E). The repression of Acc2 and induction of Cd36, Pdk4, Angptl4, and Cpt1b, all of which are involved in FAO, suggest that CES1 may regulate FAO. Indeed, overexpression of Ces1 increased FAO (Figure 8F). Thus, hepatic expression of Ces1 induces triglyceride hydrolysis and increases FAO, leading to reduced hepatic triglyceride level. 31 Figure 8. Hepatic expression of CES1 increases triglyceride hydrolase activity and activates PPARα (A) HepG2 cells were transfected with a control plasmid or a Ces1expressing plasmid together with a 3xPPRE-Luc reporter plasmid. Luciferase activity was determined (top panel). In the bottom panel, the diagram shows that CES1 hydrolyzes TG and releases FFAs, which bind to PPARα/RXR complex and then induce PPARα activity. (B, C) TGH activity was assessed using lysates from COS-7 cells (B) or the liver (C). (D, E) Hepatic fatty acid profile was determined by GC-mass spectrometry 32 (D) and hepatic FFA levels were quantified (n=8 mice per group) (E). (F) FAO was performed in the liver cell line AML12 cells that were infected with Ad-GFP or AdCES1 for 48 h, or treated with either vehicle or carnitine (1 mM) (n=3–5 per group). Carnitine treatment serves as a positive control. Veh, vehicle. 33 Knockdown of hepatic CES1 causes hepatic steatosis and elevated plasma cholesterol level. To determine whether hepatic CES1 is required for maintaining normal lipid and glucose homeostasis, we injected C57BL/6J mice with Ad-shLacZ (control) and AdshCes1. Hepatic Ces1 mRNA level was substantially reduced (Figure 9A). Its protein levels were also significantly decreased (Figure 9B). Despite the plasma TG level was not significantly changed, plasma TC level was increased by ~2 folds in mice injected with shCes1 (Figure 9C). In agreement with increased plasma TC level, hepatic Ces1 deficiency drastically increased VLDL-cholesterol and LDL-cholesterol levels and slightly reduced HDL-cholesterol level (Figure 9D). The liver of mice injected with shCes1 showed paler color (Figure 9E, top panel). Oil red O staining confirmed that knockdown of hepatic Ces1 led to increased accumulation of lipid droplets (Figure 9E, bottom panel). The latter finding suggests that hepatic Ces1 deficiency results in the development of fatty liver. Indeed, Hepatic TG level was increased more than 2 folds, whereas hepatic TC level was unchanged (Figure 9F). Collectively, knockdown of Ces1 results in fatty liver. 34 Figure 9. Loss of hepatic CES1 causes fatty liver and increased plasma cholesterol level (A–F) C57BL/6 mice were i.v. injected with either Ad-shLacZ or Ad-shCes1 (n=8 mice per group). After a 6-h fast, mice were euthanized. Hepatic mRNA (A) and protein (B) levels were determined. Plasma TG and total cholesterol (TC) levels were quantified (C). Plasma cholesterol lipoprotein profile was determined by FPLC (D). Representative liver images (top panel) and oil red O staining of liver sections (lower panel) are shown (E). Hepatic TG and TC levels were quantified (F). 35 Hepatic CES1 deficiency results in increased lipogenesis. Next, we determined whether the increased hepatic TG is caused by increased lipogenesis. Analysis of hepatic gene expression indicated that many SREBP- or cholesterol/LXR(liver X receptor)-regulated genes, including Hmgcs, Acc-1, Dgat1, Dgat2, ATP citrate lyase (Acl), Abca1, and Cd36, were significantly induced (Figure 10A). In agreement with the induction of a number of lipogenic genes, hepatic de novo lipogenesis of palmitate (Figure 10B), TG (Figure 10C), and cholesterol (Figure 10D), were induced by 2.1, 2.9 and 2.1 fold, respectively. Consistent with increased mRNA level of Srebp1c and Srebp2, the protein levels of hepatic mature/nuclear SREBPs (nSREBPs) were increased in mice injected with shCes1 (Figure 10E), suggesting that loss of Ces1 results in elevated SREBPs translocation from cytosol to nucleus. SREBP processing is known to be regulated by the change in intracellular sterol levels and is sensitive to cholesterol ester (CE) to free cholesterol (FC) ratio; when cellular sterol levels are low or CE/FC ratio are high, mature SREBPs are increased by inducing SREBP processing [83]. Given that CES1 has CEH property, we hypothesized that loss of Ces1 results in a reduction of cholesterol ester hydrolysis and leads to a subsequent reduction of free cholesterol level, which in turn induces SREBP processing. As predicted, hepatic FC level was reduced in Ces1-deficient mice (data not shown). The CE/FC ratio was increased. Thus, loss of hepatic Ces1 increased lipogenesis through increasing SREBPs processing, which may be accounted, at least in part, by reduced FC levels. 36 Figure 10. Loss of hepatic CES1 induces de novo lipogenesis (A) Hepatic mRNA levels were determined by qRT-PCR (n=8 per group). (B–D) De novo lipogenesis was determined in mice after injection of 2H2O (n=5 per group). The levels of newly synthesized [2H]palmitate (B), [2H]TG (C), or [2H]cholesterol (D) in the liver were quantified. (E) Hepatic protein levels were determined by Western blot assays (E, left panel) and then quantified by using Image J software (E, right panel). nBP-1, nuclear 37 form SREBP-1. nBP-2, nuclear form SREBP-2. (F) Hepatic ratio of CE to FC was determined (n=8 mice per group). The CE/FC ratio in shLacZ-treated mice was set at 1. 38 Hepatic CES1 is regulated by FXR. Activation of FXR is shown to be beneficial in regulating lipid and carbohydrate metabolism. We found that activation of FXR by GW4064, a FXR agonist, reduced hepatic TG level but did not change TC level (Figure 11A). Activation of FXR induced hepatic mRNA levels of Shp, an FXR target, and Ces1, but mRNA levels of Ces2 and Ces3 were unchanged (Figure 11B). Figure 11 C showed that GW4064 treatment increased protein levels of hepatic CES1, but did not change ApoB and MTP protein levels, suggesting that activation of FXR does not change VLDL secretion. In addition, mice were treated with obeticholic acid (OCA, INT-747), a potent and selective FXR agonist, or cholic acid, the endogenous FXR ligand, hepatic Ces1 mRNA was also induced (Figure 11D and E). On the other hand, hepatic Ces1 mRNA level was reduced in Fxr mice (Figure 11F). Therefore, hepatic CES1 expression is regulated by FXR. 39 Figure 11. Hepatic CES1 is regulated by FXR (A–C) C57BL/6 mice were gavaged with vehicle (0.5% CMC (carboxymethyl cellulose)) or GW4064 (30 mg/kg, twice a day) for 7 days (n=8 mice per group). Hepatic TG and TC levels were determined (A). Hepatic mRNA levels were quantified by qRT-PCR (B) and hepatic protein levels were determined by Western blot assays (C). (D) C57BL/6 mice were gavaged with either 40 0.5% CMC (vehicle) or INT-747 (OCA, 30 mg/kg/d) for 7 days (n=5 mice per group). Hepatic mRNA levels were determined. Cyp7a1 and Cyp8b1 serve as positive controls. (E) C57BL/6 mice were fed a chow diet or 0.5% cholic acid (CA) for 7 days. Hepatic mRNA levels were quantified. (F) Hepatic mRNA levels were quantified by qRT-PCR in wild-type or Fxr−/− mice (n=8 mice per group). 41 CES1 is a direct FXR target. Next, we determined how FXR regulated CES1 expression. A luciferase assay was performed using a serial 5’-deletion construct along with Fxr expressing plasmid. Figure 12A showed that activation of FXR increased Ces1 promoter activity. Luciferase and mutagenesis assays revealed that the FXR response element was located in Ces1 promoter region between 300bp and 150bp upstream of transcriptional start site (Figure 12A and B). Electrophoretic mobility shift assays showed that the FXR/RXR complex bound to this FXRE in vitro and this binding was competed away by cold wild-type oligonucleotides but not by mutant oligonucleotides (Figure 12C). Lastly, chromatin immunoprecipitation assay showed that FXR bound to the promoter of Ces1 in the liver. Akr1b7, a known FXR target gene [84], serves as a positive control (Figure 12 D). Overall, Figure 11 and Figure 12 suggested that Ces1 is regulated by FXR and its direct target gene. 42 Figure 12. CES1 is a direct FXR target gene (A, B) Transient transfection assays were performed using promoter-luciferase constructs containing a serial of 5'-deletions (A) or mutations (B). (C) EMSA assays were performed using in vitro transcribed/translated proteins. Wild-type (WT) and mutant (MUT) oligos were used in the competition assays. (D) Chromatin immunoprecipitation (ChIP) assays were performed using liver lysates (n=3 per group). Akr1b7 serves as a positive control. 43 Hepatic CES1 is essential for activated FXR to improve lipid homeostasis. Activation of FXR induced the expression of Ces1. This finding prompted us to study whether hepatic Ces1 mediated FXR-controlled lipid homeostasis. C57BL/6 mice were injected i.v. with either Ad-shLacZ or Ad-shCes1, followed by treatment with either vehicle or OCA for 7 days. As expected, knockdown of hepatic Ces1 increased plasma cholesterol level by ~4 fold (Figure 13A). Interestingly, OCA treatment reduced plasma cholesterol levels in the control mice but not in Ces1-deficient mice (Figure 13A). Similarly, OCA treatment also reduced plasma and hepatic TG levels in the control mice but not in the Ces1-deficient mice (Figure 13B and C). This result suggested that hepatic Ces1 is important in FXR-controlled lipid homeostasis. 44 Figure 13. Essential roles of hepatic CES1 in FXR-regulated lipid homeostasis (A–C) C57BL/6 mice were i.v. injected with Ad-shLacZ or Ad-shCes1. The next day, these mice were gavaged with either vehicle (0.5% CMC) or INT-747 (OCA, 30 mg/kg/d) for 7 days (n=8–10 mice per group). After a 5-h fast, mice were euthanized. Plasma total cholesterol (TC) (A) and TG (B) levels as well as hepatic TG levels (C) were determined. 45 CES1 deficiency alters the lipid profile in mice treated with FXR agonist Activation of FXR tended to reduce plasma glucose level in control mice (shLacZ). Similarly, plasma glucose level was reduced in OCA treated Ces1-deficient mice (Figure 14A). Analysis of plasma by FPLC showed that loss of hepatic Ces1 markedly increased plasma non-HDL-C levels (Figure 14B) and also slightly increased LDL triglyceride levels (Figure 14C), and these changes were exacerbated following OCA treatment (Figure 14B and C). Thus, hepatic CES1 is critical for an FXR agonist to lower plasma TG and cholesterol levels. In addition, OCA did not significantly change hepatic cholesterol level, whereas it increased cholesterol level in Ces1-deficient mice (Figure 14D). 46 Figure S7 Figure S7 A A B B C C D D Figure 14. Effects of the FXR agonist OCA (INT-747) on lipid and glucose homeostasis in C57BL/6 mice. C57BL/6 mice were injected i.v. with either Ad-shLacZ or Ad-shCes1. On the next day, these mice were gavaged with either vehicle or OCA (n=7-9 mice per group). After 7 days, mice were fasted for 5 h. Plasma glucose levels (A) and hepatic cholesterol levels (D) were measured. Plasma cholesterol (B) and triglyceride (C) lipoprotein profiles were determined by FPLC. 47 Ces1 is important in FXR-controlled lipid homeostasis in ob/ob mice. To further define the role of Ces1 in FXR-controlled lipid homeostasis, ob/ob mice were injected with Ad-shLacZ or Ad-shCes1, followed by OCA treatment for 7 days. Similar to what we observed in C57BL/6 mice, we found that OCA treatment lowered plasma cholesterol and TG levels as well as hepatic TG levels in a CES1dependent manner (Figure 15A, B and D), but had no effect on hepatic cholesterol levels (Figure 15C). Analysis of hepatic gene expression showed that OCA treatment induced hepatic mRNA levels of Ces1 in the control mice and Shp in both the control and Ces1deficient mice (Figure 15F). Collectively, hepatic CES1 is indispensable for activated FXR to regulate both plasma lipid and hepatic TG levels. 48 Figure S8 A B C D E F Figure 15. Effects of the FXR agonist OCA on lipid homeostasis in ob/ob mice. Ob/ob mice were injected i.v. with either Ad-shLacZ or Ad-shCes1. On the next day, these mice were gavaged with either vehicle or OCA (n=6 per group). After 7 days, mice were fasted for 5 h. Plasma total cholesterol (TC) (A), plasma triglycerides (TG) (B), hepatic TC (C) and hepatic TG (D) levels were determined. Hepatic mRNA levels of Ces1 (E) and Shp (F) were quantified by qRT-PCR. * p<0.05, ** p<0.01. NS, nonspecific. 