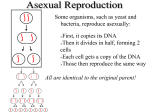
Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
15 Oct 2004 19:50 AR AR230-GE38-08.tex AR230-GE38-08.sgm P1: GCE LaTeX2e(2002/01/18) 10.1146/annurev.genet.38.072902.091500 Annu. Rev. Genet. 2004. 38:233–71 doi: 10.1146/annurev.genet.38.072902.091500 c 2004 by Annual Reviews. All rights reserved Copyright First published online as a Review in Advance on June 21, 2004 RECOMBINATION PROTEINS IN YEAST Berit Olsen Krogh and Lorraine S. Symington Annu. Rev. Genet. 2004.38:233-271. Downloaded from arjournals.annualreviews.org by UNIVERSITY OF SOUTH CAROL on 02/10/05. For personal use only. Columbia University Medical Center, New York, New York 10032; email: [email protected]; [email protected] Key Words double-strand break repair, RAD52 epistasis group, Holliday junction resolution, replication fork collapse ■ Abstract The process of homologous recombination promotes error-free repair of double-strand breaks and is essential for meiosis. Central to the process of homologous recombination are the RAD52 group genes (RAD50, RAD51, RAD52, RAD54, RDH54/TID1, RAD55, RAD57, RAD59, MRE11, and XRS2), most of which were identified by their requirement for the repair of ionizing radiation-induced DNA damage in Saccharomyces cerevisiae. The Rad52 group proteins are highly conserved among eukaryotes. Recent studies showing defects in homologous recombination and doublestrand break repair in several human cancer-prone syndromes have emphasized the importance of this repair pathway in maintaining genome integrity. Herein, we review recent genetic, biochemical, and structural analyses of the genes and proteins involved in recombination. CONTENTS INTRODUCTION . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . SUBSTRATES AND MECHANISMS OF HOMOLOGOUS RECOMBINATION . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . The DSBR and SDSA Models . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . One-Ended Strand Invasion . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Single-Strand Annealing . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Recombination at Stalled or Collapsed Replication Forks . . . . . . . . . . . . . . . . . . . . DSB FORMATION . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Initiation of Meiotic Recombination . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Initiation of Mitotic Recombination . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . FORMATION OF 3 ssDNA TAILS AT BREAK SITES . . . . . . . . . . . . . . . . . . . . . . . Properties of the MRX Complex . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Resection of DSBs . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . FORMATION OF THE RAD51 NUCLEOPROTEIN FILAMENT . . . . . . . . . . . . . . Properties of Rad51 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Mediators of Rad51 Filament Assembly . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 0066-4197/04/1215-0233$14.00 234 235 235 235 236 237 239 239 240 240 240 243 246 246 248 233 15 Oct 2004 19:50 234 AR KROGH AR230-GE38-08.tex AR230-GE38-08.sgm LaTeX2e(2002/01/18) P1: GCE SYMINGTON Annu. Rev. Genet. 2004.38:233-271. Downloaded from arjournals.annualreviews.org by UNIVERSITY OF SOUTH CAROL on 02/10/05. For personal use only. Homologous Pairing and Strand Exchange . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Negative Regulation of Recombination by Srs2 . . . . . . . . . . . . . . . . . . . . . . . . . . . . RAD51-INDEPENDENT RECOMBINATION . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Rad52: a DNA Annealing Protein . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Dmc1 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . HOLLIDAY JUNCTION RESOLUTION ACTIVITIES . . . . . . . . . . . . . . . . . . . . . . . The Mus81-Mms4 (Eme1) Nuclease . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Resolvase A . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Dissolution of dHJs . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . NONHOMOLOGOUS END JOINING . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 249 250 250 250 252 253 253 255 255 256 INTRODUCTION Recombination refers to the exchange, or transfer, of information between DNA molecules and is found in all organisms that have been studied in detail. Homologous recombination (HR) involves the interaction of DNA sequences with perfect, or near-perfect, homology over several hundreds of base pairs. Recombination between sequences with little or no sequence homology is referred to as nonhomologous recombination or end joining (NHEJ). The process of homologous recombination plays essential roles in the maintenance of genome stability in prokaryotic and eukaryotic organisms. The primary function of homologous recombination in mitotic cells is to repair double-strand breaks (DSBs) or singlestrand gaps that form as a result of replication fork collapse, from processing of spontaneous damage, and from exposure to DNA-damaging agents. Homologous recombination is also required for telomere maintenance, and consequently, proliferation, in the absence of telomerase. During meiosis, HR is essential to establish a physical connection between homologous chromosomes to ensure their correct disjunction at the first meiotic division. In addition, the high frequency of meiotic recombination contributes to diversity by creating new linkage arrangements between genes, or parts of genes. Yeast has proved a valuable system for the analysis of recombination in eukaryotes. Much of our understanding of the mechanisms of recombination is based on organisms, such as Saccharomyces cerevisiae, in which all of the products of an individual meiosis can be recovered for analysis in the form of asci containing four haploid spores. The analysis of the segregation of heterozygous markers during meiosis forms the basis for most recombination models. The ability to manipulate the genome to create well-defined genetic and physical markers has also contributed to our understanding of the mechanisms of HR. Genetic screens for mutants with altered rates of recombination, decreased survival in response to DNA-damaging agents, reduced sporulation, or spore viability have led to the identification of more than 30 genes involved in recombination. The proteins encoded by the RAD52 epistasis group genes [RAD50, RAD51, RAD52, RAD54, RAD55, RAD57, RAD59, RDH54 (TID1), MRE11 (RAD58), and XRS2] play direct roles in HR and are the focus of this review. 15 Oct 2004 19:50 AR AR230-GE38-08.tex AR230-GE38-08.sgm LaTeX2e(2002/01/18) RECOMBINATION PROTEINS IN YEAST P1: GCE 235 Annu. Rev. Genet. 2004.38:233-271. Downloaded from arjournals.annualreviews.org by UNIVERSITY OF SOUTH CAROL on 02/10/05. For personal use only. SUBSTRATES AND MECHANISMS OF HOMOLOGOUS RECOMBINATION Two types of recombination events have been identified based on the segregation of heterozygous markers during meiosis: crossing over and gene conversion. Studies in yeast and other fungi have shown that meiotic gene conversion events are frequently associated with the exchange of flanking markers (79). This association was thought to be due to alternate resolution of a Holliday junction (HJ) containing intermediate to generate crossover or noncrossover products. However, recent studies suggest crossover and noncrossover products are generated from different intermediates in meiotic recombination (7). Several models have been proposed to explain the molecular mechanisms of recombination (52, 72, 125, 139, 203), and of these the double-strand-break repair (DSBR) and synthesis-dependent strandannealing (SDSA) models are most consistent with the available genetic data. The DSBR and SDSA Models Studies of DSB-induced recombination between plasmid and chromosomal sequences provided the impetus for the DSBR model (144, 145, 203), and subsequent meiotic studies were in good agreement with its predictions (30, 124, 168, 196). In this model the initiating event is a DSB (Figure 1). The ends are resected to form 3 single-stranded (ss) tails that are active in strand invasion with a homologous duplex. Following strand invasion, the 3 end is extended by DNA synthesis. The D-loop formed by strand invasion is able to pair with the other side of the DSB (second end capture), and the 3 end of the noninvading strand is also extended by DNA synthesis. Gap filling and ligation results in the formation of a double Holliday junction (dHJ) intermediate. Random resolution of the two Holliday junctions is expected to yield equal numbers of crossover and noncrossover products. The DSBR model adequately explains several features of meiotic recombination. To account for the low levels of associated crossing over observed for mitotic DSB-induced gene conversion events, the synthesis-dependent strand-annealing (SDSA) model was proposed (52, 139, 189) (Figure 1). In the SDSA model, one or both ends of the broken duplex invade the donor and initiate DNA synthesis as envisioned in the DSBR model. For two-ended invasions, both strands are subsequently displaced and able to anneal through the newly synthesized complementary regions (139). For a one-ended invasion, the elongated invading strand is displaced after extensive DNA synthesis and pairs with the other side of the break (52). One-Ended Strand Invasion Under some circumstances, a broken chromosome may present only one end for repair (Figure 2). Collapsed replication forks generate only one end for strand invasion to restart replication. A one-ended DSB will also be generated when a telomere becomes uncapped and is degraded (62, 66, 113). The chromosome 15 Oct 2004 19:50 Annu. Rev. Genet. 2004.38:233-271. Downloaded from arjournals.annualreviews.org by UNIVERSITY OF SOUTH CAROL on 02/10/05. For personal use only. 236 AR KROGH AR230-GE38-08.tex AR230-GE38-08.sgm LaTeX2e(2002/01/18) P1: GCE SYMINGTON Figure 1 Models for the repair of double-strand breaks. The DSBR and SDSA models both initiate with invasion of a 3 end (left panel). After priming DNA synthesis, the second end is captured and a double Holliday junction intermediate is formed (DSBR). Resolution can occur in either plane at both junctions to generate crossover or noncrossover products. In the SDSA model, the nascent strand is displaced, pairs with the other 3 single-stranded tail, and DNA synthesis completes repair. A DSB made between direct repeats is subject to resection to generate 3 single-stranded tails (right panel). When complementary sequences are revealed owing to extensive resection, the single-stranded DNA anneals resulting in deletion of one of the repeats and the intervening DNA. The 3 tails are removed by the Rad1/10 nuclease and the nicks are ligated. 3 ends are indicated by arrowheads, newly synthesized DNA is represented by dashed lines. end can then invade homologous sequences (sister chromatid or homologue, or subtelomeric repeats of other chromosomes) and initiate DNA synthesis from the site of strand invasion to the telomere, templated by the donor duplex. This process is termed break-induced replication (BIR). Homology-dependent duplication of an entire chromosome arm to the end of a transformed linearized plasmid has also been detected in yeast and is presumed to occur by a similar mechanism (43, 134). BIR can occur at a conventional DSB if the gene conversion pathway is restricted by mutation of RAD51, or by limiting homology to one side of the break (99, 116). Single-Strand Annealing Single-strand annealing (SSA) is restricted to DSBs that occur between direct repeats (Figure 1). These events have been detected in yeast and animal cells using artificial direct repeats and could conceivably be important for repair in genomes of higher eukaryotes that contain many repeated sequences (54, 107, 119). After formation of the DSB, the ends are resected to produce 3 single-stranded tails, 15 Oct 2004 19:50 AR AR230-GE38-08.tex AR230-GE38-08.sgm LaTeX2e(2002/01/18) Annu. Rev. Genet. 2004.38:233-271. Downloaded from arjournals.annualreviews.org by UNIVERSITY OF SOUTH CAROL on 02/10/05. For personal use only. RECOMBINATION PROTEINS IN YEAST P1: GCE 237 Figure 2 One-ended strand invasion. If a telomere becomes uncapped, or the other side of a DSB is not available for HR, strand invasion can occur and replication proceeds to the end of the chromosome (BIR). which can anneal together when resection is sufficient to reveal complementary single-stranded regions. Single-stranded tails are removed by nucleases and the resulting gaps/nicks filled in by DNA repair synthesis and ligation. This process is accompanied by deletion of one of the repeats and the DNA between the direct repeats and is therefore considered to be mutagenic. Recombination at Stalled or Collapsed Replication Forks In yeast, HR efficiently repairs DSBs, but it is not clear if all mitotic recombination events initiate from DSBs. Single-stranded gaps at stalled replication forks might also initiate recombination and may occur more frequently than DSBs during vegetative growth of budding yeast (50). In Escherichia coli, generation of DSBs at stalled replication forks is thought to occur about once every two to three rounds of replication (126). In vertebrates, DSBs are thought to occur during every S-phase, explaining the essential requirement for recombination factors such as RAD51 (183, 214). There are several possible mechanisms for the formation of DSBs during replication. First, the replication fork could run into a nick on the template strand, resulting in collapse of the replication fork. Assuming that the nascent 15 Oct 2004 19:50 Annu. Rev. Genet. 2004.38:233-271. Downloaded from arjournals.annualreviews.org by UNIVERSITY OF SOUTH CAROL on 02/10/05. For personal use only. 238 AR KROGH AR230-GE38-08.tex AR230-GE38-08.sgm LaTeX2e(2002/01/18) P1: GCE SYMINGTON Figure 3 Role of recombination in restoring collapsed or stalled replication forks. DSBs made by replication fork collapse, or resolution of a regressed fork, are repaired by strand invasion restoring the fork. A gap on the lagging strand can be used to pair with the homologous sister chromatid to allow bypass of the lesion. strand of the other arm becomes ligated to the template strand of the same polarity, the broken arm could invade this duplex to restart replication (Figure 3). Second, the nascent strands at a stalled fork could pair and reverse branch migrate to form a pseudo-Holliday structure, also known as a regressed fork or chicken foot (71). This intermediate could then be cleaved by an HJ-specific endonuclease, resulting in collapse of the fork (170). The collapsed fork would then have to undergo strand invasion as described above. There is genetic evidence for regressed forks in E. coli (170), but in yeast, regressed forks have only been observed in hydroxyurea (HU)-arrested cells in the absence of the S-phase checkpoint kinase Rad53 (111). If replication stalls because of a lesion present on the leading strand, the fork could regress such that the lagging strand of the paired nascent strands templates synthesis of the leading strand (71). The regressed fork could then be resolved by branch migration to restart replication without strand invasion (Figure 3). Recent studies in E. coli have shown continued synthesis of the lagging strand when the leading strand is stalled, providing further support for this template-switching model (147). Template switching may also function in the repair of polymeraseblocking lesions on the lagging strand. In this case, the single-stranded region could pair with the parental strand of the sister chromatid and the newly synthesized leading strand could template synthesis of the stalled lagging strand in order to bypass the lesion (Figure 3) (50). The template-switching models do not involve a DSB intermediate, but are likely to require HR proteins for annealing and branch migration. 15 Oct 2004 19:50 AR AR230-GE38-08.tex AR230-GE38-08.sgm LaTeX2e(2002/01/18) RECOMBINATION PROTEINS IN YEAST P1: GCE 239 The above models share a number of common features, including formation of single-stranded DNA at break sites (or gaps), strand invasion or strand annealing, and resolution of recombination intermediates. Below, we review genetic, molecular, and biochemical studies of the yeast proteins thought to catalyze these steps of the homologous recombination reaction. DSB FORMATION Annu. Rev. Genet. 2004.38:233-271. Downloaded from arjournals.annualreviews.org by UNIVERSITY OF SOUTH CAROL on 02/10/05. For personal use only. Initiation of Meiotic Recombination During meiosis, the Spo11 protein catalyzes DSB formation at 150–200 sites within the genome to initiate high frequency of recombination (94). Spo11 is a meiosis-specific protein with sequence similarity to the small subunit (Top6A) of a type IIB topoisomerase from Sulfolobus shibatae (18, 95). Classical type II topoisomerases transiently break both strands of duplex DNA by formation of covalent 5 phosphotyrosyl enzyme-DNA intermediates. Following substrate topoisomerization, the covalent linkage is resolved and the DNA strands resealed. Spo11 appears to catalyze only the first of these two steps and in certain mutants (rad50S, sae2, mre11-S, and mre11-58) remains stably covalently joined to the 5 ends at break sites (5, 95, 123, 138, 158, 209). Tyr-135 is the active site tyrosine in Spo11 and accordingly, mutation of this residue results in the inability to induce DSBs in vivo (18, 48). The currently favored model is that Spo11 is removed from ends by the Mre11-Rad50-Xrs2 (MRX) complex and Sae2, and the resulting free 5 ends are then available for resection. Spo11 cleaves nonrandomly, but does not exhibit sequence-specific cleavage. The sites of DSBs coincide with DNase I hypersensitive sites in chromatin and are generally in the 5 noncoding regions of genes (17, 65, 232). Nine other genes, MEI4, MER2/REC107, REC102, REC103/SKI8, REC104, REC114, MRE11, RAD50, and XRS2, as well as two meiosis-specific RNA splicing factors (Mer1 and Mre2), are required for DSB formation during meiosis (94). MRE11, RAD50, and XRS2 are members of the RAD52 epistasis group, and are required for the repair of ionizing radiation (IR)-induced DNA damage in mitotic cells in addition to meiotic DSB formation in S. cerevisiae (4, 202). Mre11 transiently associates with sequences within chromatin that are the sites of Spo11dependent DSBs (23). This association, however, is still observed in the spo11Y135F active site mutant, indicating that Mre11 localization is not dependent on the presence of DSBs. Mre11 remains associated with DSB sites in strains that are defective for resection, but still dissociates from chromatin in strains defective for strand invasion, indicating that it is resection, not repair, that is the signal for removal of Mre11. The requirement for the MRX complex in DSB formation may be to ensure that the factors required for processing of Spo11-induced DSBs are present at the time of cleavage. The role of the other genes required for DSB formation is poorly understood. SKI8 is the only other gene, in addition to MRE11, RAD50, and XRS2, that is 15 Oct 2004 19:50 240 AR KROGH AR230-GE38-08.tex AR230-GE38-08.sgm LaTeX2e(2002/01/18) P1: GCE SYMINGTON expressed in vegetative cells and because of its well characterized role in RNA metabolism was thought to function indirectly during meiosis (94). However, Arora et al. (10) have shown that Ski8 and Spo11 directly interact and that both are required to recruit other proteins to chromatin, implicating a scaffold function for Ski8 in assembly of the pre-DSB complex (10). Annu. Rev. Genet. 2004.38:233-271. Downloaded from arjournals.annualreviews.org by UNIVERSITY OF SOUTH CAROL on 02/10/05. For personal use only. Initiation of Mitotic Recombination Mitotic recombination is initiated by accidental DSBs, single-strand gaps, and by the induction of specialized endonucleases. The best characterized of these endonucleases are HO and I-SceI, which have long nonpalindromic recognition sequences and generate 4-bp staggered cuts with 3 -OH overhangs (36, 98). HO endonuclease cleaves a 24-bp sequence within the mating-type locus MAT, to initiate gene conversion between the resident MAT allele and one of the two transcriptionally silent HM cassettes (141, 189). The HO endonuclease is normally expressed at the end of G1 in haploid mother cells, and the protein is highly unstable owing to the presence of a PEST signal that signals degradation via the ubiquitin –26S proteosome system (92). The DSB made by HO is repaired using the same factors required to repair IR-induced DSBs (see below). The mitochondrial endonuclease I-SceI is encoded by the optional intron of the 21S rRNA gene and is responsible for intron mobility (36). I-SceI recognizes and cleaves an 18-bp nonsymmetrical sequence within omega− DNA and the omega+ intron is used to template repair resulting in propagation of the intron. The recombination factors within mitochondria that promote repair of the I-SceI-induced break are unknown. When expressed in the nucleus, I-SceI cleaves its target site and subsequent repair occurs by the same mechanisms as in other chromosomal DSBs (156). FORMATION OF 3 ssDNA TAILS AT BREAK SITES Properties of the MRX Complex Repair of DNA double-strand breaks (DSBs) by homologous recombination requires nucleolytic processing of the DNA ends to form invasive 3 ssDNA tails. In S. cerevisiae, 3 ssDNA regions can be detected that extend hundreds to a few thousand nucleotides into the sequences flanking meiotic and mitotic DSBs (197, 225, 227). The initial observation that unresected meiotic DSBs accumulate in rad50S mutants suggested a role for Rad50 and its complex partners Mre11 and Xrs2 (MRX complex) in processing DSBs during meiosis. Yeast rad50, mre11 and xrs2 null mutants are also extremely sensitive to DNA-damaging agents such as IR and methyl methane-sulfonate (MMS) (28, 59, 88, 209). Moreover, the null mutants have slowed resection of HO-induced DSBs at the MAT locus (83, 191, 209). These observations suggested a common role for MRX in processing both meiotic and mitotic DSBs. 15 Oct 2004 19:50 AR AR230-GE38-08.tex AR230-GE38-08.sgm LaTeX2e(2002/01/18) Annu. Rev. Genet. 2004.38:233-271. Downloaded from arjournals.annualreviews.org by UNIVERSITY OF SOUTH CAROL on 02/10/05. For personal use only. RECOMBINATION PROTEINS IN YEAST P1: GCE 241 Structural and functional homologues of the Mre11/Rad50 complex exist in all domains of life (9). In E. coli, the Rad50/Mre11 homologues SbcC/SbcD form a large complex with ATP-dependent dsDNA 3 -5 exonuclease and ATPindependent ssDNA endonuclease activities (38, 171). Consistent with the homology to the nuclease subunit SbcD, human and yeast Mre11 have manganesedependent nuclease activities in vitro (149, 207, 208). The nucleolytic repertoire of Mre11 includes: (a) 3 -5 exonuclease activity on blunt and 3 recessed ends; (b) endonuclease activity on circular and linear ssDNA, but not on homopolymers; and (c) endonuclease cleavage of hairpin ends and 3 ssDNA overhangs at the single-/double-stranded transition. The observation that ssDNA homopolymers are resistant to cleavage suggests that Mre11 is structure specific and cleaves ssDNA by trapping transient secondary structures that allow the enzyme to cut at the preferred single-/double-stranded transition. Rad50 and Xrs2 both influence MRX substrate binding and potentiate the intrinsic nuclease activity of Mre11 (206, 207). The N-terminal region of the 78-kDa Mre11 protein contains five conserved phosphoesterase motifs found in a superfamily of phosphoesterases that includes di-metal Ser/Thr protein phosphatases and E. coli SbcD (88, 171). The crystal structure of Pyrococcus furiosus (Pf) Mre11 (res. 1–342) shows a protein with a two-domain fold. Domain I encompasses the Mre11 active site with seven residues from the conserved phosphoesterase motifs coordinating two Mn2+ ions. Domain II forms a cap structure that partially overhangs the active site (76). S. cerevisiae (Sc) Rad50 is a 153-kDa protein with homology to E. coli SbcC (5, 162, 171). Both proteins share the modular sequence architecture found in the structural maintenance of chromosomes (SMC) family of proteins (190). Common features include N-terminal Walker A and C-terminal Walker B motifs, linked by a 600–900 residue α-helical coiled-coil domain (9). The crystal structure of PfRad50 catalytic domain (cd) (res. 1–152 & 735–882) reveals that the Walker A and B domains assemble into a single globular intramolecular ATP-binding cassette (ABC) type ATPase domain. The α-helical region linking the two parts of the composite ATPase domain folds up as a protruding antiparallel coiled-coil (Figure 4) (75, 76, 78). In ScRad50 each coiled-coil extends approximately 60 nm away from the globular ATPase domain (8). PfRad50 forms homodimers in the presence of ATP (75, 78). Two nucleotides are bound per dimer, each specifically coordinated in cis by the Walker A motif of one monomer and in trans by the ABC signature motif of the other monomer, thereby linking ATP binding with dimer formation (129). Dimerization creates a DNA-binding interface across the Rad50 dimer that could explain the ATP stimulated DNA binding observed for both yeast and P. furiosus Rad50 (77, 162). Mre11 binds to the base of the coiled-coil near the Rad50 DNA-binding interface, suggesting the formation of a composite DNA-binding site within the (Mre11)2/(Rad50) 2 heterotetramer. The model MR-tetramer has two protruding coiled-coil tails pointing away from a globular head containing an Mre11 dimer bound to the Rad50 ABC-ATPase dimer (8, 75, 76). A conserved Cys-X-X-Cys motif at the apex of the coiled-coil forms a molecular zinc-hook, which allows Rad50 to form intermolecular dimers across the coiled-coil domains 15 Oct 2004 19:50 Annu. Rev. Genet. 