49 1.4 DISCUSSION CES1 is highly expressed in the liver. The current studies show that CES1 has TGH activity; it reduced hepatic TG level via increasing TG hydrolysis and subsequent fatty acid oxidation (FAO) [52,85]. Lipid homeostasis has a profound impact on insulin sensitivity and glucose metabolism. Plasma free fatty acid (FFA) levels correlate negatively to the degree of insulin sensitivity [86,87]. Given the regulatory role of CES1 in TG hydrolysis and FAO, it is not surprising to see that increased hepatic CES1 expression lowers plasma glucose levels and improve insulin sensitivity. In the present study, we investigate the regulation of hepatic CES1 by glucose and the physiological role of such regulation. Our data reveal a novel role for hepatic CES1 in postprandial glucose control. CES1 regulates the postprandial glucose levels. Poor control of postprandial glucose is a significant contributor to type 2 diabetes mellitus. Persistent, moderate increase in postprandial glucose levels is a significant risk factor for macrovascular complications, and is more indicative of atherosclerosis than fasting glucose. In light of the risks of postprandial hyperglycemia for vascular events, tight control of postprandial glucose levels is important for long-term indices of diabetes control. After the start of a meal, blood glucose levels are increased. The increased blood glucose levels induce hepatic CES1 expression, which in turn helps lower blood glucose levels likely by increasing hepatic insulin sensitivity. This conclusion is supported by the 50 findings that increased hepatic CES1 expression lowers plasma glucose levels whereas loss of hepatic CES1 results in increased postprandial blood glucose levels. Although our data show that hepatic CES1 is required for regulating postprandial glucose levels, the underlying mechanism remains to be fully determined. Global Ces1mice have elevated plasma levels of triglyceride, free cholesterol, FFAs and insulin [60]. These mice also present insulin resistance, which results from reduced insulin sensitivity in both skeletal muscle and white adipose tissue [60]. However, CES1 is not expressed in skeletal muscle and its expression level in white adipose tissue is low (data not shown). Thus, insulin resistance observed in global Ces1mice likely results from a deficiency in hepatic Ces1. Consistent with the speculation, our data show that hepatic Ces1 deficiency results in impaired postprandial glucose clearance. Although hepatic CES1 deficiency may cause hepatic insulin resistance, skeletal muscle and white adipose tissues are the major organs responsible for plasma glucose clearance. Therefore, hepatic Ces1 deficiency may affect peripheral insulin sensitivity. To precisely understand how hepatic CES1 deficiency regulates glucose homeostasis or insulin sensitivity, heperinsulinemic-euglycemic clamp studies will be needed to help characterize the underlying mechanism. 51 CES1 is induced by glucose. We have previously shown that farnesoid X receptor regulates CES1 expression [1]. The present study showed that hepatic CES1 is also regulated under physiological and pathological conditions. Under these latter conditions, hepatic CES1 expression is altered likely due to the change in plasma glucose levels. Indeed, our data show that glucose induces CES1 expression both in vitro and in vivo. Consistent with our finding, very recent data by Xiong et al. also show that glucose induces the expression of CESs in mouse primary hepatocytes [88]. Several lines of evidence suggest that glucose regulates gene expression through epigenetic modifications [78] and that nuclear ACL is important for glucose-mediated histone acetylation [89]. In this study, we show that glucose induces CES1 expression by epigenetic modifications of the acetylation status (H3 and H4) of the CES1 chromatin in an ACL-dependent manner. The histone tails interact with a region about 150bp upstream of transcription start site of CES1. Consistent with this finding, the data from the luciferase-promoter assays show that this region is required for glucose to induce CES1 promoter activity. Thus, glucose induces CES1 expression via acetylation of H3 and H4 in the CES1 chromatin. 52 Recent reports show that global deletion of CES1 promotes hepatic steatosis, obesity and hyperlipidemia [60]; intestinal CES1 regulates chylomicron secretion [59]. The TG and CE hydrolase activity of CES1 have also been reported, which highlight its important role in regulating hepatic lipid homeostasis. In the present studies, we demonstrate that hepatic CES1 controls lipid and carbohydrate metabolism, reinforcing its values as a therapeutic target in treating NAFLD. Additionally, we find that FXR regulates hepatic CES1 expression, and the induction of hepatic CES1 is indispensable for activated FXR to improve lipid homeostasis. CES1 plays a key role in regulating hepatic triglyceride and cholesterol levels. We clearly demonstrated that hepatic CES1 has TG hydrolase activity, suggesting its role in maintaining hepatic TG homeostasis. Hepatic TG mobilization is little understood. Previous data suggest that ATGL may play an important role in mobilizing hepatic TG. However, ATGL expression is low in hepatocytes [90]. CES1, expressed abundantly in liver, may be one of major enzymes which control hepatic TG levels. The key finding of this study is that over-expression of hepatic CES1 increases TG hydrolysis, reduces hepatic TG levels and increases the release of FFA, which in turn activates PPAR and promotes FAO, leading to reduced FFA levels. The reduced FFA levels, in turn, lead to increased hepatic insulin sensitivity and reduced plasma glucose levels, suggesting that hepatic expression of Ces1 also improves carbohydrate metabolism. On the other hand, loss of hepatic CES1 significantly increases lipogenesis 53 and SREBPs processing. These results suggest that CES1 is required for maintaining normal lipid homeostasis. CES1 is important in FXR-controlled lipid homeostasis. FXR is emerging as a significant target for treatment of fatty liver disease [91]. However, the mechanism underlying FXR-mediated alleviation of hepatic steatosis remains undetermined. Our study provides compelling evidence demonstrating that CES1 is a direct downstream target of FXR and is necessary for activated FXR to lower plasma TG and cholesterol levels. Using ob/ob mice, we demonstrate that loss of hepatic CES1 abolishes the effect of activation of FXR on reducing hepatic steatosis, suggesting that hepatic expression of CES1 is essential for using FXR agonist in treating metabolic disease. Therefore, hepatic CES1 plays an essential role in FXR-controlled lipid homeostasis. In summary, in chapter 1, we proposed a glucose-CES1-glucose (Figure 5I) reciprocal cascade, in which glucose induces hepatic CES1 expression, which in turn increases insulin sensitivity and lowers blood glucose levels. This cascade plays an important role in regulating postprandial glucose levels. Since high levels of postprandial blood glucose contribute to macrovascular complications, CES1 may be targeted for prevention of vascular diseases associated with hyperglycemia. In addition to its role in regulation of carbohydrate metabolism, over-expression of hepatic CES1 is shown to have beneficial effects on lipid metabolism. In contrast, loss of hepatic CES1 causes fatty liver and proatherogenic lipid profile. Extensive studies demonstrate that CES1 is a 54 direct target of FXR and is critical for FXR-mediated improvement of lipid homeostasis. Together, our data suggest that hepatic CES1 is essential in normal and FXR-controlled lipid and carbohydrate homeostasis, and also suggest that hepatic CES1 may represent a therapeutic target for treatment of NAFLD. 55 CHAPTER 2. THE ROLE OF CARBOXYLESTERASE 1 IN ALCOHOLIC FATTY LIVER DISEASE 2.1 INTRODUCTION Alcoholic liver disease (ALD) is a major cause of chronic liver disease, ranging from simple steatosis, hepatitis, to more severe forms including fibrosis, cirrhosis and hepatocellular carcinoma (HCC). Liver cirrhosis is the 12th leading cause of death in the US, with a total of 36,427 deaths in 2013, 49.8% of which are related to alcohol [92]. Abstinence is an effective strategy for the treatment of ALD, but sustainable lifestyle changes are difficult for many patients to achieve. Pharmacological treatments, such as the use of alcohol dehydrogenase inhibitors and anti-craving drugs, do not achieve satisfactory effect in patients with ALD [92] Alcohol-induced liver damage is characterized by a reduction of NAD/NADH and subsequent inhibition of fatty acid oxidation, up-regulation of lipogenesis, and an increase in inflammation and oxidative stress which promote hepatocyte necrosis and apoptosis [93,94,95]. Alcohol also increases hepatic free fatty acid (FFA) uptake and decreases lipolysis, resulting in impaired very low density lipoprotein (VLDL) secretion [96]. Alcohol-induced liver injury is associated with enhanced lipid peroxidation, protein carbonyl formation, formation of free radicals, and decrease in glutathione [97]. 56 Enzymes, which govern the metabolism of alcohol, include alcohol dehydrogenase, acetaldehyde dehydrogenase and cytochrome P450 system, and abnormal regulation of these enzymes are associated with the development of ALD. Alcoholinduced hepatic steatosis is prominent in ALD. Alcohol disrupts fatty acid oxidation by down-regulating peroxisome proliferator-activated receptor (PPAR)-α, and damaging mitochondrial functions [98,99,100]. Chronic alcohol administration suppresses sirtuin 1 (SIRT1)-AMP-activated kinase (AMPK) axis [101,102,103,104], thus represses rates of fatty acid oxidation and enhances lipogenesis through modulating PPAR-γ coactivator-α (PGC-1α)/PPARα and sterol regulatory element-binding protein 1 (SREBP-1) [101,105,106]. Alcohol increases gut epithelial membrane permeability, thus increases translocation of lipopolysaccharides (LPS), which in turn elicits Toll-like receptor (TLR)mediated signaling cascade and subsequent increase in productions of cytokines (TNF-α, IL-1β, and IL-6), resulting in inflammation [107,108]. Furthermore, alcohol metabolites (acetaldehyde and acetate) [109], free fatty acids [110], and reactive oxygen species (ROS) [111] contribute to chronic alcohol feeding-induced inflammation. Lipid homeostasis plays a central role in the pathogenesis of ALD. Many data have shown that CES1 is essential in regulating lipid homeostasis in liver [1,52,60], intestine [59], and macrophages [51]. CES1 is also a phase I drug metabolizing enzyme, which can metabolize xenobiotics. It is plausible to speculate that CES1 may be involved in alcoholinduced hepatotoxicity. In addition, Over-expression of human CES1 increases the production of fatty acyl ethyl esters (FAEEs) via transesterification of short-chain and 57 long-chain fatty acids with ethanol [112]. FAEEs are toxic byproduct of alcohol abuse. So far, the role of CES1 in the development of ALD is unknown. Hepatocyte nuclear factor 4 (HNF4) is a nuclear hormone receptor that is constitutively active to regulate lipid, glucose and bile acid metabolism. Loss of hepatic HNF4 causes fatty liver by reducing very low-density lipoprotein (VLDL) secretion. HNF4 expression is markedly reduced in diabetes, obesity, non-alcoholic fatty liver disease (NAFLD) and high fat diet (HFD) feeding, likely as a result of increased free fatty acids, cholesterol and miR-34a expression [113]. A previous study shows that farnesoid X receptor (FXR) regulates CES1 expression [1]. It is unclear whether HNF4 also regulates CES1 expression. The present studies determined the role of CES1 in the development of ALD. We found that alcohol reduces the expressions of CES1 and HNF4 in patients with alcoholic steatohepatitis and in mice treated with alcohol. HNF4DR1 response element is identified in Ces1 promoter. Adenovirus-mediated knockdown of hepatic CES1 (AdshCes1) deteriorates alcohol-induced hepatic steatosis and liver inflammation. Compared with wild-type mice, Ces1 mice does not show aggravated hepatic steatosis in response to alcohol treatment, whereas Ces1 mice display aggravated liver inflammation upon ethanol consumption and methionine-choline deficient (MCD) diet feeding. Hepatic acetaldehyde levels, mitochondrial ROS and hepatic MDA levels are markedly increased in ethanol-fed Ces1 mice, compared with ethanol-fed wild-type mice. 58 2.2 METHODS Mice, diets and human ALD samples. Liver-specific Hnf4 mice were generated by crossing Hnf4fl/fl mice with albumin-Cre mice (all from the Jackson Laboratory. Bar Harbor, ME). Ces1 mice were generated by replacing exon 1 with an ACNLacZ cassette as described (30). Detailed description of the Ces1 mice will be reported elsewhere. Ces1 mice were backcrossed with C57BL/6 mice for at least 5 generations prior to experimentation. MCD diets were purchased from Harlan Laboratories (Cat # TD.90262; Madison, WI). LieberDeCarli diets were purchased from Bio-Serv (Flemington, NJ). Human ALD liver samples were obtained from the Liver Tissue Cell Distribution System at University of Minnesota. All the animal experiments were approved by the Institutional Animal Care and Use Committee at Northeast Ohio Medical University (NEOMED) and the use of human tissue samples were approved by Institutional Review Board at NEOMED. Chronic-binge alcohol feeding. C57BL/6J mice (12 weeks old) were administered control Lieber-DeCarli diet (cat#F1259SP, BioServ) for 5 days. On the 6th day, mice were fed an ethanol LieberDeCarli diet (cat#F1258SP, BioServ) containing 5% (vol/vol) ethanol or pair-fed a control Lieber-DeCarli diet for 10 days [114]. On the 16th day, mice were gavaged with a single dose of ethanol (5g/kg body weight) or isocaloric maltose dextrin. For some studies, on the 6th day, mice were also injected i.v. with either Ad-shLacZ or Ad-shCes1. 