2004.38:233-271. Downloaded from arjournals.annualreviews.org by UNIVERSITY OF SOUTH CAROL on 02/10/05. For personal use only. 242 AR KROGH AR230-GE38-08.tex AR230-GE38-08.sgm LaTeX2e(2002/01/18) P1: GCE SYMINGTON (75). Each Rad50 coiled-coil contributes two cysteines from the CXXC motif to form an interlocking metal binding site with one tetrahedrally coordinated Zn2+ ion. The overall structure of the MR complex and intermolecular dimers/multimers formed through the Rad50 zinc-hook have been observed by electron microscopy (EM) (8, 75) and scanning force microscopy (SFM) (44, 219). SFM analyses have also revealed that the HsMR and ScMRX complexes associate with ends of dsDNA molecules and through intermolecular interactions between the extended coiled-coils and zinc-hooks can tether linear DNA molecules together (34, 44, 219). Juxtaposition of DNA ends may be one of the important functions of MRX in NHEJ. The flexibility of the Rad50 coiled-coil and the overall length (120 nm) of a coiled-coil dimer suggests that the MRX complex may in fact be able to connect sister chromatids during HR (Figure 4). Mutations in the ScRad50 C687YLC690 Figure 4 Models of the Mre112Rad502Xrs21 pentamer and MRX DNA tethering functions. The terminal CXXC-hooks form an interlocking dimerization interface in the Rad50 coiled-coils. Intermolecular dimerization between individual complexes allows MRX to tether DNA ends (bottom) and perhaps tether and align sister chromatids (top). 15 Oct 2004 19:50 AR AR230-GE38-08.tex AR230-GE38-08.sgm LaTeX2e(2002/01/18) Annu. Rev. Genet. 2004.38:233-271. Downloaded from arjournals.annualreviews.org by UNIVERSITY OF SOUTH CAROL on 02/10/05. For personal use only. RECOMBINATION PROTEINS IN YEAST P1: GCE 243 motif result in defective MRX complex formation or increased IR sensitivity, underscoring the structural and functional importance of the zinc-hook (75). The third complex component, Xrs2 in yeast (MRX) or Nbs1 in vertebrates (MRN), is found only in eukaryotes (31, 88, 215, 220). Xrs2 is part of the complex through direct interaction with Mre11. Sedimentation analyses and size-exclusion chromatography suggest a M2R2X1 complex stoichiometry (8, 34, 215). Although they are thought to be functional analogues, the sequence similarity between Xrs2 and Nbs1 is limited to an N-terminal forkhead-associated (FHA) domain. The FHA domain is a phosphorylation-dependent protein-protein interaction motif found in a variety of eukaryotic proteins. The HsMRN and ScMRX complexes play critical roles in initiation of the intra S-phase checkpoint in response to DNA damage, and Nbs1/Xrs2 are both phosphorylated as part of the damage signal by kinases from the ATM family of protein kinases (41, 106, 214a, 233, 236). The checkpoint function in combination with the DNA tethering and nuclease activities is likely to contribute to the complexity of phenotypes displayed by mrx mutants. Resection of DSBs The predicted nucleolytic activity needed for processing DSBs into 3 ssDNA ends is a 5 -3 exonuclease. Mre11 is a 3 -5 exonuclease, which leaves a directionality conflict inherently at odds with the involvement of Mre11 in DSB resection. However, point mutations in the conserved phosphoesterase motifs [D16A, D56N, H125N, and H213Y (mre11-58S)], which abolish Mre11-nuclease activity in vitro, result in accumulation of unresected meiotic DSBs and sporulation failure in vivo (57, 131, 209). The phenotype caused by these mre11-nuclease defective (nd) alleles is similar to that found for the rad50S mutants, which accumulate unresected DSBs with the meiotic Spo11-transesterase still covalently attached to the 5 DNA end (4, 95, 197). Similar biochemical analyses have not been repeated for the mre11-nd mutants, but based on the similarity in DSB accumulation and lack of resection, it is likely that these breaks also contain the covalent Spo11DNA adduct. The directionality paradox notwithstanding, these observations are evidence of a direct role for the MRX complex and Mre11-nuclease activity in meiotic DSB processing and Spo11 removal. Despite the severe meiotic phenotypes associated with mre11-nd alleles, the majority of these nuclease mutants show only mild-to-intermediate sensitivity to IR, and mating-type switching and resection of HO-induced DSBs occur with wild-type kinetics (29, 57, 103, 131). Moreover, the rad50, mre11 and xrs2 null mutations slow resection of HO-induced DSBs but do not abolish processing (83). Two alleles of mre11 show sensitivity to IR and MMS comparable to the null allele, mre11-4 (H241L,H243Y) and mre11-58S (H213Y). Both mutants, however, fail to interact with Rad50 in two-hybrid or coimmunoprecipitation assays, implying that for mitotic DSB resection, MRX as a complex is more important than the catalytic activity of Mre11 (29, 209, 215). To reconcile the conflicts surrounding the roles of MRX in meiotic versus mitotic DSB processing, we suggest the following resection models (Figure 5). To 15 Oct 2004 19:50 Annu. Rev. Genet. 2004.38:233-271. Downloaded from arjournals.annualreviews.org by UNIVERSITY OF SOUTH CAROL on 02/10/05. For personal use only. 244 AR KROGH AR230-GE38-08.tex AR230-GE38-08.sgm LaTeX2e(2002/01/18) P1: GCE SYMINGTON Figure 5 Model for the roles of the MRX complex in processing of DSBs. For simplicity, only one side of the DSB is included in this illustration. Resection of DSBs with modified/blocked ends is initiated by MRX (center). The “clean” ends are then substrate for continued processing by MRX (center) or by a 5 -3 exonuclease, e.g., Exo1 (right). Unmodified DSB ends can be resected directly by a 5 -3 exonuclease, independent of Mre11-mediated initiation (right) or through DNA unwinding by an unknown helicase and cleavage by the Mre11 structure-specific endonuclease (center and left). solve the nuclease directionality problem, MRX could potentially cooperate with a helicase to unwind the DNA duplex from the break and Mre11 processing the 5 strand by continuous endonucleolytic cleavage at transient secondary structures. For meiotic DSBs, resection of the 5 strand would be intrinsic to removal of Spo11, because the protein is released by the nucleolytic reaction most proximal to the DSB. It is also possible that Mre11 only acts as the initiating nuclease, 15 Oct 2004 19:50 AR AR230-GE38-08.tex AR230-GE38-08.sgm LaTeX2e(2002/01/18) Annu. Rev. Genet. 2004.38:233-271. Downloaded from arjournals.annualreviews.org by UNIVERSITY OF SOUTH CAROL on 02/10/05. For personal use only. RECOMBINATION PROTEINS IN YEAST P1: GCE 245 responsible for removing only a short stretch of DNA attached to the covalent DNA-(5 -phosphotyrosyl)-Spo11 adduct. Once the terminal adduct is removed, a 5 -3 exonuclease could take over and continue resection. A weak ATP-dependent DNA-unwinding activity has been associated with the human Rad50/Mre11/Nbs1 complex (150). Perhaps this weak helicase activity is sufficient to unwind a short stretch of DNA duplex from the Spo11 break site, enough for Mre11 to release Spo11 by the structure-specific endonuclease activity. We propose a model for mitotic DSB resection similar to the model for Mre11processing of Spo11-attached meiotic DSBs in which the Mre11-nuclease is specialized to deal with DSBs carrying modified ends that other nucleases are unable to process. Ionizing radiation as a source of DNA damage often generates DSBs with modified terminal nucleotides or even protein-DNA adducts. In our model, these modified DSBs require Mre11 for initial resection or “clean-up.” In contrast, HO-induced DSBs and other mitotic DSBs with unmodified clean ends would be direct substrates for processing by a bona fide resection nuclease or other redundant nucleases, even without Mre11 activity. Exo1, a 5 -3 dsDNA exonuclease and flap-endonuclease, is the only nuclease with an identified role in DSB resection in the absence of MRX (53). The exo1mre11 double mutant has severe growth defects and increased IR sensitivity compared with mre11 (210). When expressed from a high-copy-number plasmid, EXO1 partially rescues the mitotic DNA repair defects of mre11 strains, indicating that Exo1 is capable of some DSB processing in the absence of MRX (103, 132). In contrast, overexpression of Exo1 in a mre11-nd strain does not lead to an increase in IR resistance (132). The incomplete rescue of mre11 by Exo1 and the failure to rescue mre11-nd mutants support the model that certain DSBs require initial processing by MRX. In addition to a role in initiating DSB resection, the MRX complex may be involved in recruiting the resection nuclease(s) to the site of damage. The high concentration of Exo1 needed to ameliorate the mre11 phenotypes could reflect poor recruitment of Exo1 to DSBs in the absence of MRX. This added recruitment function would explain the relatively mild phenotypes resulting from most mre11-nd alleles compared with the mre11 and other mre11 mutants that are not proficient for MRX complex formation. Finally, the observation that mating-type switching in an exo1mre11-nd double mutant proceeds with wild-type kinetics suggests that at least one additional nuclease is capable of substituting for Mre11 and Exo1 in processing unmodified DSBs (132). Consistent with the idea that Mre11 is a specialized nuclease capable of removing bulky adducts from DNA ends, E. coli SbcC/SbcD efficiently removes biotin-avidin complexes from the 5 ends of DNA duplexes in vitro (37). Additional support for the Mre11-nuclease processing terminal DNA adducts is found in the apparent requirement for the MRN complex in removal of the terminal protein of adenovirus. In early region E4-deleted adenoviruses, newly replicated genomes fail to package owing to extensive MRX-dependent concatemerization mediated through nonhomologous end joining (188, 226). No concatemerization was observed in similar experiments using cells expressing the nuclease-deficient 15 Oct 2004 19:50 246 AR KROGH AR230-GE38-08.tex AR230-GE38-08.sgm LaTeX2e(2002/01/18) P1: GCE SYMINGTON Annu. Rev. Genet. 2004.38:233-271. Downloaded from arjournals.annualreviews.org by UNIVERSITY OF SOUTH CAROL on 02/10/05. For personal use only. Mre11-3 protein (equivalent to the yeast mre11-3 H125L,D126V allele) (188). These results suggest that the Mre11-nuclease is responsible for initiating viral concatemerization by removing the terminal protein-DNA adduct. Sae2, a protein with no known homologues, is a possible fourth member of the MRX complex based on the similarity in meiotic phenotypes of sae2 null and rad50S mutants (123, 158). In addition, the sae2 null mutant is epistatic to and mimics the mitotic phenotypes of nuclease mutant mre11-D56N (160; B.O. Krogh, E.A. Morgan & L.S. Symington, unpublished data). These results indicate that in vivo the catalytic activity of MRX is perhaps modulated by, and only fully functional in, the presence of Sae2. FORMATION OF THE RAD51 NUCLEOPROTEIN FILAMENT Properties of Rad51 The 3 ssDNA tails formed by resection are substrates for binding by the Rad51 protein to promote pairing and strand exchange with the homologous duplex. S. cerevisiae Rad51 is a 43-kDa protein with 30% identity to the catalytic domain of RecA encompassing the Walker A and B motifs for nucleotide binding and/or hydrolysis, referred to as the ATPase domain (AD) (1, 16, 174). The crystal structures of the human RAD51 AD and full-length P. furiosus Rad51 have been determined (151, 172) and show similar structural organization to the AD of RecA protein (187). The PfRad51 N-terminal domain (absent from RecA) forms a small globular domain that is implicated in DNA binding and is separated from the AD by a flexible linker (3, 172). The crystal structure of PfRad51 reveals a double heptameric ring, consistent with ring structures observed by EM (234). The ATPase domains of PfRad51 are arranged as pie-shaped wedges within the heptameric ring. Interaction between monomers is mediated by the polymerization motif (PM) previously defined as the BRC-peptide interaction domain (151, 172). The ring structure is proposed to be the inactive, or storage, form of Rad51, which under appropriate conditions is targeted to ssDNA, disassembles, and then binds to ssDNA as a filament (172). ScRad51 forms right-handed helical filaments on single- and double-stranded DNA with structural similarity to those formed by RecA (142, 200). Binding to ssDNA activates the Rad51 ATPase. Mutation of the conserved lysine residue within the Walker A box to alanine (Rad51-K191A) abolishes DNA binding and ATPase activities of ScRad51. When the same lysine residue is substituted with arginine (Rad51-K191R), the protein retains ATP-dependent DNA binding, but no significant hydrolysis of ATP. In vivo, the rad51-K191A mutation confers a null phenotype and is dominant negative in diploids, whereas rad51-K191R mutants are partially functional for repair and recombination (133, 174, 201). RPA stimulates binding of Rad51 to ssDNA that contains secondary structures, suggesting that the role of RPA in presynapsis is to remove secondary structures from ssDNA to allow formation of a continuous Rad51 nucleoprotein filament (Figure 6). However, RPA 15 Oct 2004 19:50 AR AR230-GE38-08.tex AR230-GE38-08.sgm LaTeX2e(2002/01/18) 247 Annu. Rev. Genet. 2004.38:233-271. Downloaded from arjournals.annualreviews.org by UNIVERSITY OF SOUTH CAROL on 02/10/05. For personal use only. RECOMBINATION PROTEINS IN YEAST P1: GCE Figure 6 Model for strand invasion by Rad51 and mediator proteins, Rad52, Rad54 & Rad55/Rad57 (only one side of the DSB is shown). 1) 3 ssDNA tails are produced by the MRX complex and/or a 5 -3 resection exonuclease. 2) The ssDNA tails are coated by RPA to eliminate secondary structures. 3) Rad52 recruits Rad51 to the RPA-ssDNA complex. 4) The Rad51 nucleoprotein filament extends on the ssDNA mediated by Rad55/Rad57 and RPA is displaced. 5) The Rad51 nucleoprotein filament locates a homologous DNA donor sequence. 6) Rad54 interacts with Rad51, promotes chromatin remodeling, DNA unwinding and strand annealing between donor DNA and the incoming Rad51 nucleoprotein filament. 15 Oct 2004 19:50 248 AR KROGH AR230-GE38-08.tex AR230-GE38-08.sgm LaTeX2e(2002/01/18) P1: GCE SYMINGTON has a higher affinity for ssDNA, and in vitro Rad51 only successfully competes for DNA binding if added to the reaction prior to RPA (199). In vivo, RPA is very abundant and presumably binds to ssDNA prior to Rad51. This poses a problem because Rad51 has to replace RPA to form an active filament for strand exchange. Annu. Rev. Genet. 2004.38:233-271. Downloaded from arjournals.annualreviews.org by UNIVERSITY OF SOUTH CAROL on 02/10/05. For personal use only. Mediators of Rad51 Filament Assembly The severe reduction in Rad51-mediated strand exchange products caused by addition of RPA simultaneously with or prior to Rad51 can be overcome by addition of Rad52 (140, 175, 198). The 52-kDa Rad52 protein interacts directly with both RPA and Rad51 and is thought to mediate Rad51 filament assembly by displacing RPA and simultaneously delivering Rad51 to ssDNA (Figure 6) (182, 194). Mutations that delete the C-terminal region of Rad52, or residues 409–412, responsible for Rad51 interaction are defective in this mediator function in vitro (100, 175). Strains that express C-terminal truncations of Rad52 show higher resistance to IR than rad52 null mutants and are also suppressed by RAD51 present in high copy. Rad51 localization to sites of DSBs in vivo has been shown by immunofluoresence, chromatin immunoprecipitation (ChIP), and by the use of GFP-Rad51 in live cells (19, 49, 128, 193, 230). Localization of Rad51 to the site of DSBs requires Rad52, consistent with the biochemical data indicating Rad52 is essential for loading of Rad51 onto RPA-coated ssDNA (64, 128, 193, 230). The Rad55 and Rad57 proteins are also implicated as mediators in assembly of the Rad51 nucleoprotein filament (Figure 6). Rad55 and Rad57 are referred to as Rad51 paralogs because they share 20%–30% sequence identity to Rad51. Null mutants of rad55 or rad57 are as sensitive to IR as rad51 mutants at low temperatures (20◦ C), but the IR sensitivity is significantly suppressed at 30◦ C (112). This cold sensitivity is thought to be due to the role of Rad55 and Rad57 in stabilization of the Rad51 nucleoprotein filament. The Rad55 and Rad57 proteins interact to form a stable heterodimer that binds to ssDNA and exhibits a weak ATPase activity (68, 87, 199). Mutation of the conserved lysine residue within the Walker A box to alanine in Rad57 confers only a weak DNA repair defect, whereas the equivalent mutation in Rad55 confers a severe defect in response to IR (87). Both RAD55 and RAD57 are required for the formation of Rad51 foci in meiotic cells, but studies in mitotic cells show only a partial defect in Rad51 localization to DSBs (63, 64, 128, 193, 230). Using ChIP, Sugawara et al. found a delay in assembly of the Rad51 filament at an HO-induced DSB, but no pairing between the 3 ssDNA tail and donor sequences, suggesting Rad55 (and presumably Rad57) could have another function after formation of the Rad51 filament, or that the Rad51 filament is only partially assembled and this is insufficient to support homologous pairing (193). Overexpression of RAD51 or point mutations within RAD51 that result in higher affinity of the mutant proteins for DNA suppress the IR sensitivity of rad55 or rad57 mutants, consistent with a role for these proteins in formation or stabilization of the Rad51 filament (55, 68, 87). The purified Rad55/Rad57 heterodimer functions as a mediator in the strand-exchange reaction by allowing 15 Oct 2004 19:50 AR AR230-GE38-08.tex AR230-GE38-08.sgm LaTeX2e(2002/01/18) RECOMBINATION PROTEINS IN YEAST P1: GCE 249 Annu. Rev. Genet. 2004.38:233-271. Downloaded from arjournals.annualreviews.org by UNIVERSITY OF SOUTH CAROL on 02/10/05. For personal use only. Rad51 to nucleate on ssDNA in the presence of RPA (199). Rad55 interacts with Rad51, but not with RPA, suggesting that the mediator activity is distinct from that exerted by Rad52 (68, 87). The Rad54 protein, a member of the Swi2/Snf2 family of chromatin remodeling proteins, also functions to facilitate Rad51 binding to ssDNA in the presence of RPA and to stabilize Rad51 nucleoprotein filaments in vitro (121, 230). However, the significance of these biochemical studies is unclear because Rad51 foci still form in response to DSBs, and persist longer, in rad54 mutants (128). Homologous Pairing and Strand Exchange Once assembled, the Rad51 nucleoprotein filament is capable of interacting with a second DNA molecule, either single- or double-stranded DNA, to initiate strand exchange. Synapsis entails alignment of the nucleoprotein filament with homologous sequences within the second molecule. Although the details of this step of the reaction have not been well characterized for Rad51, studies with RecA protein suggest that the homology search is rapid and involves random collisions of the two molecules. Once homology is found, strand exchange occurs between the two aligned molecules if one of the partners has a free end to allow stable intertwining of the complementary strands. The reaction requires ATP, but is also supported by nonhydrolyzable analogs of ATP. The polarity of strand exchange is 5 to 3 with respect to the complementary strand of the DNA duplex, opposite to the polarity observed for RecA (200). Although the strand transfer assay has been used extensively to characterize recombinases, the reaction between linear ssDNA and homologous supercoiled plasmid DNA (D-loop assay) is thought to better mimic the in vivo substrates. Rad51 and RPA inefficiently promote D-loop formation, but the reaction is greatly stimulated by the addition of Rad54 or the Rad54-related protein Rdh54 (152, 154). Both Rad54 and Rdh54 exhibit dsDNA-specific ATPase activity and translocate on duplex DNA to generate unconstrained negative and positive supercoils (122, 154, 155, 217). These activities are dependent on ATP, and mutation of the conserved lysine within the Walker A box of Rad54 or Rdh54 results in a null phenotype in vivo (35, 154, 155). The ATPase and translocation activities of Rad54 are stimulated by Rad51, or by Rad51-ssDNA complexes (122, 217). Rad54 does not stimulate the pairing activity of RecA, suggesting that the physical interaction between Rad51 and Rad54 is functionally important (35, 86, 122, 217). The strand separation at unwound regions is expected to facilitate the search for homology between duplex DNA and the incoming Rad51 nucleoprotein filament and to promote formation of the nascent joint when homology is located. The ATP-dependent translocation activity of Rad54 on duplex DNA also functions to displace nucleosomes (and presumably non-nucleosomal proteins as well) that could be inhibitory to strand invasion by the Rad51 nucleoprotein filament (6, 84). Sugawara et al. (193) showed homologous pairing between the MAT and HML loci in vivo in rad54 mutants, but DNA synthesis to extend the invading 3 end 15 Oct 2004 19:50 250 AR KROGH AR230-GE38-08.tex AR230-GE38-08.sgm LaTeX2e(2002/01/18) P1: GCE SYMINGTON was not observed, suggesting that the primary function of Rad54 is postsynaptic (Figure 6). Rad54 has been shown to promote heteroduplex DNA extension during strand exchange and to displace Rad51 from duplex DNA (180, 181). Disruption of Rad51-dsDNA complexes could be important for turnover of Rad51 at the end of recombination, to uncover the 3 end of paired intermediates to initiate DNA repair synthesis, or to remove unproductive association of Rad51 with duplex DNA during presynapsis. Annu. Rev. Genet. 2004.38:233-271. Downloaded from arjournals.annualreviews.org by UNIVERSITY OF SOUTH CAROL on 02/10/05. For personal use only. Negative Regulation of Recombination by Srs2 SRS2 encodes a DNA helicase that is suggested to limit heteroduplex extension and/or reverse abnormal recombination intermediates (33, 163, 218). Mutations in SRS2 suppress the UV sensitivity of rad6 and rad18 mutants, and this suppression is thought to occur by channeling lesions from error-prone repair into the recombinational repair pathway (1, 102, 167). Elevated rates of gene conversion are observed between direct repeats in srs2 mutants, but in some DSB-induced recombination assays, the recovery of recombinants is reduced (2, 11). The UV sensitivity of srs2 diploids is suppressed by homozygous mutations in RAD51, RAD52, RAD55, and RAD57, suggesting that lethal recombination intermediates are formed in the absence of srs2 (1). Such intermediates are thought to arise spontaneously in srs2 rad54 and srs2 sgs1 mutants (61, 97). In vitro, the Srs2 protein inhibits strand exchange mediated by Rad51 (101, 221). Srs2 inhibits the reaction by displacement of Rad51 from the presynaptic filament rather than by disruption of joint molecules. The current model is that Srs2 prevents initiation of inappropriate recombination events presynaptically by disrupting the Rad51 filament, but Srs2 could also function at later steps in the removal of Rad51, or by acting on DNA intermediates (81). RAD51-INDEPENDENT RECOMBINATION Rad52: a DNA Annealing Protein The RAD51-dependent pathway for recombination is the most efficient pathway for gene conversion and is required to repair most DSBs in mitotic cells. However, in some assays, significant rates of recombination are detected in rad51 mutants, suggesting an alternate pathway for strand invasion. The alternate pathway requires RAD52 and to some extent, RAD59. Deletion of RAD52 in S. cerevisiae results in severe defects in gene conversion, SSA, recombination between inverted repeats, and formation of survivors in the absence of telomerase (202). In contrast, rad51 mutants show no defect in SSA, a five- to tenfold reduction in the rate of spontaneous recombination between inverted repeats and a defect in one of the two telomere recombination pathways (82, 161, 213). The rad52-327 allele, encoding a truncated protein lacking the Rad51 interaction domain, promotes RAD51-independent recombination, suggesting that a core activity capable 15 Oct 2004 19:50 AR AR230-GE38-08.tex AR230-GE38-08.sgm LaTeX2e(2002/01/18) Annu. Rev. Genet. 