59 RNA isolation and quantitative real-time PCR. Total RNA was isolated using TRIzol Reagent (Life Technologies, NY). mRNA levels were determined by quantitative reverse-transcription polymerase chain reaction (qRT-PCR) on a 7500 real-time PCR machine from Applied Biosystems (Foster City, CA). Relative mRNA levels were calculated using the comparative cycle threshold (Ct) method and were normalized to the values of 36B4 mRNA levels. Western blotting. Tissues were homogenized in ice-cold modified RIPA buffer and protein concentrations were determined using a Pierce BCA Protein Assay Kit (Thermo Scientific, IL). Antibodies against mouse CES1, HNF4 and -actin were purchased from Abcam (ab45957, Cambridge, MA), Santa Cruz Biotechnology (sc6556, Dallas, TX) and Novus Biologicals (NB600-501, Littleton, CO), respectively. Transient Transfection and Mutagenesis assays. Ces1 promoter regions (-1.9 kb, -0.98 kb, -0.3 kb, -0.25 kb and -0.21 kb) were cloned to pGL3-basic plasmid (Promega, Madison, WI).Transient transfection assay was performed as described [115]. pGL3-Ces1 luciferase reporter constructs were transfected into HepG2 cells using lipofetamine 3000 (Invitrogen, CA) along with either pCDNA3 or pCDNA3-HNF4 plasmid. After 36 hrs, luciferase activities were determined and normalized to -galactosidase activity. The mutant pGL3 promoter-luciferase construct (pGL3-Ces1-mut(-1.9k))was generated using a QuickChange Site-directed Mutagenesis 60 kit from Agilent (Santa Clara, CA). Subsequently, the lucifrease activity of the mutant was tested. Chromatin immunoprecipitation (ChIP) assay. ChIP assay was performed following the manufacturer’s instructions (cat#17-295, Millipore, MA). Antibody against HNF4 was purchased from Santa Cruz Biotechnology (sc6556, Dallas, TX). CAGAACACTGAGGTTTGAATTCC The primer (forward) sequences were and TCACACCGACCTAGAGTTTAAAC (reverse), which amplified a fragment between 250bp and -300bp in the Ces1 gene promoter. Electrophoretic Mobility Shift Assay (EMSA). Oligonucleotides were labeled by biotin on the 3’ end following the manufacturer’s instruction (cat#89818, Thermo Fisher Scientific). Then, EMSA was performed using a kit purchased from Thermo Fisher Scientific and following the manufacturer’s instructions (cat# 20148, Thermo Fisher Scientific). The HNF4 protein was made using TnT T7 Quick Coupled Tanscription/Translation Reactions kit (cat# L1170, Promega) and pCDNA3-hnf4 plasmid. The wild-type oligonucleotide sequences used for EMSA were 5’- CCCTGTCTGAAGGCCTGCTGTGCTACTCTCTGCCTTTGGGAGGCCGACAG-3’ (top strand) and 5’-CTGTCGGCCTCCCAAAGGCAGAGAGTAGCACAGCAGGCCTT CAGACAGGG-3’ (bottom strand). The mutant oligonucleotide sequences were 5’CCCTGTCTGAAGGCCTGCTGTGTTACTTTTTGTTTTTGGGAGGCCGACAG-3’ 61 (top strand) and 5’- CTGTCGGCCTCCCAAAAACAAAAAGTAACACAGCAGGCCTCCAGACAGGG-3’ (bottom strand). Adenovirus. Ad-shCes1 was designed using BLOCK-iT RNAi Designer (Invitrogen, CA), annealed, and ligated to pEnter/U6 vector (Invitrogen, CA). The oligonucleotide sequences were ’GCTGATTCCAGCAGTATTGACGAATCAATAGCTGCTGGAATCAGC-3’ 5(top strand) and 5’-GCTGATTCCAGCAGCTATTGATTCGTCAATAGCTGCTGGAATAGC-3’ (bottom strand). All the adenoviruses were grown in 293A cells and purified by Cesium chloride gradient centrifugation. About 2x109 plaque formation units (pfu) of adenovirus was transfused into each mouse intravenously. Primary hepatocyte isolation. Mouse primary hepatocytes were isolated as described [65,66]. Mice were anaesthetized by intraperitoneal injection of 50 mg/kg pentobarbital. The portal vein was cannulated with a 23-gauge plastic cannula. Mouse livers were perfused with Hank’s Balanced Salt Solution (HBSS, cat#14170-112, Thermo Fisher Scientific) containing 0.19g/L EDTA. Simultaneously, the inferior vena cava was cut open. Subsequently, livers were perfused with HBSS, calcium, magnesium buffer (cat#14025092, Thermo Fisher Scientific) with 0.8mg/mL Collagenase from Clostridium histolyticum type IV 62 (Sigma, St. Louis, MO). Primary hepatocytes were released and collected in a 50 mL centrifuge tube. After centrifugation at 50g for 5 minutes and washing with DMEM, cells were cultured in 6-well plate pre-coated with 0.1% gelatin in a 2 mL of DMEM+10% FBS. Measurement of hepatic acetaldehyde level using High Performance Liquid Chromotography (HPLC). The hepatic acetaldehyde level was determined using derivatization with DNPH, followed by HPLC separation described previously [116]. 100mg liver tissue was homogenized in 3M perchloric acid. The pH of the solution was adjusted immediately to 4.0 using 2 volumes 3M sodium acetate buffer, pH9.0. After centrifugation (12000 rpm at 4 C for 20 minutes), the supernatant was transferred into an ice-cold tube, followed by addition of 80-fold molar excess 2,4-dinitrophenylhydrazine (DNPH, Cat# 119266, Sigma-Aldrich) in 6N HCl. The mixture was then placed on a shaker for 1 hour at room temperature. Derivatization was stopped with 3 volumes of 3M sodium acetate buffer, pH 9.0. Two volumes of acetonitrile were added to extract AcH-DNP. After centrifugation (10000g for 5 minutes at 4 C), the organic phase was condensed to 50 L. The ultra high performance liquid chromatograph (UHPLC) machine was purchased from Shimadzu Corp. (Columbia, MD). A Restek C18 HPLC column (25cm x 4.6mm i.d.,5m) coupled with an Ultra C18 guard column (10mm x 4mm i.d.) were purchased from Fisher Scientific (Pittsburgh, PA). The elution program was described previously [116]. An AcH-DNP standard (Sigma-Aldrich, St.Louis. MO) was used to 63 create a standard curve. The values of the area under the curve of the AcH-DNP peaks were determined to calculate the concentration of acetaldehyde in each sample. Malondialdehyde (MDA) assay. MDA assay was performed as described [115]. Buffer I was made by dissolving 2-Thiobarbituric acid in 10% perchloric acid to a final concentration 0.67%. Assay buffer was made by adding 20% trichloroacetic acid to buffer I (2:3,v/v), mixed well. 1,1,3,3,tetraethoxypropane was used as a standard. 30mg liver tissue was homogenized with 500L saline. After centrifugation, 100L supernatant was collected and added to 1mL assay buffer. The reaction mixture was incubated at 95 °C for 30 minutes, cooled down and centrifuged at 3000rpm for 10 minutes. The absorbance was measured at OD532nm. Mitochondrial H2O2 assay. Approximately 50 mg of liver tissue was homogenized in 1 mL of a buffer containing 25mM Hepes pH7.4, 1mM EDTA, 0.25M sucrose, 2mM MgCl2, 1M butylated hydroxytoluene (BHT), 1:200 dilution of Sigma P8340 and 1M diethylenetriaminepentaacetic acid. The homogenate was centrifuged at 500g for 5 minutes to pellet nuclei and debri. Then, the supernatant was centrifuged at 10,000g for 10 minutes at 4°C to obtain Mitochondria. The pellets were resuspended with 150 L homogenization buffer. About 20L solution was saved for measuring protein concentration. Mitochondrial H2O2 was detected using Amplex Red assay as described [117]. The working solution was prepared using 100L of 10mM Amplex Red reagent (Thermo 64 Fisher Scientific), 2L of 1000U/mL horseradish peroxidase (HRP) and 10mL of 50mM potassium phosphate pH7.7 with 0.5mM diethylenetriaminepentaacetic acid. 50 L of sample or standard were pipetted into 96 plates, followed by addition of 50 L of the Amplex Red reagent/HPR working solution. The reaction mixture was incubated at room temperature for 30 minutes, protected from light. The fluorescence was then measured with excitation in the range of 530-560 nm and emission at 590nm. Mitochondrial H2O2 levels were normalized to the concentration of protein. Lipid Analysis. Plasma triglyceride and cholesterol were measured using Infinity reagent from Thermo Scientific (Waltham, MA). Plasma and hepatic free fatty acid was measured using kit from Wako Diagnostics (Richmond, VA). To measure lipids in liver, approximately 100mg liver tissue was homogenized in methanol and extracted in chloroform/methanol (2:1 v/v). Hepatic triglyceride and cholesterol levels were then quantified using Infinity reagents from Thermo Scientific (Waltham, MA). Plasma Alanine Aminotransferase (ALT) and Aspartate Aminotransferase (AST) analysis. Plasma ALT and AST levels were determined using Infinity reagent (Middletown, VA) following the manufacture’s instruction. 65 Statistical Analysis. The data were analyzed using unpaired Student t test and ANOVA (GraphPad Prisim, CA). All values were expressed as meanSEM. Differences were considered statistically significant at P<0.05. 66 2.3 RESULTS Alcohol reduces CES1 and HNF4 levels in patients with alcoholic steatohepatitis and in mice treated with alcohol. To investigate whether CES1 is associated with the development of ALD, we investigated the expression of CES1 in patients with alcoholic hepatitis. Hepatic CES1 mRNA level was reduced by 75% (Figure 16A) and protein level decreased by ~ 85% (Figure 16B). Interestingly, the nuclear receptor HNF4 was also markedly repressed by >84% in both mRNA (Figure 16A) and protein (Figure16B) levels. In mice chronically fed a Liber-DeCarli ethanol diet for 10 days, hepatic Ces1 and Hnf4 mRNA levels were decreased by ~ 50% (Figure 16C). In addition, ethanol treatment significantly repressed Ces1 and Hnf4 mRNA levels in primary hepatocytes (Figure 16D). Thus, the data of Figure1 indicate that ethanol inhibits hepatic CES1 and HNF4 expression in both mice and humans. 67 Figure 16. CES1 and HNF4 expressions are reduced in patients with alcoholic steatohepatitis and in mice. Liver were collected from normal individuals and patients with ALD, mRNA (A) and protein (B) levels of CES1 and HNF4 were measured in patients. C57BL6 mice were fed an ethanol Lieber-DeCarli diet containing 5% ethanol for 10 days followed by a single dose of ethanol administration (5g/kg), mRNA levels were determined (C). Mouse primary hepatocytes were treated with 0mM, 50mM and 100mM ethanol for 24 hours. Then, CES1 and HNF4 mRNA levels were tested (D). *p<0.05, **p<0.01. 68 CES1 is regulated by HNF4and is its direct target. Then, we determined how CES1 expression was regulated by alcohol. We previously reported that CES1 is regulated by nuclear receptor farnesoid X receptor. We then tested whether CES1 is regulated by HNF4. Figure 17 A and B showed that overexpression of HNF4increased mRNA levels of Ces1 in HepG2 cells and mouse primary hepatocytes with inductions of 2.3 and 2.9 fold, respectively. In mice injected with Ad-hHNF4, the mRNA level of Ces1 was increased by ~1.8 fold (Figure. 17 C). In contrast to increased Ces1 mRNA level by over-expression of HNF4, CES1 mRNA (Figure. 17D) and protein (Figure. 17E) levels were reduced ~50% in liver-specific Hnf4mice. These data together suggested that HNF4 regulates CES1 expression both in vitro and in vivo. Next, we examined whether CES1 was a direct target of HNF4. Promoter luciferase assays showed that HNF4 induced the luciferase activity by 10 fold, 4 fold and 6 fold in the 1.9 kb, 0.98 kb and 0.3 kb Ces1 promoters, respectively, but not in the 0.25 kb Ces1 promoter (Figure 18A), suggesting that HNF4 may bind to the response element(s) located between 0.3 kb and 0.25 kb of the Ces1 promoter. Indeed, there was a potential DR-1 element (direct repeat separated by one base pair) between 300 bp and 287 bp upstream of the transcription start site (Figure 18 B). Mutation of the DR-1 element abolished the induction of Ces1 promoter activity by HNF4 (Figure 18 A). The chromatin immunoprecipitation assay showed that HNF4 protein bound to the Ces1 promoter containing the DR-1 element in the liver (Figure 18C). Finally, electrophoretic mobility shift assays showed that HNF4 protein bound to the DR-1 element and this 69 binding was competed away by the wild-type but not mutant DR-1 oligos (Figure 18D, left panel). In addition, an HNF4 antibody was able to supershift the DNA/protein complex (Figure 18D, right panel), indicating that HNF4 binds to the DR-1 element in vitro. Collectively, the data of Figs. 17 and 18 demonstrate that HNF4 regulates Ces1 expression by binding to a DR-1 element located between 300 bp and 287 bp upstream of the transcription start site. 70 Figure 17. CES1 is regulated by HNF4. HepG2 cells (A) and mouse primary hepatocytes (B) were infected with empty adenovirus (Ad-empty) and adenovirus expressing human HNF4(Ad- hHNF4n=3). After 48h, Ces1 mRNA levels were analyzed. C57BL6 mice were injected with Ad-empty and Ad-hHNF4n=6). 7 days later, hepatic Ces1 mRNA levels were tested (C). Hepatic CES1 mRNA (D) and protein levels (E) were measured in Hnf4fl/fl and L-Hnf4mice (n=5). *p<0.05. 71 Figure 18. CES1 is a direct target of HNF4. Luciferase assay was performed using pGL3-Ces1 and pGL3-Ces1-mut luciferase reporter constructs together with pCDNA3 and pCDNA3-HNF4 plasmids (n=6). After 36 hrs, luciferase activities were determined and normalized to -galactosidase activity (A). Wild-type (WT) HNF4 response element in Ces1 gene promoter was shown in the top. The mutant HNF4 response element was shown at the bottom (B). Chromatin Immunoprecipitation (ChIP) assay was performed using liver lysate and chromatin enrichment was determined (C). EMSA assays were performed using in vitro transcribed/translated proteins. Wild-type (WT) and mutant (MUT) oligos were used in the competition assays (left panel). Supershift assays were performed in the presence of an HNF4 antibody (right panel) (D). *p<0.05, **p<0.01. 72 Hepatic CES1 deficiency exacerbates alcohol-induced hepatic steatosis and inflammation. Our initial study showed that over-expression of Ces1 prevented alcohol-induced lipid accumulation in AML-12 cells (Figure.19). This result coupled with decreased expressions of CES1 in patients with alcoholic steatohepatitis implicates that CES1 plays a crucial role in the pathogenesis of ALD. To test this hypothesis, we injected the C57BL/6 mice with Ad-shLacZ or Ad-shCes1 together with Liber-DeCarli ethanol diet administration as described previously [114]. Their body weights and food intake were comparable (Figure. 20A-B). Alcohol treatment led to increased plasma levels of triglyceride, total cholesterol, free fatty acids (FFA) in wild-type (shLacZ) mice (Figure. 20C-F). These observations were consistent with previous reports [101,114]. Compared with ethanol-fed mice injected with shLacZ, ethanol-fed mice injected with shCes1 displayed elevated plasma triglyceride level (1.8 fold), slightly reduced plasma total cholesterol level and unchanged plasma FFA and hepatic total cholesterol levels (Figure.20C-F). 73 Figure 19. Over-expression of hepatic Ces1 protects against alcohol-induced triglyceride accumulation in AML12 cells. AML12 cells were cultured in a 6-well plates and infected with Ad-GFP and Ad-Ces1, followed by treatment with 50mM ethanol for 24 hours. After 24 hours, cells were collected in 100L PBS buffer. 20L of cell lysate were used for testing protein concentration. Lipids were extracted using methanol: chloroform (1:2 v/v). Triglyceride levels were tested using Infinity kit (Waltham, MA). 74 30 20 10 0 shLacZ triglyceride(mg/dL) C Control EtOH 250 ** ** 200 150 * 100 50 0 shLacZ FFA(M) 1500 1000 shCes1 E Control EtOH * * 500 0 shLacZ food intake shLacZ shCes1 15 10 5 0 y1 y2 y3 y4 y5 y6 y7 y8 y9 10 da da da da da da da da da ay d shCes1 Total cholesterol (mg/mL) body weight (g) 40 before after Lieber DeCarli ethanol diet (mL) body weight D 200 ** 150 * Control EtOH * 100 50 0 shLacZ Total cholesterol (g/mg) A B F Control EtOH 5 4 3 2 1 0 shLacZ shCes1 shCes1 shCes1 Figure 20 hepatic CES1 deficiency alters plasma lipid levels in response to alcohol treatment. C57BL6 mice were subject to alcohol feeding described in figure 5. Their body weights were shown before and after alcohol feeding (A). Food intake were recorded (B). Plasma triglyceride, total cholesterol and FFA levels were tested (C-E). Total cholesterol levels in the liver were determined and normalized to liver tissue weight (F). 75 Furthermore, we found that knockdown of hepatic Ces1 (shCes1) significantly increased hepatic TG and FFA levels (Figure. 21 A-B) in ethanol-fed mice. The higher triglyceride and FFA levels may result from increased lipogenesis. Indeed, knockdown of Ces1 in ethanol-fed mice markedly increased mRNA levels of lipogenic genes, including sterol regulatory element-binding protein 1 (Srebp-1), fatty acid synthase (Fas), acetylCoA carboxylase-1 (Acc-1), acetyl-CoA carboxylase-2 (Acc-2), diacylglyceriol Oacyltransferase 2 (Dgat-2), and peroxisome proliferator-activated receptor (Ppar ) (Figure. 21C). Oil Red O staining revealed that ethanol-fed mice injected with shCes1 had a higher degree of hepatic steatosis than those injected with shLacZ (Figure. 21D). 76 Figure 21. Hepatic CES1 deficiency exacerbates alcohol-induced hepatic steatosis. C57BL/6J mice (12 weeks old) were administered control Lieber-DeCarli diet for 5 days. On the 6th day, mice were injected with Ad-shLacZ or Ad-shCes1. Then, mice were either fed an ethanol Lieber-DeCarli diet containing 5% (vol/vol) ethanol or pair-fed a control Lieber-DeCarli diet for 10 days. On the 16th day, mice were gavaged with a single dose of ethanol (5g/kg body weight) or isocaloric maltose dextrin. Hepatic triglyceride (A) and FFA (B) levels were measured. mRNA levels of lipogenic genes, including sterol regulatory element-binding protein 1 (Srebp-1), fatty acid synthase (Fas), acetyl-CoA carboxylase-1 (Acc-1), acetyl-CoA carboxylase-2 (Acc-2), diacylglyceriol O- acyltransferase 2 (Dgat-2), and peroxisome proliferator-activated receptor (Ppar ) were tested (C). Liver sections of mice injected with Ad-shLacZ or Ad-shCes1 were subject to Oil Red O staining (D). *p<0.05, **p<0.01, #p<0.05. In figure 4C, * represents 77 comparison between ethanol and control diet. # represents comparison between shCes1 and shLacZ. 78 Next, we determined whether hepatic CES1 deficiency resulted in more liver injury. Plasma alanine aminotransferase (ALT) and aspartate aminotransferase (AST) levels were higher in ethanol-fed mice injected with shCes1 (Figure. 22A-B). Compared with ethanol-fed control mice, Ces1 deficiency increased mRNA levels of tumor necrosis factor (Tnf), chemoattractant interleukin-1 protein-1 (Mcp-1) (IL-1), interleukin-6 (Figure. 22C-F). (IL-6) and Interestingly, monocyte plasma - hydroxybutyrate levels were increased in ethanol-fed wild-type mice, but not in the ethanol-fed hepatic Ces1 deficient mice (Figure. 22G), suggesting that hepatic CES1 deficiency may cause mitochondrial dysfunction. Collectively, hepatic CES1 deficiency exacerbated alcohol-induced hepatic steatosis and liver inflammation. 79 Figure 22. Hepatic CES1 deficiency exacerbates alcohol-induced liver inflammation. Mice were subject to alcohol feeding as described in figure 4. Plasma ALT and AST 80 levels were measured (A and B). mRNA levels of tumor necrosis factor (Tnf) (C), interleukin-1 (IL-1) (D), interleukin-6 (IL-6) (E) and monocyte chemoattractant protein-1 (Mcp-1) (F) were determined. Plasma -hydroxybutyrate level was tested (G). *p<0.05, **p<0.01. 81 Global deletion of CES1 does not change alcohol-induced hepatic steatosis, but exacerbates liver inflammation. To test whether global deletion of CES1 exacerbates alcohol-induced liver injury, Ces1 and Ces1 mice were administered a Lieber-DeCarbli ethanol diet containing 5% ethanol for 10 days followed by a single dose of ethanol (3g/kg) administration as described [114]. Protein expressions of hepatic CES1 were completely abolished in Ces1 mice (Figure. 23A). The chronic-binge feeding did not affect their body weights (Figure. 23B). Compared with ethanol-fed Ces1mice, ethanol-fed Ces1mice displayed moderately higher plasma triglyceride (~35%) and mildly lower total cholesterol (~20%) levels (Figure. 23C-D). Surprisingly, hepatic triglyceride and hepatic total cholesterol levels were comparable (Figure. 23E-F). 82 B body weight before after body weight (g) 40 30 20 10 250 200 150 Control EtOH * ** 100 50 0 Ces1 E Triglyceride(g/mg) * Control EtOH 40 30 Ces1 * * 20 10 0 Ces1 Ces1 Total cholesterol (g/mg) Triglyceride(mg/dL) C Total cholesterol (mg/mL) 0 Ces1 D Ces1 Control EtOH 150 100 10 50 0 Ces1 F * 8 Ces1 Control EtOH 6 4 2 0 Ces1 Ces1 Figure 23. global deletion of Ces1 does not exacerbate alcohol-induced hepatic steatosis. Wild-type and Ces1 mice were subject to alcohol feeding described in the legend of figure 6. The protein levels of hepatic CES1 was shown in (A). Body weights were shown before and after alcohol feeding (B). Plasma triglyceride (C) and total cholesterol (D) levels were tested. Hepatic triglyceride (E) and total cholesterol (F) levels were determined. 83 Furthermore, ethanol feeding significantly increased plasma levels of ALT (Figure. 24A) and AST (Figure. 24B) in both Ces1+/+ and Ces1 mice, and such increases were further enhanced by Ces1 deficiency. In addition, ethanol significantly induced hepatic the mRNA levels of Tnf (Figure. 24C), IL-1 (Figure. 24D), IL-6 (Figure. 24E) and Mcp-1 (Figure. 24F) in Ces1 mice, and the induction of Tnf, Il1 and Il6 was not significant in Ces1 mice (Figure 24C-24D). Importantly, Ces1 deficiency significantly potentiated ethanol-induced Tnf, Il1, Il6 and Mcp1 mRNA levels in the liver (Figure 24C-24F). Overall, figure 23 and 24 demonstrate that global deletion of CES1 exacerbates alcohol-induced liver inflammation, but does not change hepatic steatosis. 84 Figure 24. Global deletion of CES1 exacerbates alcohol-induced liver inflammation. Ces1 and Ces1 mice were administered control Lieber-DeCarli diet for 5 days, followed by ethanol diet or pair-fed control diet administration for 10 days and a single dose of ethanol (3g/kg body weight) gavage on the 16th day. Plasma ALT (A) and AST (B) levels were measured. mRNA levels of Tnf (C), IL-1 (D), IL-6 (E) and Mcp-1(F) were tested. *p<0.05, **p<0.01. 85 Global deletion of CES1 increases MCD diet-induced liver inflammation. We then tested whether global deletion of CES1 affects MCD diet-induced liver injury. MCD diet results in liver injury similar to human nonalcoholic steatohepatitis (NASH). There was a 21% increase in hepatic TG level but unchanged hepatic cholesterol level (Figure. 25A). Fat mass of Ces1 mice was higher (5.03%) than that in wild-type mice (3.07%) (Figure. 25B). Plasma ALT level and mRNA levels of TnfIL1IL-6, and transforming growth factor (Tgf) were potentiated in MCD diet-fed Ces1 mice (Figure. 25C-G), suggesting that CES1 deficiency aggravates MCD dietinduced liver inflammatory response and liver injury. Global deletion of CES1 does not change MCD diet-induced liver fibrosis. We further determined MCD diet-induced liver fibrosis in wild type and Ces1 mice. Ces1 deficiency did not potentiate the expressions of figrogenic genes (Figure. 26), neither did it aggravate MCD diet-induced fibrosis, which is shown by unchanged degree of fibrosis in Sirius red stained liver sections (Figure. 27). 86 Figure 25. Global deletion of CES1 aggravates MCD diet-induced liver injury. Ces1 and Ces1 mice were fed a MCD diet for 4 weeks. At the end of MCD diet 87 feeding, hepatic lipids were tested (A). Fat and lean mass were measured using EchoMRI (B). Plasma ALT level (C) and mRNA levels of Tnf (D), IL-1 (E), IL-6 (F) and TGF(G) were determined. *p<0.05, **p<0.01. Figure 26. CES1 deficiency does not change fibrogenic gene expressions. Ces1 and Ces1 mice were fed an MCD diet for 4 weeks. mRNA levels of -SMA ( -smooth muscle actin), TGF-, Col1 (collagen 1) and TIMP (tissue inhibitor of metalloproteinase) were determined. 88 Figure 27. Global deletion of CES1 does not exacerbate MCD diet-induced fibrosis. Sirius Red staining was performed using the livers collected from MCD diet-fed mice. 89 Global deletion of CES1 results in increased hepatic acetaldehyde level, elevated oxidative stress and mitochondrial dysfunction To define how Ces1 deficiency aggravates ethanol-induced liver injury, we analyzed acetaldehyde level in the liver. Acetaldehyde is a reactive compound, and is highly toxic to hepatocytes because it can sensitize cells to oxidative stress or other damaging signals, ultimately lead to mitochondria damage and cell death [92,118]. At the basal level, hepatic acetaldehyde level was increased by 2.2 fold in Ces1 mice than in Ces1 mice (Figure 28A). Upon ethanol feeding, hepatic acetaldehyde level was increased by 1.7 fold in Ces1 mice, 2 fold in Ces1mice and 2.4 fold in Ces1 mice than in Ces1 mice (Figure 28A). Consistent with this observation, the mRNA level of acetaldehyde dehydrogenase 2 (Aldh2) was reduced by 50% in Ces1 mice (p<0.05; data not shown). Oxidative stress is an important factor in the pathogenesis of ALD. Mitochondrial reactive oxygen species (ROS) triggers proinflammatory cytokine production, damages mitochondrial DNA and promotes lipid peroxidation [119,120,121]. We examined whether the aggravated liver inflammation in Ces1 mice was associated with increased ROS. Indeed, figure 28B showed that mitochondrial H2O2 level was increased more than 3 fold in ethanol-fed Ces1 mice compared with ethanol-fed wild-type mice. Consistently, malondialdehyde (MDA) level was higher (2 fold) in the livers of ethanolfed Ces1 mice (Figure. 28C). The increased production of ROS is a crucial cause of mitochondrial DNA mutation, degradation and mitochondria dysfunction [122]. We then tested mRNA levels of genes which encode mitochondrial enzymes, including 90 cytochrome oxidase subunit I (Cox1), cytochrome b (Cyt b), NADH dehydrogenase, subunit (Nd1), and ATP synthase F0 subunit 6 (Atp6), all of which were significantly down-regulated (Figure. 28D). Thus, global deletion of CES1 led to increased alcoholinduced ROS production, increased MDA and acetaldehyde formation, possibly resulting in mitochondrial dysfunction. 