2004.38:233-271. Downloaded from arjournals.annualreviews.org by UNIVERSITY OF SOUTH CAROL on 02/10/05. For personal use only. RECOMBINATION PROTEINS IN YEAST P1: GCE 251 of strand invasion is retained in the N-terminal region (212). Intermolecular recombination shows a much greater dependence on RAD51 than intramolecular events, but even in this context HR events are still detected in rad51 mutants and these occur at higher rates than those observed in rad52 mutants (15, 43). These observations suggest that Rad52 can promote efficient intramolecular recombination and a low incidence of intermolecular strand invasion. Both yeast and human Rad52 are multimeric and form ring structures as visualized by electron microscopy (159, 176, 185). Using both conventional and scanning transmission electron microscopy, the HsRad52 protein was observed to form heptameric rings with a strong pinwheel appearance and a central channel (185). Rad52 appears to have two modes of self-association. Assembly of monomers into rings requires sequences in the conserved N-terminal domain of Rad52, whereas the formation of higher-order multimers is mediated by the C terminus (159). Formation of multimers is consistent with genetic studies showing intragenic complementation between N- and C-terminal mutations of rad52 (27). The crystal structure of the N-terminal domain (residues 1–209 or 1–212) of the HsRad52 protein has been determined and reveals an 11-membered ring (90, 177). The overall structure resembles a mushroom, consisting of a stem containing highly conserved hydrophobic residues and a domed cap. A positively charged groove was identified under the domed cap and mutational analysis was consistent with this region comprising a DNA-binding surface (90). One residue identified as important for DNA binding, Arg55, is equivalent to Arg70 of ScRad52, previously shown to be important for Rad52 function in vivo (13, 136). The purified Rad52 protein binds preferentially to single-stranded DNA and promotes annealing of complementary DNA sequences (135). Rad52-promoted annealing of long molecules is stimulated by RPA, whereas Rad52 efficiently anneals oligonucleotides in the absence of RPA (135, 176, 195). The probable role of RPA in strand annealing is to remove secondary structures from single-stranded DNA to allow annealing by Rad52. The important function of Rad52 in SSA is to anneal the complementary single-stranded regions exposed by resection (179). The human Rad52 protein has been shown to promote the formation of D-loops. This activity, as well as DNA binding, is retained in a truncated form of the protein comprising the first 237 residues (89). The formation of D-loops by Rad52 probably occurs by annealing between the incoming single-stranded DNA and transiently single-stranded DNA present in the supercoiled plasmid. Because RAD52 is the only gene known to be essential for RAD51-independent recombination events, it is assumed that the annealing activity of Rad52 can be used for strand invasion (116, 161). Presumably, some donor sequences are transiently single-stranded and can be annealed by Rad52 to ssDNA generated from a resected DSB. Once this annealing step has occurred, the 3 end could be extended by DNA synthesis to stabilize the intermediate. Like Rad51, Rad52 forms discrete foci in response to IR-, HO-, and Spo11induced DSBs and during S-phase of unirradiated cells (64, 109, 128). Surprisingly, only one or two foci are observed using a Rad52-GFP fusion protein, independent 15 Oct 2004 19:50 Annu. Rev. Genet. 2004.38:233-271. Downloaded from arjournals.annualreviews.org by UNIVERSITY OF SOUTH CAROL on 02/10/05. For personal use only. 252 AR KROGH AR230-GE38-08.tex AR230-GE38-08.sgm LaTeX2e(2002/01/18) P1: GCE SYMINGTON of the dose of IR, suggesting that damaged DNA is sequestered at one or two sites within the nucleus, perhaps at specialized repair centers (108). The formation of Rad52 foci is independent of Rad51 (128). The RAD59 gene was identified in a screen for mutants defective for RAD51independent spontaneous mitotic recombination between inverted repeats (14). The rad59 mutation was shown to cause modest defects in several mitotic recombination assays and moderate sensitivity to IR. RAD59 is important for SSA between chromosomal direct repeats, especially as the repeat length decreases (13, 192). This suggests that Rad52 becomes more dependent on Rad59 when the repeats to be annealed are short, or when short homologies are embedded within extensive regions of heterology. Consistent with this hypothesis, recent studies have shown that RAD59 is required for spontaneous recombination between homologous sequences, whereas the RAD51-promoted pathway acts preferentially on sequences with extensive homology (184). RAD52 present in more than one copy suppresses the radiation sensitivity of rad59 mutants, suggesting that the proteins interact and/or have overlapping functions (14). RAD59 encodes a 26-kDa protein with homology to the N terminus of Rad52, which includes the self-interaction and DNA-binding domains of Rad52. Rad52 interacts with Rad59 raising the possibility of formation of a heteromeric Rad52-Rad59 ring (42). Purified Rad59 binds to ss and dsDNA and promotes annealing of complementary ssDNA (42, 153). However, the annealing activity of Rad59 is not stimulated by RPA and even though the biochemical activities of the two proteins are similar, Rad59 is unable to substitute for Rad52 in Rad51independent recombination (14, 42). We suggest Rad59 is a stimulatory factor for Rad52 and is important for recombination between short repeats or when the activity of Rad52 is compromised. Dmc1 DMC1 encodes a 334-amino acid polypeptide with significant similarity to RecA and Rad51, which is expressed only during meiosis (20). Dmc1 is 45% identical to Rad51 along its entire length, and biochemical studies indicate a higher functional conservation than the other Rad51 paralogs to RecA/Rad51. Both Dmc1 and Rad51 form foci during meiosis, and the foci indicate significant colocalization of the two proteins (19). Mutation of DMC1 leads to the accumulation of DSBs in meiosis and a reduction in the frequency of reciprocal recombinants, as measured by physical assay (20). RAD51 and DMC1 are both required for high rates of meiotic recombination and have overlapping, but nonidentical, roles in meiosis (173). Overexpression of RAD51 partially suppresses the recombination and sporulation defects of dmc1 diploids, consistent with the proteins being partially redundant (211). Dmc1 is important to direct repair between homologous nonsister chromatids during meiosis, a function shared by three other meiosis-specific proteins, Hop1, Mek1, and Red1 (169, 223). HsDmc1 has a weak ATPase activity, binds to ss and dsDNA, and catalyzes strand exchange between oligucleotides in vitro (105, 120). The yeast Dmc1 protein 15 Oct 2004 19:50 AR AR230-GE38-08.tex AR230-GE38-08.sgm LaTeX2e(2002/01/18) RECOMBINATION PROTEINS IN YEAST P1: GCE 253 also promotes annealing of complementary ssDNA (74). Dmc1 has been observed to form octameric rings or helical filaments on DNA by EM (120, 148, 169a). Annu. Rev. Genet. 2004.38:233-271. Downloaded from arjournals.annualreviews.org by UNIVERSITY OF SOUTH CAROL on 02/10/05. For personal use only. HOLLIDAY JUNCTION RESOLUTION ACTIVITIES The DSBR model predicts the formation of a dHJ intermediate, which must be resolved for segregation of the recombinant duplexes. Models for the repair of collapsed replication forks and template switching also predict the requirement for branch migration and Holliday junction resolution activities. The Mus81-Mms4 (Eme1) Nuclease The mus81 and eme1 (mms4 in S. cerevisiae) mutants of Schizosaccharomyces pombe exhibit meiotic phenotypes consistent with a defect in the resolution of recombination intermediates. mus81 and eme1 mutants produce less than 1% viable spores and show aberrant segregation of DNA, frequently with the DNA present in only one spore, suggesting a defect in chromosome segregation (21). Remarkably, the spore inviability and unequal DNA distribution observed in mus81 mutants is suppressed by expression of the bacterial HJ resolvase RusA (21). Meiotic crossing over is reduced 20- to 50-fold in mus81 diploids, but gene conversion events unassociated with crossing over occur at normal, or even higher, frequency than in wild-type cells (146, 178). Thus, Mus81 and Eme1 are specifically required to resolve meiotic recombination intermediates to form crossover products in S. pombe. Mus81 and Mms4/Eme1 were identified in two-hybrid screens by interaction with Rad54 and checkpoint kinases, by sensitivity to MMS, and in a screen for mutations that cause synthetic lethality with sgs1 (21, 22, 46, 80, 137, 157). SGS1 encodes a helicase of the RecQ family and is homologous to the human BLM and WRN helicases, which play important roles in genome stability (12, 60). Mus81 has sequence similarity to the nuclease domain of Rad1, and Mms4 has limited similarity to Rad10, suggesting that they are components of a structurespecific nuclease (21, 137). The Mus81-Mms4/Eme1 heterodimer cleaves a variety of branched molecules, including simple Y-structures, duplex Y-structures, and Xforms, with a strong preference for duplex Y-structures (21, 91). The activity on X-form structures is weak, but greatly enhanced if one of the four component strands is discontinuous in the vicinity of the junction (58, 146). Doubts over the generality of Mus81-Mms4 as the HJ resolvase have been raised by genetic studies in S. cerevisiae. In budding yeast, mus81 and mms4 mutants exhibit a sporulation defect, but genetic and physical studies have shown only a twofold decrease in crossing over relative to wild type (45, 46). mus81 mutants exhibit no sensitivity to IR, are proficient for mating-type switching and show no alteration in the proportion of crossover events during DSBR in mitotic cells (80, 81). The conflicting results are best explained by proposing alternate pathways for processing meiotic recombination intermediates in these two yeasts (70, 73, 146). 15 Oct 2004 19:50 254 AR KROGH AR230-GE38-08.tex AR230-GE38-08.sgm LaTeX2e(2002/01/18) P1: GCE SYMINGTON Annu. Rev. Genet. 2004.38:233-271. Downloaded from arjournals.annualreviews.org by UNIVERSITY OF SOUTH CAROL on 02/10/05. For personal use only. The substrate specificity and meiotic crossover defect in S. pombe led Osman et al. (146) to suggest that Mus81-Eme1 cleaves meiotic strand invasion intermediates in two steps, prior to formation of a dHJ, to yield crossover products (Figure 7). They propose that single-end invasion intermediates (SEIs) can be resolved by Mus81Eme1 to form crossover products, or dissociate as envisioned by the SDSA model to form noncrossover products in S. pombe (Figure 7). In budding yeast, a subset of Figure 7 Resolution of recombination intermediates. A dHJ intermediate can be resolved by endonucleolytic cleavage of the junctions or dissolved by the combined actions of BLM (Sgs1) and TOPO IIIα. An unligated strand invasion intermediate can be resolved in two steps to form crossover products by the Mus81-Mms4/Eme1 endonuclease. 15 Oct 2004 19:50 AR AR230-GE38-08.tex AR230-GE38-08.sgm LaTeX2e(2002/01/18) RECOMBINATION PROTEINS IN YEAST P1: GCE 255 SEIs is channeled into a pathway to form dHJ intermediates. These are processed by a bona fide resolvase to form crossover products. This pathway is subject to crossover interference and regulated by Msh4 and Msh5 (45, 73). The interference pathway and synaptonemal complex are absent from S. pombe (146). The other SEIs can be processed in the same way as in S. pombe, to form crossover or noncrossover products by a pathway that is not subject to crossover interference. Annu. Rev. Genet. 2004.38:233-271. Downloaded from arjournals.annualreviews.org by UNIVERSITY OF SOUTH CAROL on 02/10/05. For personal use only. Resolvase A Biochemical evidence in support for two distinct resolvase activities has come from characterization of fractionated mammalian cell extracts. Fractionation of HeLa cell extracts revealed two Holliday junction resolvase activities, one corresponding to Mus81, and the other to the previously described Resolvase A (39, 40). Resolvase A shows high specificity for Holliday junctions and the resulting cleavage products can be ligated (39). In contrast, Mus81 cleaves 3 flap and Y-shaped molecules more efficiently than intact X-forms and the products of resolution are not ligatable (39). Resolvase A promotes a concerted branch migration/resolution reaction similar to that catalyzed by the E. coli RuvABC complex (40). Although the identity of Resolvase A is currently unknown, recent studies indicate that it is intimately associated with the Rad51 paralog, RAD51C (110). Dissolution of dHJs An alternative mode of resolving dHJ intermediates to form noncrossover products is through the action of a topoisomerase. Recent biochemical studies provided direct evidence for resolution of dHJ intermediates by the combined action of BLM and hTOPO IIIα (231). BLM is homologous to S. cerevisiae Sgs1 and was previously shown to exhibit DNA helicase activity to promote branch migration of Holliday junctions (93). As a type IA topoisomerase, Topo III can make singlestranded incisions in ssDNA and catalyzes decatenation of negatively supercoiled plasmids. The yeast TOP3 gene was originally identified in a genetic screen for increased recombination between short repeated sequences (222). The top3 mutant has a severe growth defect, which is suppressed by deletion of SGS1 (60). Like top3 mutants, sgs1 mutants show elevated rates of mitotic recombination and hypersensitivity to chemicals that stall replication forks. Mutations in BLM are causative of the human cancer predisposition syndrome, Bloom’s syndrome (12). A defining cytological feature of cells from BLM patients is the high rate of sister chromatid exchange (32). It is unclear whether the elevated rate of recombination in sgs1 and BLM cell lines is due to increased numbers of recombinogenic lesions formed by aberrant replication, or if the mutants have altered processing of recombination intermediates. sgs1 mutants do not show sensitivity to IR, suggesting that Sgs1 is not essential for DSBR. However, recent studies have shown a significant increase in crossovers during DSBR in sgs1 and top3 mutants (81). This result was interpreted as a role for Sgs1 and Top3 in resolving dHJ recombination intermediates into noncrossover products. The biochemical studies with the equivalent 15 Oct 2004 19:50 Annu. Rev. Genet. 