91 B ** * 200 150 * 100 ** 50 0 Ces1 * C Relative mRNA MDA(mol/g) * 2 0 Ces1 E 400 Control EtOH 300 * 200 100 0 Ces1 100 Ces1 * * 50 0 Ces1 Ces1 Ces1+/+ , EtOH Ces1-/-, EtOH 1.0 * * 0.5 * 0.0 Ces1 * Control EtOH D 1.5 ** Control EtOH 4 150 Ces1 6 FFA(nmol/mg protein) H2O2 (M/mg protein) 250 Control EtOH Cox1 -hydroxybutyrate(M) Acetaldehyde (nmol/g) A Cytb Nd1 F 1000 800 Atp6 Control EtOH * * 600 400 200 0 Ces1 Ces1 Figure 28. Global deletion of CES1 increases hepatic acetaldehyde level and oxidative stress. Ces1 and Ces1 mice were subject to alcohol feeding as described in the legend of figure 24. Hepatic acetaldehyde levels were measured using HPLC (A). Mitochondrial H2O2 levels were measured using liver lysate and normalized to protein concentration (B). MDA assay was performed using liver lysate and normalized to liver 92 weight (C). mRNA levels of mitochondrial genes were tested in ethanol-fed Ces1 and Ces1 mice (D). -hydroxybutyrate (E) and FFA (F) levels were determined. *p<0.05, **p<0.01. 93 Global deletion of CES1 inhibits alcohol-induced elevation of -hydroxybutyrate, but it enhances hepatic FFA level. Ethanol feeding significantly increases acetate level and therefore the synthesis of -hydroxybutyrate (-HB). On the other hand, elevated mitochondrial FAO is also accompanied by increased -HB synthesis. As such, plasma -HB level may reflect mitochondrial functions under normal conditions. As shown in Figure 28F, ethanol significantly increased plasma -HB level in Ces1 mice but not in Ces1 mice. In addition, ethanol induced a 2.5-fold increase in hepatic free fatty acid (FFA) level in Ces1 mice but not in Ces1 mice (Figure 28E). The increase in hepatic FFA level was not accompanied by any change in genes involved in fatty acid synthesis (Srebp1c, Fas, Acc1, Acc2), transport (CD36, Fabp1) or oxidation (Ppar, Cpt1, Mcad, Acox1, and Acox2) (Figure 29). These data suggest that Ces1 mice may have an impaired mitochondrial function. 94 Figure 29. global deletion of CES1 does not change mRNA levels of genes involved in fatty acid metabolism. mRNA levels of genes were tested in ethanol-fed wild-type and Ces1 mice. These genes are sterol regulatory-element binding protein 1c(Srebp1c), fatty acid synthase (Fas), acetyl-CoA carboxylase 1 (Acc1), acetyl-CoA carboxylase 2 (Acc2), cluster of differentiation 36 (CD36), fatty acid binding protein 1 (Fabp1), peroxisome proliferator activated receptor (Ppar ), carnitine palmitoyltransferase 1-a (Cpt1-), medium-chain acyl coenzyme A dehydrogenase (Mcad1), peroxisomal acylcoenzyme A oxidase 1 (Acox1), peroxisomal acyl-coenzyme A oxidase 2 (Acox2). 95 2.4 DISCUSSION In the current study, we find that mRNA and protein expressions of CES1 and HNF4 are markedly reduced in patients with alcoholic steatohepatitis and in mice treated with alcohol. We clearly demonstrate that CES1 is transcriptionally regulated by HNF4 and that it is a direct HNF4 target. Then, using chronic-binge alcohol feeding, we find that hepatic CES1 deficiency exacerbates hepatic steatosis and inflammation in response to alcohol challenge. In addition, global deletion of CES1 does not change alcohol-induced hepatic steatosis, but it exacerbates alcohol and MCD diet-induced liver inflammation, likely through increased hepatic acetaldehyde level, elevated oxidative stress and enhanced lipid peroxidation. Alcohol suppresses CES1 expression through, at least in part, repressing HNF4. HNF4 is known to orchestrate a variety of genes associated with lipid, glucose and bile acids metabolism, differentiation and morphogenesis [123]. Our results suggest that ethanol inhibits CES1 expression through, at least in part, repressing HNF4. However, how ethanol suppresses HNF4 expression is unknown. It is worth to note that transcriptional activity of HNF4 is inhibited by unsaturated fatty acyl-CoAs with greatest inhibition by C18:3, -3, and C20:5, -3 [124]. In addition, HNF4 protein level is reduced by alcohol treatment in Sprague Dawley rats, but it is restored in alcohol combined with saturated fatty acids treatment [125]. Extensive studies show that HNF4 protein level is decreased by unsaturated linoleic acid in HepG2 cells and primary hepatocytes [125,126]. On the other hand, it is known that alcohol feeding alters fatty 96 acid composition. In absolute terms, alcohol feeding significantly increases hepatic levels of monounsaturated FA (MUFA) and polyunsaturated FA (PUFA), but it does not change hepatic saturated FA (SFA) levels [127]. Accordingly, we hypothesize that the increased level of unsaturated fatty acids as a result of alcohol administration may partly contribute to the suppression of HNF4. Given that tissue specific loss of HNF4in the liver results in increased lipid deposits in hepatocytes [128], the suppression of HNF4CES1 axis may have profound impact on the pathogenesis of alcohol-induced hepatic steatosis. In addition, under the conditions of common metabolic stress (diabetes, obesity and NAFLD, HFD feeding), elevated cholesterol, p53 and miR-34a orchestrate to inhibit HNF4 expression. Thus, it is also possible that other unknown mechanism(s) are involved in inhibiting HNF4 expression. Global CES1 deficiency does not exacerbate alcohol-induced hepatic steatosis. An unexpected finding in this study is that unlike ethanol-fed mice injected with shCes1 which had increased ethanol-induced hepatic steatosis, ethanol-fed Ces1 mice do not display aggravated hepatic steatosis. It is evident that CES1 also expresses in the intestine; intestinal CES1 regulates chylomicron assembly [59]. We speculate that the intestinal fat absorption may be diminished in Ces1 mice. Thus, in Ces1 mice, the net effect of the absence of intestinal and hepatic CES1 on lipid homeostasis results in unchanged hepatic triglyceride and cholesterol levels, compared with those in Ces1 mice. In addition, our previous report shows that loss of hepatic CES1 increases lipogensis [1]. Our present study further proves that lipogenic gene expression levels are 97 elevated by the loss of hepatic CES1 and alcohol feeding. This finding suggests that hepatic CES1 plays a protective role against ALD. Global deletion of CES1 exacerbates alcohol-induced liver inflammation. In the current study, global deletion of Ces1 aggravates ethanol-induced liver inflammation without affecting hepatic steatosis. Although hepatic steatosis appears to correlate with more severe forms of liver diseases which require the imposition of second “hit” to incite hepatic inflammation and fibrosis, it has been well documented that hepatocyte triglycerides per se are not hepatotoxic [129]. Our results support this point of view. On the other hand, our results show that the hepatic level of FFA, a direct hepatotoxic substance, is higher in ethanol-fed Ces1 mice than that in ethanol-fed wildtype mice. Mitochondrial gene expressions are reduced. This may be indicative of mitochondrial dysfunction, which in turn affects mitochondrial oxidation, resulting in decreased FFA disposal and increased ROS production. Indeed, hepatic FAA, mitochondrial H2O2, MDA levels are significantly elevated in ethanol-fed Ces1 mice. Furthermore, hepatic acetaldehyde, a reactive hepatotoxic compound, is markedly increased in ethanol-fed Ces1 mice. In addition to acetaldehyde, increased productions of fatty acid ethyl ester (FAEE), palmitic acid are also associated with the pathogenesis of ALD. It is shown that CES1 is involved in the synthesis of FAEE [130]. Thus, we will not rule out other contributing factors to the aggravated liver injury in Ces1 mice. In the present study, we find that -hydroxybutyrate is not induced by alcohol in Ces1 mice. -hydroxybutyrate is one of the ketone bodies which are synthesized from acetylCoA; it is increased in patients with excessive alcohol consumption [131]. Alcohol- 98 induced increase in -hydroxybutyrate level may be attributed to 1) increased acetate which in turn becomes a source of acetyl-CoA [131] and 2) the increased ratio of NADH/NAD+ which inhibits TCA cycle and promotes ketone body production. The unchanged level of -hydroxybutyrate in response to alcohol challenge in Ces1 mice remains unknown. We speculate that either the production of acetyl-CoA is reduced or TCA cycle remains intact. All together, the elevated ROS, MDA and hepatic acetaldehyde levels accompanied by increased hepatic FFA level are accountable for the aggravated liver inflammation in ethanol-fed Ces1mice. CES1 has the ability to hydrolyze amide or ester bonds. Our previous studies show that TG is a substrate for CES1 [1]. Defective TG hydrolase activity in Ces1deficient hepatocytes is responsible for elevated TG accumulation in the liver. Ethanol is unlikely to be a direct substrate of CES1. However, the metabolites of CES1 may negatively affect alcohol metabolism and mitochondrial functions, leading to accumulation of toxic substances, such as acetaldehyde and reactive oxygen species. These toxic substances are sufficient to cause damages on mitochondria and cell membranes, eventually resulting in liver inflammation and damage. One of our future directions will be to determine which metabolite(s) cause elevation of these toxic substances. In summary, we have presented novel evidence that Ces1 deficiency exacerbates alcohol-induced liver injury. Our findings further highlight that HNF4CES1 axis may play an important role in the pathogenesis of ALD. The present studies identify CES1 as a potential therapeutic target for treating ALD. 99 CHAPTER 3: THE ROLE OF CARBOXYLESTERASE 1 IN ATHEROSCLEROSIS. 3.1 INTRODUCTION Atherosclerosis-associated cardiovascular disease (CVD) is a leading cause of death in the developed countries. It is characterized by increased lipid accumulation in intima of the arterial walls, accompanied with chronic inflammation of arterial endothelium [132]. Risk factors for this disease include obesity, dyslipidemia and type 2 diabetes; and keeping theses metabolic disorders under control is an effective strategy for reducing cardiac events associated with atherosclerosis. High plasma cholesterol level is a hallmark and a direct cause of atherosclerosis. Current pharmacological intervention concentrates on reducing high cholesterol levels in patients, through enhancing cholesterol efflux from macrophages, promoting reverse cholesterol transport (RCT), enhancing plasma HDL level, and reducing plasma LDL level. Atheromatous plaques, formed from fat, cholesterol, calcium and other substances, build up in the inner lining of the arteries and impede blood flow. Life threatening events such as myocardial infarction or stroke result from the rupture or ulceration of an “unstable” plaque [133]. Inflammation and hypercholesterolemia are important features of atherosclerosis. LDLs, trapped in the subendothelial space, are subject to oxidative modification; and the oxidized lipids triggers monocyte recruitment to endothelial layer of the artery [134]. Monocytes will then penetrate into the tunica intima and acquire characteristics of the tissue macrophage. Furthermore, Foam cells, derived from 100 macrophages as a result of accumulation of lipid droplets, will secrete pro-inflammatory cytokines to amplify the local inflammatory response in the lesion [135]. The progression of lesion induces arterial calcification and predisposes to plaque rupture at sites of monocytic infiltration. Oxidized LDL induces endothelial cells and monocytes to express high levels of tissue factor in the lesion, and results in thrombosis, which is the proximate cause of the clinical event [136]. Macrophage cholesterol efflux is shown to have antiatherosclerotic effect. Free cholesterol efflux from macrophage foam cells requires ATP-binding cassette transporter A1 (ABCA1) and ATP-binding cassette transporter G1 (ABCG1), which mediate free cholesterol (FC) transportation to cholesterol-deficient and phospholipid depleted apolipoprotien (apo)A1 and HDL in plasma, respectively [137] (Figure.30). Another efflux pathway is mediated by the scavenger receptor class B type 1(SRB1), which effluxes FC to the mature HDL. In addition, SRB1 also mediates cholesterol uptake in macrophage. In macrophage, cholesteryl esters, accumulation of which causes “foamy” characteristic of macrophage, are hydrolyzed to free cholesterol, which is an antiatherosclerotic process in the body [137] (Figure.30). 101 Figure 30. macrophage cholesterol efflux. (picture is adapted from Rader, D et al. Molecular regulation of HDL metabolism and function: implications for novel therapies. J Clin Invest. 2006; 116(12))[138] The antiatheroclerotic role of CESs has been explored. Over-expression of macrophage human CES1, an equivalent form of mouse Ces3, leads to increases in cholesteryl ester (CE) hydrolysis [51], mobilization of cytoplasmic CE [57], free cholesterol (FC) efflux [58] and attenuation of atherosclerosis in Ldlr mice [51]. Overexpression of hepatic human CES1 has anti-atherogenic effects, through enhancing the elimination of cholesterol into bile [139]. The studies in chapter 3 show the role of hepatic Ces1 in the development of atherosclerosis. We find that hepatic CES1 deficiency aggravates western diet-induced atherosclerosis in ApoE mice. This may result from increased cholesterol synthesis and increased plasma cholesterol level in hepatic Ces1 deficient ApoE mice. 102 3.2 METHODS Mice and diet. 8 week old ApoE mice were purchased from Jackson Laboratory (Bar Harbor, Maine). ApoEmice were fed a western diet containing 0.21% cholesterol and 41% fat from Research diets (New Burnswick, NJ) for 1 week, followed by adenovirus injection with either shLacZ (n=7 per group) or shCes1 (n=7 per group). Mice were given a western diet for another 3 weeks. 3 weeks after adenovirus injection, mice were euthanized and their hearts and aortas were isolated as described [140,141]. Aorta isolation. The aorta, including the ascending arch, thoracic, and abdominal segments, were dissected, gently cleaned of the adventitia, and stained with Oil Red O as described previously [140,141]. In addition, the aortic roots were collected from the base of the heart including the atria and embedded in optimal cutting temperature compound. Sections (5 μm) were obtained every 50 μm from the base of the aortic leaflets to 400 μm above. After staining with Oil Red O, images were captured with a microscope, and the lesion area for each aortic ring was analyzed with Image J software [142]. Peritoneal macrophage isolation. Peritoneal macrophages were isolated according to previous report [143]. C57BL/6J mice were injected with 1mL of 3% (w/v) brewer thioglycollate medium into their peritoneal cavity. 3-5 days later, mice were euthanized. The outer skin of the 103 peritoneum was cut with gentle and pulled back to expose the inner skin lining the peritoneal cavity. DMEM medium with 10%FBS was injected into the peritoneal cavity using a 21.5g needle, followed by gently massage the peritoneum to dislodge any attached cells into the medium. The cells were collected into a 5mL syringe. DMEM medium was injected 1-2 more time to retrieve more cells. Cell suspension was spun at 1500rpm, resuspended and cultured in 6-well plates. Lipid Analysis. Plasma triglyceride and cholesterol were measured using Infinity reagent from Thermo Scientific (Waltham, MA). To measure lipids in liver, approximately 100mg liver tissue was homogenized in methanol and extracted in chloroform/methanol (2:1 v/v). Hepatic triglyceride and cholesterol levels were then quantified using Infinity reagents from Thermo Scientific (Waltham, MA). Statistical Analysis. The data were analyzed using unpaired Student t test and ANOVA (GraphPad Prisim, CA). All values were expressed as meanSEM. Differences were considered statistically significant at P<0.05. 104 3.3 RESULTS The macrophages of Ces1 mice accumulate more lipids. Our interest in studying the role of CES1 in atherosclerosis originates from previous studies of macrophage CESs which are shown to protect against atherosclerosis. In this regard, we fed wild type and Ces1 mice a western diet for 8 weeks and their macrophages were isolated. Oil red O staining revealed that more lipids accumulated in Ces1 deficient macrophages (Figure 31A). Macrophagic total cholesterol levels were higher in Ces1 mice (Figure 31B). Thus, Ces1 deficiency increased total cholesterol levels in macrophages. Figure 31. Global deletion of CES1 results in increased lipid accumulation in macrophages. Oil red O staining of peritoneal macrophages of wild type (left penal) and Ces1 mice (right penal) fed a western diet for 8 weeks (A). Total cholesterol levels were measured and normalized to protein concentration (B). 105 Loss of hepatic CES1 increases plasma and hepatic lipids in ApoE mice. To investigate whether hepatic Ces1 plays a role in atherosclerosis, ApoEmice were fed a western diet for 1 week. Then mice were then injected with either Ad-shLacZ or Ad-shCes1. Mice continued receiving western diet for another 3 weeks. 21 days after adenovirus injection, plasma, liver and aorta were collected. Figure 32 showed that plasma triglyceride, cholesterol levels were higher in Ces1 deficient ApoE mice (Figure. 32 A and B). In addition, hepatic triglyceride and cholesterol levels were higher in ApoEmice injected with shCes1 (Fig. 32 C and D). Figure 32. Loss of hepatic CES1 increases lipid contents in ApoE mice. ApoE mice were fed a western diet for 1 week, followed by adenovirus injection of shLacZ or shCes1. After virus injection, mice received a western diet for another 3 weeks. Plasma 106 triglyceride (A) and plasma cholesterol (B) were measured. Hepatic triglyceride (C) and total cholesterol (D) were determined. *p<0.05, **p<0.01. Loss of hepatic CES1 shows atherosclerotic lipid profile. Plasma HLD-C and LDL-C are significant factors for determining the atherosclerosis. Next, we determined the lipoprotein levels using fast protein liquid chromatography (FPLC). Figure 33 showed that VLDL-TG was increased in ApoE mice injected with shCes1 (Figure. 33A). Similarly, VLDL-C and LDL-C were higher in those mice (Figure. 33B). Thus, these data suggest that hepatic Ces1 deficiency has atherosclerotic lipid profile. 107 (A) Triglyceride (g/fraction) 40 sh-LacZ sh-Ces1 30 20 10 30 27 24 21 18 15 12 9 6 3 0 0 Fraction (B) Cholesterol (g/fraction) 150 sh-LacZ sh-Ces1 100 50 27 24 21 18 15 12 9 6 3 0 0 Fraction Figure 33. Loss of hepatic CES1 shows atherosclerotic lipid profile. ApoE mice were subject to adenovirus injection and western diet feeding as described in the legend of Figure 32. Plasma lipoprotein levels were measured using FPLC (A and B). 108 Loss of hepatic CES1 aggravates atherosclerosis in ApoE mice. To further determine whether loss of hepatic Ces1 affects the development of atherosclerosis, we isolated the aortas and stained them with Oil Red O. Oil red O staining revealed that loss of hepatic Ces1 increased lipid accumulations in the arterial walls (Figure. 34 A). A further quantification confirmed that the aorta lesion areas were larger in Ces1 deficient ApoE mice than those in their control counterparts (Figure. 34B). The lesion size of aortic root in Ces1 deficient ApoE mice is larger than that in control mice (Figure 34C and D). Thus, Ces1 deficiency aggravates atherosclerosis. 109 A B 10 shLacZ Lesion area% 8 shCes1 6 4 2 0 shLacZ shLacZ shCes1 D Heart lesion (mm2/section) C shCes1 1.010 5 8.010 4 6.010 4 4.010 4 2.010 4 0 shLacZ shCes1 Figure 34. Loss of hepatic CES1 aggravates atherosclerosis in ApoE mice. ApoE mice were subject to western diet feeding and adenovirus injection as described in the legend of Figure 32. Aortas of control mice (A, top penal) and Ces1 deficient mice (A, bottom penal) were isolated and stained with Oil red O. The lesion areas were analyzed using commercially available software and were shown in (B). Aortic root of control (top penal) and Ces1 deficient mice (bottom penal) was stained by oil red O (C) and lesion areas were quantified (D). 110 3.4 DISUSSION In chapter 3, we studied the role of CES1 in the development of atherosclerosis. CES3, a murine form of human CES1, has been shown to promote macrophage cholesterol efflux and cholesterol elimination in liver. In the present study, we showed that hepatic Ces1 deficiency increases plasma and hepatic lipids levels, and increases VLDL-TG, VLDL-C and LDL-C levels, all of which directly result in atherosclerosis. We have previously shown that hepatic Ces1 deficiency results in increased lipogenesis and enhanced VLDL secretion, both of which may contribute to the increased plasma lipid levels, eventually lead to atherosclerosis (Figure 32). Collectively, these data suggest that hepatic Ces1 deficiency alters systemic lipid homeostasis, likely via enhancing lipogenic genes expressions. Despite hepatic Ces1 deficient mice show striking changes in lipid homeostasis, global deletion of Ces1 does not significantly change plasma and hepatic lipid levels in response to western or high fat diet challenge, except that plasma cholesterol level is slightly decreased (~20%) (data not shown). More tissue-specific functional studies are needed to clearly understand the role of Ces1 in regulation of lipid metabolism. Since the lipid phenotype in Ces1 mice is different from that in hepatic Ces1 deficient mice, we generated Ces1Ldlrmice and these mice are fed a western diet to induce atherosclerosis. The double knockout mice will help us understand the role of macrophagic and hepatic Ces1 in the development of atherosclerosis. 111 In summary, using adenovirus gene delivery system, we find that loss of hepatic Ces1 aggravates atherosclerosis. However, more data have to be collected from Ces1Ldlrmice to clearly understand the role of Ces1 in regulation of atherosclerosis. 112 CHAPTER 4: CONCLUSION CES1 is a phase I drug metabolizing enzyme which is shown to be involved in biotransformation and detoxification of a variety of drugs and prodrugs, including angiotensin-converting enzyme inhibitors, necrotic, anti-tumor and anti-virus drugs. Given its nature of hydrolyzing ester and amide bond, CES1 is reported to have triglyceride hydrolase and cholesteryl ester hydrolase activity. Previous in vitro studies show that CES1 hydrolyzes triglyceride and promotes fatty acid oxidation. Our studies focus on in vivo elucidation and delineation of CES1 in regulation of lipid and carbohydrate metabolism. First, we find that in diabetic mouse models, CES1 expression is significantly increased; and we further demonstrate that CES1 is induced by glucose. The induction of CES1 expression, in turn, reduces plasma glucose levels, through increasing insulin sensitivity. Second, we clearly demonstrate that overexpression of hepatic CES1 protects against NAFLD, through reducing hepatic triglyceride and promoting fatty acid oxidation. In contrast, knockdown of hepatic CES1 induces hepatic steatosis by increasing SREBP processing and lipogenesis. We also find that CES1 is a farnesoid X receptor (FXR) target gene, and that activation of FXR reduces hepatic and plasma triglyceride levels through, at least in part, inducing CES1. Third, we show that CES1 is inhibited by alcohol, and that CES1 deficiency exacerbates alcohol-induced liver injury, possibly resulting from increased hepatic acetaldehyde 113 level, elevated ROS level and enhanced lipid peroxidation, suggesting that CES1 plays a protective role against ALD. We also find that CES1 is a direct target of HNF4, and HNF4-CES1 axis may have a profound impact on the development of ALD. Lastly, we demonstrate that loss of hepatic CES1 aggravates western diet-induced atherosclerosis in ApoE mice. These exciting data lead us to conclude that CES1 plays an essential role in regulating lipid and carbohydrate metabolism, and also protects against NAFLD, ALD and atherosclerosis. One of our future directions will focus on studying how CES1 regulates insulin sensitivity. Different from deletion of hepatic CES1 which causes insulin resistance, global deletion of CES1 does not change insulin sensitivity (data not shown). How CES1 affects systemic insulin sensitivity and whether CES1 in adipose tissue or muscle is involved in the regulation of insulin signaling will be our future focuses. Furthermore, Ces1 mice display reduced plasma cholesterol level (~20%) compared to wild type mice (data not shown). Therefore, we will investigate whether the fat absorption is changed in Ces1 deficient mice. In addition, we plan to use liquid chromatography-mass spectrometry (LC-MS) to study the endogenous substrates and metabolomics of hepatic CES1 in mice treated with alcohol or western diet. This research will help us understand how CES1 deficiency causes liver injury. Lastly, we are interested in investigating the pathway mediated by CES1 in regulation of the development of atherosclerosis. We plan to use macrophage and liver specific Ces1 deficient Ldlrmice as well as global Ces1 deficient Ldlrmice to study the underlying mechanism. 113 In conclusion, the present studies underscore CES1 as a potential therapeutic target in treating metabolic disorders. We propose that manipulation of CES1 expression is a plausible strategy for managing metabolic disease. 115 REFERENCES 1. Xu J, Li Y, Chen WD, Xu Y, Yin L, et al. (2013) Hepatic carboxylesterase 1 Is essential for both normal and farnesoid X receptor-controlled lipid homeostasis. Hepatology. 2. Chalasani N, Younossi Z, Lavine JE, Diehl AM, Brunt EM, et al. (2012) The diagnosis and management of non-alcoholic fatty liver disease: practice Guideline by the American Association for the Study of Liver Diseases, American College of Gastroenterology, and the American Gastroenterological Association. Hepatology 55: 2005-2023. 3. Rinella ME (2015) Nonalcoholic fatty liver disease: a systematic review. JAMA 313: 2263-2273. 4. Firneisz G (2014) Non-alcoholic fatty liver disease and type 2 diabetes mellitus: The liver disease of our age? World Journal of Gastroenterology 20: 9072-9089. 5. Sass DA, Chang P, Chopra KB (2005) Nonalcoholic fatty liver disease: a clinical review. Dig Dis Sci 50: 171-180. 6. Neuschwander-Tetri BA, Brunt EM, Wehmeier KR, Oliver D, Bacon BR (2003) Improved nonalcoholic steatohepatitis after 48 weeks of treatment with the PPAR-gamma ligand rosiglitazone. Hepatology 38: 1008-1017. 115 7. Promrat K, Lutchman G, Uwaifo GI, Freedman RJ, Soza A, et al. (2004) A pilot study of pioglitazone treatment for nonalcoholic steatohepatitis. Hepatology 39: 188-196. 8. Marchesini G, Brizi M, Bianchi G, Tomassetti S, Zoli M, et al. (2001) Metformin in non-alcoholic steatohepatitis. Lancet 358: 893-894. 