2004.38:233-271. Downloaded from arjournals.annualreviews.org by UNIVERSITY OF SOUTH CAROL on 02/10/05. For personal use only. 256 AR KROGH AR230-GE38-08.tex AR230-GE38-08.sgm LaTeX2e(2002/01/18) P1: GCE SYMINGTON human proteins support this hypothesis (231). Although no resolution activity was detected on single HJs, the dHJ substrate was efficiently resolved by the combined activities of BLM and hTOPO IIIα. Presumably, the branch migration activity of BLM on the constrained substrate creates torsional stress, which activates the decatenation activity of hTOPO IIIα.This mode of resolution, termed dissolution, yields only noncrossover products. In summary, eukaryotic cells resolve recombination intermediates by diverse mechanisms. Resolution of dHJ intermediates can occur by endonucleolytic cleavage to form crossover or noncrossover products, or by dissolution to form noncrossover products. Single-end strand invasion intermediates can be cleaved to form crossover products, or dissociated (SDSA model) to form noncrossovers. The redundancy for this step of the reaction is a plausible explanation for why no single mutant with the predicted resolution-defective phenotype has been identified in budding yeast. NONHOMOLOGOUS END JOINING Nonhomologous end joining is a major pathway for repair of DSBs in higher eukaryotes (85). The key components of this pathway in mammals are the DNAdependent protein kinase (DNA-PK), composed of the Ku heterodimer and DNAPK catalytic subunit (DNA-PKcs), and the DNA ligase IV/XRCC4 complex. Mutations in any of these components result in hypersensitivity to ionizing radiation and in defective V(D)J recombination. Because the rad52 group mutants are hypersensitive to IR, and haploid yeast cells in the G1 stage of the cell cycle show high sensitivity to IR, it was originally thought that yeast cells lack a mammalian-like NHEJ pathway. However, random integration of linear DNA with no homology to chromosomal sequences occurs with low frequency during transformation, and haploid yeast cells have an efficient mechanism for the ligation of plasmids linearized with restriction enzymes when these molecules are used to transform yeast cells (25, 166). The joining of cohesive ends occurs with high fidelity to restore the original sequence of the plasmid (25). However, plasmids with blunt ends, or incompatible overhang ends, are poor substrates for the rejoining pathway. This contrasts with S. pombe and mammalian cells, which show efficient joining of blunt, cohesive ends and incompatible overhangs (117, 164, 228). Although end joining occurs with high efficiency in haploid yeast cells, diploids show much lower levels of NHEJ. As described below, this is because of an essential NHEJ factor that is regulated by mating-type heterozygosity (56, 96, 143, 216). It is assumed that diploids repress NHEJ because a chromosome homologue is available throughout the cell cycle to template HR, whereas haploids cannot use the higher fidelity HR pathway during G1. A heterodimeric protein complex with homology to the mammalian Ku70/Ku80 complex was first identified as a high-affinity DNA end-binding activity in yeast extracts (51, 127). Mutation of the YKU70 or YKU80 genes does not confer sensitivity to IR, MMS, or bleomycin, but the mutants are defective in the rejoining of linear plasmid DNA during transformation, or joining of chromosomal DSBs 15 Oct 2004 19:50 AR AR230-GE38-08.tex AR230-GE38-08.sgm LaTeX2e(2002/01/18) Annu. Rev. Genet. 2004.38:233-271. Downloaded from arjournals.annualreviews.org by UNIVERSITY OF SOUTH CAROL on 02/10/05. For personal use only. RECOMBINATION PROTEINS IN YEAST P1: GCE 257 made with EcoRI or HO (24, 25, 104, 115, 127, 130). The end-joined products recovered from yku70 and yku80 mutants have large deletions and the junctions usually form at small homologies (1–4 nucleotides) (25). The yku70 and yku80 mutants show temperature-sensitive growth, short telomeres, and are defective for transcriptional silencing of genes close to telomeres (telomere position effect, TPE) (26). The Ku70 and Ku80 proteins localize to telomeres in yeast, but in the presence of DNA damage show a Rad9-dependent relocalization to the sites of damage (118). Recent studies have shown a direct interaction between Ku70 and the RNA component of Telomerase, TLC1, and this interaction is important for the telomere protection function of Ku (186). To date, no candidate for the DNA-PKcs has been identified in budding or fission yeast. The DNL4 gene encodes an ATPdependent DNA ligase (Ligase IV), which is required for NHEJ in yeast (165, 204, 229). dnl4 mutants are epistatic to yku70 and yku80 for plasmid rejoining but Figure 8 Repair of DSBs by NHEJ. DSB ends are stabilized and tethered together by the Ku70/Ku80 heterodimer and the MRX complex. The Ligase IV complex, Dnl4Lif1, is recruited to ligate the ends by interactions to Xrs2. The haploid specific protein, Nej1, is responsible for nuclear localization of Lif1. 15 Oct 2004 19:50 Annu. Rev. Genet. 2004.38:233-271. Downloaded from arjournals.annualreviews.org by UNIVERSITY OF SOUTH CAROL on 02/10/05. For personal use only. 258 AR KROGH AR230-GE38-08.tex AR230-GE38-08.sgm LaTeX2e(2002/01/18) P1: GCE SYMINGTON have normal growth, telomere length, and TPE, suggesting that these functions of Ku70/Ku80 are independent of their role in NHEJ. As in mammalian cells, an accessory protein, Lif1, activates and targets Dnl4 to DSBs in yeast (69, 205). The phenotype of lif1 mutants is indistinguishable from that of dnl4 mutants. Another factor that interacts with Lif1, Nej1/Lif2, was identified by two-hybrid analysis, in a genetic screen for NHEJ mutants, and as a haploid-specific gene (56, 96, 143, 216). Diploid MATa/MATα cells that constitutively express NEJ1 are proficient for NHEJ, indicating that this is the only factor in the pathway regulated by mating type. Nej1 is required for nuclear localization of Lif1 (216), but it is currently unknown if this is the only role for Nej1 in NHEJ (Figure 8). In S. cerevisiae, Rad50, Mre11, and Xrs2 are also required for NHEJ, but these factors play no role in NHEJ in S. pombe (117, 130). rad50, mre11, and xrs2 mutants are as defective in plasmid end joining as yku70 and yku80 mutants, but the rare junctions formed usually show no loss of nucleotides (26). The Mre11nuclease activity is only required for end joining if the ends have hairpins, and this specialized end-joining reaction also requires Sae2 and Pso2, the putative homolog of the human V(D)J-recombination nuclease, Artemis (235). A chromosomal assay for joining of incompatible ends revealed a requirement for Mre11 and Dnl4, but not Yku70, suggesting there are at least two pathways for NHEJ (114). Genetic and biochemical studies in animal cells also support the existence of another endjoining pathway, independent of that promoted by DNA-PK (67, 224). The purified MRX complex stimulates intermolecular DNA joining by the Dnl4/Lif1 complex (34). AFM analysis revealed juxtaposition of DNA ends by the MRX complex to form linear concatamers, suggesting that the MRX complex can align DNA molecules for ligation (Figure 4). Xrs2 interacts with Lif1, suggesting that Xrs2 might function to recruit Dnl4/Lif1 to ends held together by the MRX complex. Purified DNA-PK has been shown to synapse DNA ends, raising the possibility that the MRX complex has assumed this role in S. cerevisiae (47). ACKNOWLEDGMENTS We are grateful to the members of our laboratory, past and present, for stimulating discussions. Research performed in our laboratory that is cited in this review was supported by grants from the National Institutes of Health (GM41784 and GM54099). B.O.K. was supported by fellowships from the Danish Natural Science Research Council and The Carlsberg Foundation. The Annual Review of Genetics is online at http://genet.annualreviews.org LITERATURE CITED 1. Aboussekhra A, Chanet R, Adjiri A, Fabre F. 1992. Semidominant suppressors of Srs2 helicase mutations of Saccharomyces cerevisiae map in the RAD51 gene, whose sequence predicts a protein with similarities to procaryotic RecA proteins. Mol. Cell Biol. 12:3224– 34 15 Oct 2004 19:50 AR AR230-GE38-08.tex AR230-GE38-08.sgm LaTeX2e(2002/01/18) Annu. Rev. Genet. 2004.38:233-271. Downloaded from arjournals.annualreviews.org by UNIVERSITY OF SOUTH CAROL on 02/10/05. For personal use only. RECOMBINATION PROTEINS IN YEAST 2. Aguilera A, Klein HL. 1988. Genetic control of intrachromosomal recombination in Saccharomyces cerevisiae. I. Isolation and genetic characterization of hyper-recombination mutations. Genetics 119:779–90 3. Aihara H, Ito Y, Kurumizaka H, Yokoyama S, Shibata T. 1999. The Nterminal domain of the human Rad51 protein binds DNA: structure and a DNA binding surface as revealed by NMR. J. Mol. Biol. 290:495–504 4. Alani E, Padmore R, Kleckner N. 1990. Analysis of wild-type and rad50 mutants of yeast suggests an intimate relationship between meiotic chromosome synapsis and recombination. Cell 61:419–36 5. Alani E, Subbiah S, Kleckner N. 1989. The yeast RAD50 gene encodes a predicted 153-kD protein containing a purine nucleotide-binding domain and two large heptad-repeat regions. Genetics 122:47–57 6. Alexeev A, Mazin A, Kowalczykowski SC. 2003. Rad54 protein possesses chromatin-remodeling activity stimulated by the Rad51-ssDNA nucleoprotein filament. Nat. Struct. Biol. 10:182– 86 7. Allers T, Lichten M. 2001. Differential timing and control of noncrossover and crossover recombination during meiosis. Cell 106:47–57 8. Anderson DE, Trujillo KM, Sung P, Erickson HP. 2001. Structure of the Rad50· Mre11 DNA repair complex from Saccharomyces cerevisiae by electron microscopy. J. Biol. Chem. 276:37027–33 9. Aravind L, Walker DR, Koonin EV. 1999. Conserved domains in DNA repair proteins and evolution of repair systems. Nucleic Acids Res. 27:1223–42 10. Arora C, Kee K, Maleki S, Keeney S. 2004. Antiviral protein Ski8 is a direct partner of Spo11 in meiotic DNA break formation, independent of its cytoplasmic role in RNA metabolism. Mol. Cell 13:549–59 P1: GCE 259 11. Aylon Y, Liefshitz B, Bitan-Banin G, Kupiec M. 2003. Molecular dissection of mitotic recombination in the yeast Saccharomyces cerevisiae. Mol. Cell Biol. 23:1403–17 12. Bachrati CZ, Hickson ID. 2003. RecQ helicases: suppressors of tumorigenesis and premature aging. Biochem. J. 374: 577–606 13. Bai Y, Davis AP, Symington LS. 1999. A novel allele of RAD52 that causes severe DNA repair and recombination deficiencies only in the absence of RAD51 or RAD59. Genetics 153:1117–30 14. Bai Y, Symington LS. 1996. A Rad52 homolog is required for RAD51independent mitotic recombination in Saccharomyces cerevisiae. Genes Dev. 10:2025–37 15. Bartsch S, Kang LE, Symington LS. 2000. RAD51 is required for the repair of plasmid double-stranded DNA gaps from either plasmid or chromosomal templates. Mol. Cell Biol. 20:1194– 205 16. Basile G, Aker M, Mortimer RK. 1992. Nucleotide sequence and transcriptional regulation of the yeast recombinational repair gene RAD51. Mol. Cell Biol. 12: 3235–46 17. Baudat F, Nicolas A. 1997. Clustering of meiotic double-strand breaks on yeast chromosome III. Proc. Natl. Acad. Sci. USA 94:5213–18 18. Bergerat A, de Massy B, Gadelle D, Varoutas PC, Nicolas A, Forterre P. 1997. An atypical topoisomerase II from Archaea with implications for meiotic recombination. Nature 386:414–17 19. Bishop DK. 1994. RecA homologs Dmc1 and Rad51 interact to form multiple nuclear complexes prior to meiotic chromosome synapsis. Cell 79:1081– 92 20. Bishop DK, Park D, Xu L, Kleckner N. 1992. DMC1: a meiosis-specific yeast homolog of E. coli recA required for recombination, synaptonemal complex 15 Oct 2004 19:50 260 21. Annu. Rev. Genet. 2004.38:233-271. Downloaded from arjournals.annualreviews.org by UNIVERSITY OF SOUTH CAROL on 02/10/05. For personal use only. 22. 23. 24. 25. 26. 27. 28. 29. AR KROGH AR230-GE38-08.tex AR230-GE38-08.sgm LaTeX2e(2002/01/18) P1: GCE SYMINGTON formation, and cell cycle progression. Cell 69:439–56 Boddy MN, Gaillard PH, McDonald WH, Shanahan P, Yates JR, 3rd, Russell P. 2001. Mus81-Eme1 are essential components of a Holliday junction resolvase. Cell 107:537–48 Boddy MN, Lopez-Girona A, Shanahan P, Interthal H, Heyer WD, Russell P. 2000. Damage tolerance protein Mus81 associates with the FHA1 domain of checkpoint kinase Cds1. Mol. Cell Biol. 20:8758–66 Borde V, Lin W, Novikov E, Petrini JH, Lichten M, Nicolas A. 2004. Association of Mre11p with double-strand break sites during yeast meiosis. Mol. Cell. 13:389– 401 Boulton SJ, Jackson SP. 1996. Identification of a Saccharomyces cerevisiae Ku80 homologue: roles in DNA double-strand break rejoining and in telomeric maintenance. Nucleic Acids Res. 24:4639–48 Boulton SJ, Jackson SP. 1996. Saccharomyces cerevisiae Ku70 potentiates illegitimate DNA double-strand break repair and serves as a barrier to errorprone DNA repair pathways. EMBO J. 15:5093–103 Boulton SJ, Jackson SP. 1998. Components of the Ku-dependent nonhomologous end-joining pathway are involved in telomeric length maintenance and telomeric silencing. EMBO J. 17: 1819–28 Boundy-Mills KL, Livingston DM. 1993. A Saccharomyces cerevisiae RAD52 allele expressing a C-terminal truncation protein: activities and intragenic complementation of missense mutations. Genetics 133:39–49 Bressan DA, Baxter BK, Petrini JH. 1999. The Mre11-Rad50-Xrs2 protein complex facilitates homologous recombination-based double-strand break repair in Saccharomyces cerevisiae. Mol. Cell Biol. 19:7681–87 Bressan DA, Olivares HA, Nelms BE, 30. 