9. Lavine JE (2000) Vitamin E treatment of nonalcoholic steatohepatitis in children: a pilot study. J Pediatr 136: 734-738. 10. Bataller R, Brenner DA (2005) Liver fibrosis. Journal of Clinical Investigation 115: 209-218. 11. Ananthanarayanan M, Balasubramanian N, Makishima M, Mangelsdorf DJ, Suchy FJ (2001) Human bile salt export pump promoter is transactivated by the farnesoid X receptor/bile acid receptor. Journal of Biological Chemistry 276: 28857-28865. 12. Lambert G, Amar MJ, Guo G, Brewer HB, Jr., Gonzalez FJ, et al. (2003) The farnesoid X-receptor is an essential regulator of cholesterol homeostasis. Journal of Biological Chemistry 278: 2563-2570. 13. Ma K, Saha PK, Chan L, Moore DD (2006) Farnesoid X receptor is essential for normal glucose homeostasis. Journal of Clinical Investigation 116: 1102-1109. 14. Cariou B, van Harmelen K, Duran-Sandoval D, van Dijk TH, Grefhorst A, et al. (2006) The farnesoid X receptor modulates adiposity and peripheral insulin sensitivity in mice. Journal of Biological Chemistry 281: 11039-11049. 15. Claudel T, Staels B, Kuipers F (2005) The Farnesoid X receptor: a molecular link between bile acid and lipid and glucose metabolism. Arterioscler Thromb Vasc Biol 25: 2020-2030. 117 16. Watanabe M, Houten SM, Wang L, Moschetta A, Mangelsdorf DJ, et al. (2004) Bile acids lower triglyceride levels via a pathway involving FXR, SHP, and SREBP-1c. Journal of Clinical Investigation 113: 1408-1418. 17. Fu L, John LM, Adams SH, Yu XX, Tomlinson E, et al. (2004) Fibroblast growth factor 19 increases metabolic rate and reverses dietary and leptin-deficient diabetes. Endocrinology 145: 2594-2603. 18. Fang S, Suh JM, Reilly SM, Yu E, Osborn O, et al. (2015) Intestinal FXR agonism promotes adipose tissue browning and reduces obesity and insulin resistance. Nature Medicine 21: 159-165. 19. Teodoro JS, Rolo AP, Palmeira CM (2011) Hepatic FXR: key regulator of wholebody energy metabolism. Trends Endocrinol Metab 22: 458-466. 20. Trabelsi MS, Daoudi M, Prawitt J, Ducastel S, Touche V, et al. (2015) Farnesoid X receptor inhibits glucagon-like peptide-1 production by enteroendocrine L cells. Nature Communications 6: 7629. 21. Bjursell M, Wedin M, Admyre T, Hermansson M, Bottcher G, et al. (2013) Ageing Fxr deficient mice develop increased energy expenditure, improved glucose control and liver damage resembling NASH. Plos One 8: e64721. 22. Pineda Torra I, Claudel T, Duval C, Kosykh V, Fruchart JC, et al. (2003) Bile acids induce the expression of the human peroxisome proliferator-activated receptor alpha gene via activation of the farnesoid X receptor. Molecular Endocrinology 17: 259-272. 23. Kast HR, Nguyen CM, Sinal CJ, Jones SA, Laffitte BA, et al. (2001) Farnesoid Xactivated receptor induces apolipoprotein C-II transcription: a molecular 118 mechanism linking plasma triglyceride levels to bile acids. Molecular Endocrinology 15: 1720-1728. 24. Jiang C, Xie C, Li F, Zhang L, Nichols RG, et al. (2015) Intestinal farnesoid X receptor signaling promotes nonalcoholic fatty liver disease. Journal of Clinical Investigation 125: 386-402. 25. Shimada A, Tamai T, Oida K, Takahashi S, Suzuki J, et al. (1994) Increase in neutral cholesteryl ester hydrolase activity produced by extralysosomal hydrolysis of high-density lipoprotein cholesteryl esters in rat hepatoma cells (H-35). Biochim Biophys Acta 1215: 126-132. 26. Hosokawa M, Furihata T, Yaginuma Y, Yamamoto N, Koyano N, et al. (2007) Genomic structure and transcriptional regulation of the rat, mouse, and human carboxylesterase genes. Drug Metab Rev 39: 1-15. 27. Hosokawa M (2008) Structure and catalytic properties of carboxylesterase isozymes involved in metabolic activation of prodrugs. Molecules 13: 412-431. 28. Geshi E, Kimura T, Yoshimura M, Suzuki H, Koba S, et al. (2005) A single nucleotide polymorphism in the carboxylesterase gene is associated with the responsiveness to imidapril medication and the promoter activity. Hypertension Research 28: 719-725. 29. Mori M, Hosokawa M, Ogasawara Y, Tsukada E, Chiba K (1999) cDNA cloning, characterization and stable expression of novel human brain carboxylesterase. Febs Letters 458: 17-22. 119 30. Takai S, Matsuda A, Usami Y, Adachi T, Sugiyama T, et al. (1997) Hydrolytic profile for ester- or amide-linkage by carboxylesterases pI 5.3 and 4.5 from human liver. Biol Pharm Bull 20: 869-873. 31. Danks MK, Morton CL, Pawlik CA, Potter PM (1998) Overexpression of a rabbit liver carboxylesterase sensitizes human tumor cells to CPT-11. Cancer Research 58: 20-22. 32. Guichard SM, Morton CL, Krull EJ, Stewart CF, Danks MK, et al. (1998) Conversion of the CPT-11 metabolite APC to SN-38 by rabbit liver carboxylesterase. Clinical Cancer Research 4: 3089-3094. 33. Humerickhouse R, Lohrbach K, Li L, Bosron WF, Dolan ME (2000) Characterization of CPT-11 hydrolysis by human liver carboxylesterase isoforms hCE-1 and hCE2. Cancer Research 60: 1189-1192. 34. Kojima A, Hackett NR, Ohwada A, Crystal RG (1998) In vivo human carboxylesterase cDNA gene transfer to activate the prodrug CPT-11 for local treatment of solid tumors. Journal of Clinical Investigation 101: 1789-1796. 35. Potter PM, Pawlik CA, Morton CL, Naeve CW, Danks MK (1998) Isolation and partial characterization of a cDNA encoding a rabbit liver carboxylesterase that activates the prodrug irinotecan (CPT-11). Cancer Research 58: 2646-2651. 36. Tabata T, Katoh M, Tokudome S, Nakajima M, Yokoi T (2004) Identification of the cytosolic carboxylesterase catalyzing the 5'-deoxy-5-fluorocytidine formation from capecitabine in human liver. Drug Metab Dispos 32: 1103-1110. 120 37. Brzezinski MR, Spink BJ, Dean RA, Berkman CE, Cashman JR, et al. (1997) Human liver carboxylesterase hCE-1: binding specificity for cocaine, heroin, and their metabolites and analogs. Drug Metab Dispos 25: 1089-1096. 38. Zhang J, Burnell JC, Dumaual N, Bosron WF (1999) Binding and hydrolysis of meperidine by human liver carboxylesterase hCE-1. Journal of Pharmacology and Experimental Therapeutics 290: 314-318. 39. Furihata T, Hosokawa M, Koyano N, Nakamura T, Satoh T, et al. (2004) Identification of di-(2-ethylhexyl) phthalate-induced carboxylesterase 1 in C57BL/6 mouse liver microsomes: purification, cDNA cloning, and baculovirusmediated expression. Drug Metab Dispos 32: 1170-1177. 40. Ishiguro N, Kishimoto W, Volz A, Ludwig-Schwellinger E, Ebner T, et al. (2014) Impact of endogenous esterase activity on in vitro p-glycoprotein profiling of dabigatran etexilate in Caco-2 monolayers. Drug Metab Dispos 42: 250-256. 41. Laizure SC, Parker RB, Herring VL, Hu ZY (2014) Identification of carboxylesterase-dependent dabigatran etexilate hydrolysis. Drug Metab Dispos 42: 201-206. 42. Kojima A, Hackett NR, Crystal RG (1998) Reversal of CPT-11 resistance of lung cancer cells by adenovirus-mediated gene transfer of the human carboxylesterase cDNA. Cancer Research 58: 4368-4374. 43. Senter PD, Marquardt H, Thomas BA, Hammock BD, Frank IS, et al. (1996) The role of rat serum carboxylesterase in the activation of paclitaxel and camptothecin prodrugs. Cancer Research 56: 1471-1474. 121 44. Wierdl M, Morton CL, Weeks JK, Danks MK, Harris LC, et al. (2001) Sensitization of human tumor cells to CPT-11 via adenoviral-mediated delivery of a rabbit liver carboxylesterase. Cancer Research 61: 5078-5082. 45. Hu ZY, Edginton AN, Laizure SC, Parker RB (2014) Physiologically based pharmacokinetic modeling of impaired carboxylesterase-1 activity: effects on oseltamivir disposition. Clin Pharmacokinet 53: 825-836. 46. Suzaki Y, Uemura N, Takada M, Ohyama T, Itohda A, et al. (2013) The effect of carboxylesterase 1 (CES1) polymorphisms on the pharmacokinetics of oseltamivir in humans. European Journal of Clinical Pharmacology 69: 21-30. 47. Zhu HJ, Markowitz JS (2013) Carboxylesterase 1 (CES1) genetic polymorphisms and oseltamivir activation. European Journal of Clinical Pharmacology 69: 733-734. 48. Zhu HJ, Appel DI, Johnson JA, Chavin KD, Markowitz JS (2009) Role of carboxylesterase 1 and impact of natural genetic variants on the hydrolysis of trandolapril. Biochemical Pharmacology 77: 1266-1272. 49. Ishizuka T, Yoshigae Y, Murayama N, Izumi T (2013) Different hydrolases involved in bioactivation of prodrug-type angiotensin receptor blockers: carboxymethylenebutenolidase and carboxylesterase 1. Drug Metab Dispos 41: 1888-1895. 50. Vistoli G, Pedretti A, Mazzolari A, Bolchi C, Testa B (2009) Influence of ionization state on the activation of temocapril by hCES1: a molecular-dynamics study. Chemistry & Biodiversity 6: 2092-2100. 51. Zhao B, Song JM, Chow WN, Clair RWS, Rudel LL, et al. (2007) Macrophagespecific transgenic expression of cholesteryl ester hydrolase significantly reduces 122 atherosclerosis and lesion necrosis in Ldlr(-)/(-) mice. Journal of Clinical Investigation 117: 2983-2992. 52. Ko KWS, Erickson B, Lehner R (2009) Es-x/Ces1 prevents triacylglycerol accumulation in McArdle-RH7777 hepatocytes. Biochimica Et Biophysica ActaMolecular and Cell Biology of Lipids 1791: 1133-1143. 53. Becker A, Bottcher A, Lackner KJ, Fehringer P, Notka F, et al. (1994) Purification, cloning, and expression of a human enzyme with acyl coenzyme A: cholesterol acyltransferase activity, which is identical to liver carboxylesterase. Arterioscler Thromb 14: 1346-1355. 54. Lian J, Wei E, Wang SP, Quiroga AD, Li L, et al. (2012) Liver specific inactivation of carboxylesterase 3/triacylglycerol hydrolase decreases blood lipids without causing severe steatosis in mice. Hepatology 56: 2154-2162. 55. Wei E, Ben Ali Y, Lyon J, Wang H, Nelson R, et al. (2010) Loss of TGH/Ces3 in mice decreases blood lipids, improves glucose tolerance, and increases energy expenditure. Cell Metab 11: 183-193. 56. Crow JA, Middleton BL, Borazjani A, Hatfield MJ, Potter PM, et al. (2008) Inhibition of carboxylesterase 1 is associated with cholesteryl ester retention in human THP-1 monocyte/macrophages. Biochim Biophys Acta 1781: 643-654. 57. Ghosh S, St Clair RW, Rudel LL (2003) Mobilization of cytoplasmic CE droplets by overexpression of human macrophage cholesteryl ester hydrolase. Journal of Lipid Research 44: 1833-1840. 123 58. Zhao B, Song J, St Clair RW, Ghosh S (2007) Stable overexpression of human macrophage cholesteryl ester hydrolase results in enhanced free cholesterol efflux from human THP1 macrophages. Am J Physiol Cell Physiol 292: C405-412. 59. Quiroga AD, Lian JH, Lehner R (2012) Carboxylesterase1/Esterase-x Regulates Chylomicron Production in Mice. Plos One 7. 60. Quiroga AD, Li L, Trotzmuller M, Nelson R, Proctor SD, et al. (2012) Deficiency of carboxylesterase 1/esterase-x results in obesity, hepatic steatosis, and hyperlipidemia. Hepatology 56: 2188-2198. 61. Bligh EG, Dyer WJ (1959) A rapid method of total lipid extraction and purification. Can J Biochem Physiol 37: 911-917. 62. Zhang Y, Lee FY, Barrera G, Lee H, Vales C, et al. (2006) Activation of the nuclear receptor FXR improves hyperglycemia and hyperlipidemia in diabetic mice. Proc Natl Acad Sci U S A 103: 1006-1011. 63. Ma L, Tsatsos NG, Towle HC (2005) Direct role of ChREBP.Mlx in regulating hepatic glucose-responsive genes. Journal of Biological Chemistry 280: 1201912027. 64. Wang Q, Li S, Jiang L, Zhou Y, Li Z, et al. (2010) Deficiency in hepatic ATP-citrate lyase affects VLDL-triglyceride mobilization and liver fatty acid composition in mice. Journal of Lipid Research 51: 2516-2526. 65. Mastrodonato M, Calamita G, Rossi R, Mentino D, Bonfrate L, et al. (2011) Altered distribution of caveolin-1 in early liver steatosis. European Journal of Clinical Investigation 41: 642-651. 124 66. Edwards M, Houseman L, Phillips IR, Shephard EA (2013) Isolation of mouse hepatocytes. Methods Mol Biol 987: 283-293. 67. Xu J, Yin L, Xu Y, Li Y, Zalzala M, et al. (2014) Hepatic carboxylesterase 1 is induced by glucose and regulates postprandial glucose levels. Plos One 9: e109663. 68. Ge X, Yin L, Ma H, Li T, Chiang JY, et al. (2011) Aldo-keto reductase 1B7 is a target gene of FXR and regulates lipid and glucose homeostasis. Journal of Lipid Research 52: 1561-1568. 69. Zhang Y, Yin L, Hillgartner FB (2003) SREBP-1 integrates the actions of thyroid hormone, insulin, cAMP, and medium-chain fatty acids on ACCalpha transcription in hepatocytes. Journal of Lipid Research 44: 356-368. 70. Yin L, Ma H, Ge X, Edwards PA, Zhang Y (2011) Hepatic hepatocyte nuclear factor 4alpha is essential for maintaining triglyceride and cholesterol homeostasis. Arterioscler Thromb Vasc Biol 31: 328-336. 71. Rune A, Osler ME, Fritz T, Zierath JR (2009) Regulation of skeletal muscle sucrose, non-fermenting 1/AMP-activated protein kinase-related kinase (SNARK) by metabolic stress and diabetes. Diabetologia 52: 2182-2189. 72. Ma L, Robinson LN, Towle HC (2006) ChREBP center dot Mlx is the principal mediator of glucose-induced gene expression in the liver. Journal of Biological Chemistry 281: 28721-28730. 73. Uyeda K, Repa JJ (2006) Carbohydrate response element binding protein, ChREBP, a transcription factor coupling hepatic glucose utilization and lipid synthesis. Cell Metabolism 4: 107-110. 125 74. Postic C, Dentin R, Denechaud PD, Girard J (2007) ChREBP, a transcriptional regulator of glucose and lipid metabolism. Annual Review of Nutrition 27: 179192. 75. Dentin R, Pegorier JP, Benhamed F, Foufelle F, Ferre P, et al. (2004) Hepatic glucokinase is required for the synergistic action of ChREBP and SREBP-1c on glycolytic and lipogenic gene expression. Journal of Biological Chemistry 279: 20314-20326. 76. Wellen KE, Hatzivassiliou G, Sachdeva UM, Bui TV, Cross JR, et al. (2009) ATPcitrate lyase links cellular metabolism to histone acetylation. Science 324: 10761080. 77. Li T, Chanda D, Zhang Y, Choi HS, Chiang JY (2010) Glucose stimulates cholesterol 7alpha-hydroxylase gene transcription in human hepatocytes. Journal of Lipid Research 51: 832-842. 78. El-Osta A, Brasacchio D, Yao D, Pocai A, Jones PL, et al. (2008) Transient high glucose causes persistent epigenetic changes and altered gene expression during subsequent normoglycemia (vol 205, pg 2409, 2008). Journal of Experimental Medicine 205: 2683-2683. 79. Brasacchio D, Okabe J, Tikellis C, Balcerczyk A, George P, et al. (2009) Hyperglycemia induces a dynamic cooperativity of histone methylase and demethylase enzymes associated with gene-activating epigenetic marks that coexist on the lysine tail. Diabetes 58: 1229-1236. 80. Villeneuve LM, Reddy MA, Lanting LL, Wang M, Meng L, et al. (2008) Epigenetic histone H3 lysine 9 methylation in metabolic memory and inflammatory 126 phenotype of vascular smooth muscle cells in diabetes. Proceedings of the National Academy of Sciences of the United States of America 105: 9047-9052. 81. Iynedjian PB, Jotterand D, Nouspikel T, Asfari M, Pilot PR (1989) Transcriptional induction of glucokinase gene by insulin in cultured liver cells and its repression by the glucagon-cAMP system. Journal of Biological Chemistry 264: 2182421829. 82. Sapiro JM, Mashek MT, Greenberg AS, Mashek DG (2009) Hepatic triacylglycerol hydrolysis regulates peroxisome proliferator-activated receptor alpha activity. Journal of Lipid Research 50: 1621-1629. 83. Brown MS, Goldstein JL (1999) A proteolytic pathway that controls the cholesterol content of membranes, cells, and blood. Proceedings of the National Academy of Sciences of the United States of America 96: 11041-11048. 84. Schmidt DR, Schmidt S, Holmstrom SR, Makishima M, Yu RT, et al. (2011) AKR1B7 is induced by the farnesoid X receptor and metabolizes bile acids. Journal of Biological Chemistry 286: 2425-2432. 85. Zhao B, Song J, Ghosh S (2008) Hepatic overexpression of cholesteryl ester hydrolase enhances cholesterol elimination and in vivo reverse cholesterol transport. Journal of Lipid Research 49: 2212-2217. 86. Zhang T, Zhao S, Li W, Ma L, Ding M, et al. (2014) High-fat diet from perilla oil induces insulin resistance despite lower serum lipids and increases hepatic fatty acid oxidation in rats. Lipids in Health and Disease 13: 15. 87. Rachek LI (2014) Free fatty acids and skeletal muscle insulin resistance. Prog Mol Biol Transl Sci 121: 267-292. 127 88. Xiong J, Shang W, Wu L, Chen R, Liu W, et al. (2014) Glucose dominates the regulation of carboxylesterases induced by lipopolysaccharide or interleukin-6 in primary mouse hepatocytes. Life Sciences 112: 41-48. 89. Wellen KE, Hatzivassiliou G, Sachdeva UM, Bui TV, Cross JR, et al. (2009) ATPCitrate Lyase Links Cellular Metabolism to Histone Acetylation. Science 324: 1076-1080. 90. Reid BN, Ables GP, Otlivanchik OA, Schoiswohl G, Zechner R, et al. (2008) Hepatic overexpression of hormone-sensitive lipase and adipose triglyceride lipase promotes fatty acid oxidation, stimulates direct release of free fatty acids, and ameliorates steatosis. Journal of Biological Chemistry 283: 13087-13099. 91. Adorini L, Pruzanski M, Shapiro D (2012) Farnesoid X receptor targeting to treat nonalcoholic steatohepatitis. Drug Discov Today 17: 988-997. 92. Gao B, Bataller R (2011) Alcoholic Liver Disease: Pathogenesis and New Therapeutic Targets. Gastroenterology 141: 1572-1585. 93. Zou C, Ma J, Wang X, Guo L, Zhu Z, et al. (2007) Lack of Fas antagonism by Met in human fatty liver disease. Nature Medicine 13: 1078-1085. 94. Ji C, Kaplowitz N (2006) ER stress: Can the liver cope? Journal of Hepatology 45: 321-333. 95. Lakshman AR (2004) Some novel insights into the pathogenesis of alcoholic steatosis. Alcohol 34: 45-48. 96. Orman ES, Odena G, Bataller R (2013) Alcoholic liver disease: pathogenesis, management, and novel targets for therapy. J Gastroenterol Hepatol 28 Suppl 1: 77-84. 128 97. Dey A, Cederbaum AI (2006) Alcohol and oxidative liver injury. Hepatology 43: S63-74. 98. Baraona E, Pirola RC, Lieber CS (1975) Acute and Chronic Effects of Ethanol on Intestinal Lipid-Metabolism. Biochimica Et Biophysica Acta 388: 19-28. 99. Yu ST, Rao S, Reddy JK (2003) Peroxisome proliferator-activated receptors, fatty acid oxidation, steatohepatitis and hepatocarcinogenesis. Current Molecular Medicine 3: 561-572. 100. Wagner M, Zollner G, Trauner M (2011) Nuclear Receptors in Liver Disease. Hepatology 53: 1023-1034. 101. Shen Z, Liang XM, Rogers CQ, Rideout D, You M (2010) Involvement of adiponectin-SIRT1-AMPK signaling in the protective action of rosiglitazone against alcoholic fatty liver in mice. American Journal of PhysiologyGastrointestinal and Liver Physiology 298: G364-G374. 102. Ajmo JM, Liang XM, Rogers CQ, Pennock B, You M (2008) Resveratrol alleviates alcoholic fatty liver in mice. American Journal of Physiology-Gastrointestinal and Liver Physiology 295: G833-G842. 103. You M, Liang XM, Ajmo JM, Ness GC (2008) Involvement of mammalian sirtuin 1 in the action of ethanol in the liver. American Journal of PhysiologyGastrointestinal and Liver Physiology 294: G892-G898. 104. You M, Matsumoto M, Pacold CM, Cho WK, Crabb DW (2004) The role of AMPactivated protein kinase in the action of ethanol in the liver. Gastroenterology 127: 1798-1808. 129 105. Canto C, Auwerx J (2009) PGC-1 alpha, SIRT1 and AMPK, an energy sensing network that controls energy expenditure. Current Opinion in Lipidology 20: 98105. 106. Canto C, Gerhart-Hines Z, Feige JN, Lagouge M, Noriega L, et al. (2009) AMPK regulates energy expenditure by modulating NAD(+) metabolism and SIRT1 activity. Nature 458: 1056-U1140. 107. Hritz I, Mandrekar P, Velayudham A, Catalano D, Dolganiuc A, et al. (2008) The critical role of Toll-like receptor (TLR) 4 in alcoholic liver disease is independent of the common TLR adapter MyD88. Hepatology 48: 1224-1231. 108. Wang HJ, Gao B, Zakhari S, Nagy LE (2012) Inflammation in Alcoholic Liver Disease. Annual Review of Nutrition, Vol 32 32: 343-+. 109. Shen Z, Ajmo JM, Rogers CQ, Liang XM, Le L, et al. (2009) Role of SIRT1 in regulation of LPS- or two ethanol metabolites-induced TNF-alpha production in cultured macrophage cell lines. American Journal of Physiology-Gastrointestinal and Liver Physiology 296: G1047-G1053. 110. Lee JY, Sohn KH, Rhee SH, Hwang D (2001) Saturated fatty acids, but not unsaturated fatty acids, induce the expression of cyclooxygenase-2 mediated through Toll-like receptor 4. Journal of Biological Chemistry 276: 16683-16689. 111. Lakshminarayanan V, Beno DW, Costa RH, Roebuck KA (1997) Differential regulation of interleukin-8 and intercellular adhesion molecule-1 by H2O2 and tumor necrosis factor-alpha in endothelial and epithelial cells. Journal of Biological Chemistry 272: 32910-32918. 130 112. Fleming CD, Bencharit S, Edwards CC, Hyatt JL, Tsurkan L, et al. (2005) Structural insights into drug processing by human carboxylesterase 1: Tamoxifen, mevastatin, and inhibition by benzil. Journal of Molecular Biology 352: 165-177. 113. Gonzalez FJ (2008) Regulation of hepatocyte nuclear factor 4 alpha-mediated transcription. Drug Metab Pharmacokinet 23: 2-7. 114. Bertola A, Mathews S, Ki SH, Wang H, Gao B (2013) Mouse model of chronic and binge ethanol feeding (the NIAAA model). Nature Protocols 8: 627-637. 115. Ge XM, Yin LY, Ma HY, Li TG, Chiang JYL, et al. (2011) Aldo-keto reductase 1B7 is a target gene of FXR and regulates lipid and glucose homeostasis. Journal of Lipid Research 52: 1561-1568. 116. Guan X, Rubin E, Anni H (2012) An optimized method for the measurement of acetaldehyde by high-performance liquid chromatography. Alcoholism-Clinical and Experimental Research 36: 398-405. 117. Mishin V, Gray JP, Heck DE, Laskin DL, Laskin JD (2010) Application of the Amplex red/horseradish peroxidase assay to measure hydrogen peroxide generation by recombinant microsomal enzymes. Free Radic Biol Med 48: 14851491. 118. Labonne BEF, Gutierrez M, Gomez-Quiroz LE, Fainstein MK, Bucio L, et al. (2009) Acetaldehyde-induced mitochondrial dysfunction sensitizes hepatocytes to oxidative damage. Cell Biology and Toxicology 25: 599-609. 119. Bulua AC, Simon A, Maddipati R, Pelletier M, Park H, et al. (2011) Mitochondrial reactive oxygen species promote production of proinflammatory cytokines and 131 are elevated in TNFR1-associated periodic syndrome (TRAPS). Journal of Experimental Medicine 208: 519-533. 120. Nakahira K, Haspel JA, Rathinam VA, Lee SJ, Dolinay T, et al. (2011) Autophagy proteins regulate innate immune responses by inhibiting the release of mitochondrial DNA mediated by the NALP3 inflammasome. Nature Immunology 12: 222-230. 121. Naik E, Dixit VM (2011) Mitochondrial reactive oxygen species drive proinflammatory cytokine production. Journal of Experimental Medicine 208: 417-420. 122. Shokolenko I, Venediktova N, Bochkareva A, Wilson GL, Alexeyev MF (2009) Oxidative stress induces degradation of mitochondrial DNA. Nucleic Acids Research 37: 2539-2548. 123. Babeu JP, Boudreau F (2014) Hepatocyte nuclear factor 4-alpha involvement in liver and intestinal inflammatory networks. World J Gastroenterol 20: 22-30. 124. Hertz R, Magenheim J, Berman I, Bar-Tana J (1998) Fatty acyl-CoA thioesters are ligands of hepatic nuclear factor-4alpha. Nature 392: 512-516. 125. Li Q, Zhong W, Qiu Y, Kang X, Sun X, et al. (2013) Preservation of hepatocyte nuclear factor-4alpha contributes to the beneficial effect of dietary medium chain triglyceride on alcohol-induced hepatic lipid dyshomeostasis in rats. AlcoholismClinical and Experimental Research 37: 587-598. 126. Xu Y, Zalzala M, Xu J, Li Y, Yin L, et al. (2015) A metabolic stress-inducible miR34a-HNF4alpha pathway regulates lipid and lipoprotein metabolism. Nature Communications 6: 7466. 132 127. Clugston RD, Jiang H, Lee MX, Piantedosi R, Yuen JJ, et al. (2011) Altered hepatic lipid metabolism in C57BL/6 mice fed alcohol: a targeted lipidomic and gene expression study. Journal of Lipid Research 52: 2021-2031. 128. Palanker L, Tennessen JM, Lam G, Thummel CS (2009) Drosophila HNF4 regulates lipid mobilization and beta-oxidation. Cell Metab 9: 228-239. 129. Yamaguchi K, Yang L, McCall S, Huang J, Yu XX, et al. (2007) Inhibiting triglyceride synthesis improves hepatic steatosis but exacerbates liver damage and fibrosis in obese mice with nonalcoholic steatohepatitis. Hepatology 45: 13661374. 130. Zhao B, Natarajan R, Ghosh S (2005) Human liver cholesteryl ester hydrolase: cloning, molecular characterization, and role in cellular cholesterol homeostasis. Physiological Genomics 23: 304-310. 131. McGuire LC, Cruickshank AM, Munro PT (2006) Alcoholic ketoacidosis. Emerg Med J 23: 417-420. 132. Tabas I, Williams KJ, Boren J (2007) Subendothelial lipoprotein retention as the initiating process in atherosclerosis - Update and therapeutic implications. Circulation 116: 1832-1844. 133. Libby P (2013) Mechanisms of Acute Coronary Syndromes and Their Implications for Therapy. New England Journal of Medicine 368: 2004-2013. 134. Bondjers G, Olofsson SO, Fager A, Wiklund O (1986) Atherosclerosis basic mechanisms. Acta Med Scand Suppl 712: 10-18. 135. Libby P (2002) Inflammation in atherosclerosis. Nature 420: 868-874. 133 136. Demer LL, Watson KE, Bostrom K (1994) Mechanism of calcification in atherosclerosis. Trends Cardiovasc Med 4: 45-49. 137. Rosenson RS, Brewer HB, Jr., Davidson WS, Fayad ZA, Fuster V, et al. (2012) Cholesterol efflux and atheroprotection: advancing the concept of reverse cholesterol transport. Circulation 125: 1905-1919. 138. Rader DJ (2006) Molecular regulation of HDL metabolism and function: implications for novel therapies. Journal of Clinical Investigation 116: 3090-3100. 139. Bie JH, Wang J, Yuan Q, Kakiyama G, Ghosh SS, et al. (2014) Liver-specific transgenic expression of cholesteryl ester hydrolase reduces atherosclerosis in Ldlr(-/-) mice. Journal of Lipid Research 55: 729-738. 140. Zhang Y, Wang X, Vales C, Lee FY, Lee H, et al. (2006) FXR deficiency causes reduced atherosclerosis in Ldlr-/- mice. Arterioscler Thromb Vasc Biol 26: 23162321. 141. Paigen B, Morrow A, Holmes PA, Mitchell D, Williams RA (1987) Quantitative assessment of atherosclerotic lesions in mice. Atherosclerosis 68: 231-240. 142. Bai B, Liang Y, Xu C, Lee MY, Xu A, et al. (2012) Cyclin-dependent kinase 5mediated hyperphosphorylation of sirtuin-1 contributes to the development of endothelial senescence and atherosclerosis. Circulation 126: 729-740. 143. Ray A, Dittel BN (2010) Isolation of mouse peritoneal cavity cells. J Vis Exp. 134