31. 32. 33. 34. 35. 36. 37. 38. Petrini JH. 1998. Alteration of Nterminal phosphoesterase signature motifs inactivates Saccharomyces cerevisiae Mre11. Genetics 150:591–600 Cao L, Alani E, Kleckner N. 1990. A pathway for generation and processing of double-strand breaks during meiotic recombination in S. cerevisiae. Cell 61:1089–101 Carney JP, Maser RS, Olivares H, Davis EM, Le Beau M, et al. 1998. The hMre11/hRad50 protein complex and Nijmegen breakage syndrome: linkage of double-strand break repair to the cellular DNA damage response. Cell 93:477– 86 Chaganti RS, Schonberg S, German J. 1974. A manyfold increase in sister chromatid exchanges in Bloom’s syndrome lymphocytes. Proc. Natl. Acad. Sci. USA 71:4508–12 Chanet R, Heude M, Adjiri A, Maloisel L, Fabre F. 1996. Semidominant mutations in the yeast Rad51 protein and their relationships with the Srs2 helicase. Mol. Cell Biol. 16:4782–89 Chen L, Trujillo K, Ramos W, Sung P, Tomkinson AE. 2001. Promotion of Dnl4-catalyzed DNA end-joining by the Rad50/Mre11/Xrs2 and Hdf1/Hdf2 complexes. Mol. Cell 8:1105–15 Clever B, Interthal H, Schmuckli-Maurer J, King J, Sigrist M, Heyer WD. 1997. Recombinational repair in yeast: functional interactions between Rad51 and Rad54 proteins. EMBO J. 16:2535–44 Colleaux L, D’Auriol L, Galibert F, Dujon B. 1988. Recognition and cleavage site of the intron-encoded omega transposase. Proc. Natl. Acad. Sci. USA 85: 6022–26 Connelly JC, de Leau ES, Leach DR. 2003. Nucleolytic processing of a protein-bound DNA end by the E. coli SbcCD (MR) complex. DNA Repair 2:795–807 Connelly JC, de Leau ES, Okely EA, Leach DR. 1997. Overexpression, 15 Oct 2004 19:50 AR AR230-GE38-08.tex AR230-GE38-08.sgm LaTeX2e(2002/01/18) RECOMBINATION PROTEINS IN YEAST 39. Annu. Rev. Genet. 2004.38:233-271. Downloaded from arjournals.annualreviews.org by UNIVERSITY OF SOUTH CAROL on 02/10/05. For personal use only. 40. 41. 42. 43. 44. 45. 46. 47. 48. purification, and characterization of the SbcCD protein from Escherichia coli. J. Biol. Chem. 272:19819–26 Constantinou A, Chen X-B, McGowan CH, West SC. 2002. Human Holliday junction resolvases: two activities with distinct substrate specificities. EMBO J. 21:5577–85 Constantinou A, Davies AA, West SC. 2001. Branch migration and Holliday junction resolution catalyzed by activities from mammalian cells. Cell 104: 259–68 D’Amours D, Jackson SP. 2001. The yeast Xrs2 complex functions in S phase checkpoint regulation. Genes Dev. 15:2238–49 Davis AP, Symington LS. 2001. The yeast recombinational repair protein Rad59 interacts with Rad52 and stimulates single-strand annealing. Genetics 159:515–25 Davis AP, Symington LS. 2004. RAD51dependent break-induced replication in yeast. Mol. Cell Biol. 24:2344–51 de Jager M, van Noort J, van Gent DC, Dekker C, Kanaar R, Wyman C. 2001. Human Rad50/Mre11 is a flexible complex that can tether DNA ends. Mol. Cell 8:1129–35 de los Santos T, Hunter N, Lee C, Larkin B, Loidl J, Hollingsworth NM. 2003. The Mus81/Mms4 endonuclease acts independently of double-Holliday junction resolution to promote a distinct subset of crossovers during meiosis in budding yeast. Genetics 164:81–94 de los Santos T, Loidl J, Larkin B, Hollingsworth NM. 2001. A role for MMS4 in the processing of recombination intermediates during meiosis in Saccharomyces cerevisiae. Genetics 159: 1511–25 DeFazio LG, Stansel RM, Griffith JD, Chu G. 2002. Synapsis of DNA ends by DNA-dependent protein kinase. EMBO J. 21:3192–200 Diaz RL, Alcid AD, Berger JM, Keeney 49. 50. 51. 52. 53. 54. 55. 56. P1: GCE 261 S. 2002. Identification of residues in yeast Spo11p critical for meiotic DNA double-strand break formation. Mol. Cell Biol. 22:1106–15 Essers J, Houtsmuller AB, van Veelen L, Paulusma C, Nigg AL, et al. 2002. Nuclear dynamics of RAD52 group homologous recombination proteins in response to DNA damage. EMBO J. 21:2030–37 Fabre F, Chan A, Heyer WD, Gangloff S. 2002. Alternate pathways involving Sgs1/Top3, Mus81/ Mms4, and Srs2 prevent formation of toxic recombination intermediates from single-stranded gaps created by DNA replication. Proc. Natl. Acad. Sci. USA 99:16887–92 Feldmann H, Winnacker EL. 1993. A putative homologue of the human autoantigen Ku from Saccharomyces cerevisiae. J. Biol. Chem. 268:12895–900 Ferguson DO, Holloman WK. 1996. Recombinational repair of gaps in DNA is asymmetric in Ustilago maydis and can be explained by a migrating Dloop model. Proc. Natl. Acad. Sci. USA 93:5419–24 Fiorentini P, Huang KN, Tishkoff DX, Kolodner RD, Symington LS. 1997. Exonuclease I of Saccharomyces cerevisiae functions in mitotic recombination in vivo and in vitro. Mol. Cell Biol. 17:2764–73 Fishman-Lobell J, Rudin N, Haber JE. 1992. Two alternative pathways of double-strand break repair that are kinetically separable and independently modulated. Mol. Cell Biol. 12:1292– 303 Fortin GS, Symington LS. 2002. Mutations in yeast Rad51 that partially bypass the requirement for Rad55 and Rad57 in DNA repair by increasing stability of Rad51/DNA complexes. EMBO J. 21:3160–70 Frank-Vaillant M, Marcand S. 2001. NHEJ regulation by mating type is exercised through a novel protein, Lif2p, 15 Oct 2004 19:50 262 57. Annu. Rev. Genet. 2004.38:233-271. Downloaded from arjournals.annualreviews.org by UNIVERSITY OF SOUTH CAROL on 02/10/05. For personal use only. 58. 59. 60. 61. 62. 63. 64. 65. AR KROGH AR230-GE38-08.tex AR230-GE38-08.sgm LaTeX2e(2002/01/18) P1: GCE SYMINGTON essential to the ligase IV pathway. Genes Dev. 15:3005–12 Furuse M, Nagase Y, Tsubouchi H, Murakami-Murofushi K, Shibata T, Ohta K. 1998. Distinct roles of two separable in vitro activities of yeast Mre11 in mitotic and meiotic recombination. EMBO J. 17:6412–25 Gaillard PH, Noguchi E, Shanahan P, Russell P. 2003. The endogenous Mus81Eme1 complex resolves Holliday junctions by a nick and counternick mechanism. Mol. Cell 12:747–59 Game JC, Mortimer RK. 1974. A genetic study of X-ray sensitive mutants in yeast. Mutat. Res. 24:281–92 Gangloff S, McDonald JP, Bendixen C, Arthur L, Rothstein R. 1994. The yeast type I topoisomerase Top3 interacts with Sgs1, a DNA helicase homolog: a potential eukaryotic reverse gyrase. Mol. Cell Biol. 14:8391–98 Gangloff S, Soustelle C, Fabre F. 2000. Homologous recombination is responsible for cell death in the absence of the Sgs1 and Srs2 helicases. Nat. Genet. 25:192–94 Garvik B, Carson M, Hartwell L. 1995. Single-stranded DNA arising at telomeres in cdc13 mutants may constitute a specific signal for the RAD9 checkpoint. Mol. Cell Biol. 15:6128–38 Gasior SL, Olivares H, Ear U, Hari DM, Weichselbaum R, Bishop DK. 2001. Assembly of RecA-like recombinases: distinct roles for mediator proteins in mitosis and meiosis. Proc. Natl. Acad. Sci. USA 98:8411–18 Gasior SL, Wong AK, Kora Y, Shinohara A, Bishop DK. 1998. Rad52 associates with RPA and functions with rad55 and rad57 to assemble meiotic recombination complexes. Genes Dev. 12:2208– 21 Gerton JL, DeRisi J, Shroff R, Lichten M, Brown PO, Petes TD. 2000. Inaugural article: global mapping of meiotic recombination hotspots and coldspots 66. 67. 68. 69. 70. 71. 72. 73. 74. 75. in the yeast Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 97:11383–90 Hackett JA, Greider CW. 2003. End resection initiates genomic instability in the absence of telomerase. Mol. Cell Biol. 23:8450–61 Han JO, Erskine LA, Purugganan MM, Stamato TD, Roth DB. 1998. V(D)J recombination intermediates and nonstandard products in XRCC4-deficient cells. Nucleic Acids Res. 26:3769–75 Hays SL, Firmenich AA, Berg P. 1995. Complex formation in yeast doublestrand break repair: participation of Rad51, Rad52, Rad55, and Rad57 proteins. Proc. Natl. Acad. Sci. USA 92: 6925–29 Herrmann G, Lindahl T, Schar P. 1998. Saccharomyces cerevisiae LIF1: a function involved in DNA double-strand break repair related to mammalian XRCC4. EMBO J. 17:4188–98 Heyer WD, Ehmsen KT, Solinger JA. 2003. Holliday junctions in the eukaryotic nucleus: resolution in sight? Trends Biochem. Sci. 28:548–57 Higgins NP, Kato K, Strauss B. 1976. A model for replication repair in mammalian cells. J. Mol. Biol. 101:417–25 Holliday R. 1964. A mechanism for gene conversion in fungi. Genet. Res. 5:282– 304 Hollingsworth NM, Brill SJ. 2004. The Mus81 solution to resolution: generating meiotic crossovers without Holliday junctions. Genes Dev. 18:117–25 Hong EL, Shinohara A, Bishop DK. 2001. Saccharomyces cerevisiae Dmc1 protein promotes renaturation of singlestrand DNA (ssDNA) and assimilation of ssDNA into homologous super-coiled duplex DNA. J. Biol. Chem. 276:41906– 12 Hopfner KP, Craig L, Moncalian G, Zinkel RA, Usui T, et al. 2002. The Rad50 zinc-hook is a structure joining Mre11 complexes in DNA recombination and repair. Nature 418:562–66 15 Oct 2004 19:50 AR AR230-GE38-08.tex AR230-GE38-08.sgm LaTeX2e(2002/01/18) Annu. Rev. Genet. 2004.38:233-271. Downloaded from arjournals.annualreviews.org by UNIVERSITY OF SOUTH CAROL on 02/10/05. For personal use only. RECOMBINATION PROTEINS IN YEAST 76. Hopfner KP, Karcher A, Craig L, Woo TT, Carney JP, Tainer JA. 2001. Structural biochemistry and interaction architecture of the DNA double-strand break repair Mre11 nuclease and Rad50ATPase. Cell 105:473–85 77. Hopfner KP, Karcher A, Shin D, Fairley C, Tainer JA, Carney JP. 2000. Mre11 and Rad50 from Pyrococcus furiosus: cloning and biochemical characterization reveal an evolutionarily conserved multiprotein machine. J. Bacteriol. 182:6036–41 78. Hopfner KP, Karcher A, Shin DS, Craig L, Arthur LM, et al. 2000. Structural biology of Rad50 ATPase: ATP-driven conformational control in DNA doublestrand break repair and the ABC-ATPase superfamily. Cell 101:789–800 79. Hurst DD, Fogel S, Mortimer RK. 1972. Conversion-associated recombination in yeast (hybrids-meiosis-tetradsmarker loci-models). Proc. Natl. Acad. Sci. USA 69:101–5 80. Interthal H, Heyer WD. 2000. MUS81 encodes a novel helix-hairpin-helix protein involved in the response to UVand methylation-induced DNA damage in Saccharomyces cerevisiae. Mol. Gen. Genet. 263:812–27 81. Ira G, Malkova A, Liberi G, Foiani M, Haber JE. 2003. Srs2 and Sgs1Top3 suppress crossovers during doublestrand break repair in yeast. Cell 115: 401–11 82. Ivanov EL, Sugawara N, FishmanLobell J, Haber JE. 1996. Genetic requirements for the single-strand annealing pathway of double-strand break repair in Saccharomyces cerevisiae. Genetics 142:693–704 83. Ivanov EL, Sugawara N, White CI, Fabre F, Haber JE. 1994. Mutations in XRS2 and RAD50 delay but do not prevent mating-type switching in Saccharomyces cerevisiae. Mol. Cell Biol. 14:3414–25 84. Jaskelioff M, Van Komen S, Krebs JE, 85. 86. 87. 88. 89. 90. 91. 92. 93. P1: GCE 263 Sung P, Peterson CL. 2003. Rad54p is a chromatin remodeling enzyme required for heteroduplex DNA joint formation with chromatin. J. Biol. Chem. 278:9212–18 Jeggo PA. 1998. Identification of genes involved in repair of DNA double-strand breaks in mammalian cells. Radiat. Res. 150:S80–91 Jiang H, Xie Y, Houston P, Stemke-Hale K, Mortensen UH, et al. 1996. Direct association between the yeast Rad51 and Rad54 recombination proteins. J. Biol. Chem. 271:33181–86 Johnson RD, Symington LS. 1995. Functional differences and interactions among the putative RecA homologs Rad51, Rad55, and Rad57. Mol. Cell Biol. 15:4843–50 Johzuka K, Ogawa H. 1995. Interaction of Mre11 and Rad50: two proteins required for DNA repair and meiosisspecific double-strand break formation in Saccharomyces cerevisiae. Genetics 139:1521–32 Kagawa W, Kurumizaka H, Ikawa S, Yokoyama S, Shibata T. 2001. Homologous pairing promoted by the human Rad52 protein. J. Biol. Chem. 276: 35201–8 Kagawa W, Kurumizaka H, Ishitani R, Fukai S, Nureki O, et al. 2002. Crystal structure of the homologous-pairing domain from the human Rad52 recombinase in the undecameric form. Mol. Cell. 10:359–71 Kaliraman V, Mullen JR, Fricke WM, Bastin-Shanower SA, Brill SJ. 2001. Functional overlap between Sgs1-Top3 and the Mms4-Mus81 endonuclease. Genes Dev. 15:2730–40 Kaplun L, Ivantsiv Y, Kornitzer D, Raveh D. 2000. Functions of the DNA damage response pathway target Ho endonuclease of yeast for degradation via the ubiquitin-26S proteasome system. Proc. Natl. Acad. Sci. USA 97:10077–82 Karow JK, Constantinou A, Li JL, West 15 Oct 2004 19:50 264 94. Annu. Rev. Genet. 2004.38:233-271. Downloaded from arjournals.annualreviews.org by UNIVERSITY OF SOUTH CAROL on 02/10/05. For personal use only. 95. 96. 97. 98. 99. 100. 101. 102. 103. AR KROGH AR230-GE38-08.tex AR230-GE38-08.sgm LaTeX2e(2002/01/18) P1: GCE SYMINGTON SC, Hickson ID. 2000. The Bloom’s syndrome gene product promotes branch migration of Holliday junctions. Proc. Natl. Acad. Sci. USA 97:6504–8 Keeney S. 2001. Mechanism and control of meiotic recombination initiation. Curr. Top. Dev. Biol. 52:1–53 Keeney S, Giroux CN, Kleckner N. 1997. Meiosis-specific DNA doublestrand breaks are catalyzed by Spo11, a member of a widely conserved protein family. Cell 88:375–84 Kegel A, Sjostrand JO, Astrom SU. 2001. Nej1p, a cell type-specific regulator of nonhomologous end joining in yeast. Curr. Biol. 11:1611–17 Klein HL. 2001. Mutations in recombinational repair and in checkpoint control genes suppress the lethal combination of srs2 with other DNA repair genes in Saccharomyces cerevisiae. Genetics 157:557–65 Kostriken R, Heffron F. 1984. The product of the HO gene is a nuclease: purification and characterization of the enzyme. Cold Spring Harbor Symp. Quant. Biol. 49:89–96 Kraus E, Leung WY, Haber JE. 2001. Break-induced replication: a review and an example in budding yeast. Proc. Natl. Acad. Sci. USA 98:8255–62 Krejci L, Song B, Bussen W, Rothstein R, Mortensen UH, Sung P. 2002. Interaction with Rad51 is indispensable for recombination mediator function of Rad52. J. Biol. Chem. 277:40132–41 Krejci L, Van Komen S, Li Y, Villemain J, Reddy MS, et al. 2003. DNA helicase Srs2 disrupts the Rad51 presynaptic filament. Nature 423:305–9 Lawrence CW, Christensen RB. 1979. Metabolic suppressors of trimethoprim and ultraviolet light sensitivities of Saccharomyces cerevisiae rad6 mutants. J. Bacteriol. 139:866–87 Lee S-E, Bressan DA, Petrini JH, Haber JE. 2002. Complementation between N-terminal Saccharomyces cerevisiae 104. 105. 106. 107. 108. 109. 110. 111. 112. 113. mre11 alleles in DNA repair and telomere length maintenance. DNA Repair 1:27–40 Lewis LK, Kirchner JM, Resnick MA. 1998. Requirement for end-joining and checkpoint functions, but not RAD52-mediated recombination, after EcoRI endonuclease cleavage of Saccharomyces cerevisiae DNA. Mol. Cell Biol. 18:1891–902 Li Z, Golub EI, Gupta R, Radding CM. 1997. Recombination activities of HsDmc1 protein, the meiotic human homolog of RecA protein. Proc. Natl. Acad. Sci. USA 94:11221–26 Lim DS, Kim ST, Xu B, Maser RS, Lin J, et al. 2000. ATM phosphorylates p95/nbs1 in an S-phase checkpoint pathway. Nature 404:613–17 Lin FL, Sperle K, Sternberg N. 1984. Model for homologous recombination during transfer of DNA into mouse L cells: role for DNA ends in the recombination process. Mol. Cell Biol. 4:1020– 34 Lisby M, Mortensen UH, Rothstein R. 2003. Colocalization of multiple DNA double-strand breaks at a single Rad52 repair centre. Nat. Cell Biol. 5:572–77 Lisby M, Rothstein R, Mortensen UH. 2001. Rad52 forms DNA repair and recombination centers during S phase. Proc. Natl. Acad. Sci. USA 98:8276–82 Liu Y, Masson JY, Shah R, O’Regan P, West SC. 2004. RAD51C is required for Holliday junction processing in mammalian cells. Science 303:243–46 Lopes M, Cotta-Ramusino C, Pellicioli A, Liberi G, Plevani P, et al. 2001. The DNA replication checkpoint response stabilizes stalled replication forks. Nature 412:557–61 Lovett ST, Mortimer RK. 1987. Characterization of null mutants of the RAD55 gene of Saccharomyces cerevisiae: effects of temperature, osmotic strength and mating type. Genetics 116:547–53 Lundblad V, Blackburn EH. 1993. An 15 Oct 2004 19:50 AR AR230-GE38-08.tex AR230-GE38-08.sgm LaTeX2e(2002/01/18) RECOMBINATION PROTEINS IN YEAST 114. Annu. Rev. Genet. 2004.38:233-271. Downloaded from arjournals.annualreviews.org by UNIVERSITY OF SOUTH CAROL on 02/10/05. For personal use only. 115. 116. 117. 118. 119. 120. 121. 122. alternative pathway for yeast telomere maintenance rescues est1-senescence. Cell 73:347–60 Ma JL, Kim EM, Haber JE, Lee SE. 2003. Yeast Mre11 and Rad1 proteins define a Ku-independent mechanism to repair double-strand breaks lacking overlapping end sequences. Mol. Cell Biol. 23:8820–28 Mages GJ, Feldmann HM, Winnacker EL. 1996. Involvement of the Saccharomyces cerevisiae HDF1 gene in DNA double-strand break repair and recombination. J. Biol. Chem. 271:7910–15 Malkova A, Ivanov EL, Haber JE. 1996. Double-strand break repair in the absence of RAD51 in yeast: a possible role for break-induced DNA replication. Proc. Natl. Acad. Sci. USA 93:7131–36 Manolis KG, Nimmo ER, Hartsuiker E, Carr AM, Jeggo PA, Allshire RC. 2001. Novel functional requirements for non-homologous DNA end joining in Schizosaccharomyces pombe. EMBO J. 20:210–21 Martin SG, Laroche T, Suka N, Grunstein M, Gasser SM. 1999. Relocalization of telomeric Ku and SIR proteins in response to DNA strand breaks in yeast. Cell 97:621–33 Maryon E, Carroll D. 1991. Characterization of recombination intermediates from DNA injected into Xenopus laevis oocytes: evidence for a nonconservative mechanism of homologous recombination. Mol. Cell Biol. 11:3278–87 Masson JY, Davies AA, Hajibagheri N, Van Dyck E, Benson FE, et al. 1999. The meiosis-specific recombinase hDmc1 forms ring structures and interacts with hRad51. EMBO J. 18:6552–60 Mazin AV, Alexeev AA, Kowalczykowski SC. 2003. A novel function of Rad54 protein. Stabilization of the Rad51 nucleoprotein filament. J. Biol. Chem. 278:14029–36 Mazin AV, Bornarth CJ, Solinger JA, Heyer WD, Kowalczykowski SC. 2000. 123. 124. 125. 126. 127. 128. 129. 130. 131. P1: GCE 265 Rad54 protein is targeted to pairing loci by the Rad51 nucleoprotein filament. Mol. Cell 6:583–92 McKee AH, Kleckner N. 1997. A general method for identifying recessive diploid-specific mutations in Saccharomyces cerevisiae, its application to the isolation of mutants blocked at intermediate stages of meiotic prophase and characterization of a new gene SAE2. Genetics 146:797–816 Merker JD, Dominska M, Petes TD. 2003. Patterns of heteroduplex formation associated with the initiation of meiotic recombination in the yeast Saccharomyces cerevisiae. Genetics 165:47–63 Meselson MS, Radding CM. 1975. A general model for genetic recombination. Proc. Natl. Acad. Sci. USA 72:358– 61 Michel B, Ehrlich SD, Uzest M. 1997. DNA double-strand breaks caused by replication arrest. EMBO J. 16:430– 38 Milne GT, Jin S, Shannon KB, Weaver DT. 1996. Mutations in two Ku homologs define a DNA end-joining repair pathway in Saccharomyces cerevisiae. Mol. Cell Biol. 16:4189–98 Miyazaki T, Bressan DA, Shinohara M, Haber JE, Shinohara A. 2004. In vivo assembly and disassembly of Rad51 and Rad52 complexes during double-strand break repair. EMBO J. 23:939–49 Moncalian G, Lengsfeld B, Bhaskara V, Hopfner KP, Karcher A, et al. 2004. The rad50 signature motif: essential to ATP binding and biological function. J. Mol. Biol. 335:937–51 Moore JK, Haber JE. 1996. Cell cycle and genetic requirements of two pathways of nonhomologous end-joining repair of double-strand breaks in Saccharomyces cerevisiae. Mol. Cell Biol. 16: 2164–73 Moreau S, Ferguson JR, Symington LS. 1999. The nuclease activity of Mre11 is required for meiosis but not for mating 15 Oct 2004 19:50 266 132. Annu. Rev. Genet. 2004.38:233-271. Downloaded from arjournals.annualreviews.org by UNIVERSITY OF SOUTH CAROL on 02/10/05. For personal use only. 133. 134. 135. 136. 137. 138. 139. 140. AR KROGH AR230-GE38-08.tex AR230-GE38-08.sgm LaTeX2e(2002/01/18) P1: GCE SYMINGTON type switching, end joining, or telomere maintenance. Mol. Cell Biol. 19:556– 66 Moreau S, Morgan EA, Symington LS. 2001. Overlapping functions of the Saccharomyces cerevisiae Mre11, Exo1 and Rad27 nucleases in DNA metabolism. Genetics 159:1423–33 Morgan EA, Shah N, Symington LS. 2002. The requirement for ATP hydrolysis by Saccharomyces cerevisiae Rad51 is bypassed by mating-type heterozygosity or RAD54 in high copy. Mol. Cell. Biol. 22:6336–43 Morrow DM, Connelly C, Hieter P. 1997. “Break copy” duplication: a model for chromosome fragment formation in Saccharomyces cerevisiae. Genetics 147:371–82 Mortensen UH, Bendixen C, Sunjevaric I, Rothstein R. 1996. DNA strand annealing is promoted by the yeast Rad52 protein. Proc. Natl. Acad. Sci. USA 93: 10729–34 Mortensen UH, Erdeniz N, Feng Q, Rothstein R. 2002. A molecular genetic dissection of the evolutionarily conserved N terminus of yeast Rad52. Genetics 161:549–62 Mullen JR, Kaliraman V, Ibrahim SS, Brill SJ. 2001. Requirement for three novel protein complexes in the absence of the Sgs1 DNA helicase in Saccharomyces cerevisiae. Genetics 157:103– 18 Nairz K, Klein F. 1997. mre11S a yeast mutation that blocks double-strandbreak processing and permits nonhomologous synapsis in meiosis. Genes Dev. 11:2272–90 Nassif N, Penney J, Pal S, Engels WR, Gloor GB. 1994. Efficient copying of nonhomologous sequences from ectopic sites via P-element-induced gap repair. Mol. Cell Biol. 14:1613–25 New JH, Sugiyama T, Zaitseva E, Kowalczykowski SC. 1998. Rad52 protein stimulates DNA strand exchange by 141. 142. 143. 144. 145. 146. 147. 148. 149. 150. 151. Rad51 and replication protein A. Nature 391:407–10 Nickoloff JA, Chen EY, Heffron F. 1986. A 24-base-pair DNA sequence from the MAT locus stimulates intergenic recombination in yeast. Proc. Natl. Acad. Sci. USA 83:7831–35 Ogawa T, Yu X, Shinohara A, Egelman EH. 1993. Similarity of the yeast RAD51 filament to the bacterial RecA filament. Science 259:1896–99 Ooi SL, Shoemaker DD, Boeke JD. 2001. A DNA microarray-based genetic screen for nonhomologous end-joining mutants in Saccharomyces cerevisiae. Science 294:2552–56 Orr-Weaver TL, Szostak JW. 1983. Yeast recombination: the association between double-strand gap repair and crossing-over. Proc. Natl. Acad. Sci. USA 80:4417–21 Orr-Weaver TL, Szostak JW, Rothstein RJ. 1981. Yeast transformation: a model system for the study of recombination. Proc. Natl. Acad. Sci. USA 78:6354–58 Osman F, Dixon J, Doe CL, Whitby MC. 2003. Generating crossovers by resolution of nicked Holliday junctions: a role for Mus81-Eme1 in meiosis. Mol. Cell 12:761–74 Pages V, Fuchs RP. 2003. Uncoupling of leading- and lagging-strand DNA replication during lesion bypass in vivo. Science 300:1300–3 Passy SI, Yu X, Li Z, Radding CM, Masson JY, et al. 1999. Human Dmc1 protein binds DNA as an octameric ring. Proc. Natl. Acad. Sci. USA 96:10684–88 Paull TT, Gellert M. 1998. The 3 to 5 exonuclease activity of Mre11 facilitates repair of DNA double-strand breaks. Mol. Cell 1:969–79 Paull TT, Gellert M. 1999. Nbs1 potentiates ATP-driven DNA unwinding and endonuclease cleavage by the Mre11/Rad50 complex. Genes Dev. 13: 1276–88 Pellegrini L, Yu DS, Lo T, Anand S, Lee 15 Oct 2004 19:50 AR AR230-GE38-08.tex AR230-GE38-08.sgm LaTeX2e(2002/01/18) RECOMBINATION PROTEINS IN YEAST 152. Annu. Rev. Genet. 2004.38:233-271. Downloaded from arjournals.annualreviews.org by UNIVERSITY OF SOUTH CAROL on 02/10/05. For personal use only. 153. 154. 155. 156. 157. 158. 159. 160. M, et al. 2002. Insights into DNA recombination from the structure of a RAD51BRCA2 complex. Nature 420:287–93 Petukhova G, Stratton S, Sung P. 1998. Catalysis of homologous DNA pairing by yeast Rad51 and Rad54 proteins. Nature 393:91–94 Petukhova G, Stratton SA, Sung P. 1999. Single-strand DNA binding and annealing activities in the yeast recombination factor Rad59. J. Biol. Chem. 274:33839– 42 Petukhova G, Sung P, Klein H. 2000. Promotion of Rad51-dependent D-loop formation by yeast recombination factor Rdh54/Tid1. Genes Dev. 14:2206– 15 Petukhova G, Van Komen S, Vergano S, Klein H, Sung P. 1999. Yeast Rad54 promotes Rad51-dependent homologous DNA pairing via ATP hydrolysis-driven change in DNA double helix conformation. J. Biol. Chem. 274:29453–62 Plessis A, Perrin A, Haber JE, Dujon B. 1992. Site-specific recombination determined by I-SceI, a mitochondrial group I intron-encoded endonuclease expressed in the yeast nucleus. Genetics 130:451– 60 Prakash L, Prakash S. 1977. Isolation and characterization of MMS-sensitive mutants of Saccharomyces cerevisiae. Genetics 86:33–55 Prinz S, Amon A, Klein F. 1997. Isolation of COM1, a new gene required to complete meiotic double-strand breakinduced recombination in Saccharomyces cerevisiae. Genetics 146:781–95 Ranatunga W, Jackson D, Lloyd JA, Forget AL, Knight KL, Borgstahl GE. 2001. Human RAD52 exhibits two modes of self-association. J. Biol. Chem. 276:15876–80 Rattray AJ, McGill CB, Shafer BK, Strathern JN. 2001. Fidelity of mitotic double-strand-break repair in Saccharomyces cerevisiae. A role for sae2/com1. Genetics 158:109–22 P1: GCE 267 161. Rattray AJ, Symington LS. 1994. Use of a chromosomal inverted repeat to demonstrate that the RAD51 and RAD52 genes of Saccharomyces cerevisiae have different roles in mitotic recombination. Genetics 138:587–95 162. Raymond WE, Kleckner N. 1993. RAD50 protein of S. cerevisiae exhibits ATP-dependent DNA binding. Nucleic Acids Res. 21:3851–56 163. Rong L, Klein HL. 1993. Purification and characterization of the SRS2 DNA helicase of the yeast Saccharomyces cerevisiae. J. Biol. Chem. 268:1252–59 164. Roth DB, Wilson JH. 1986. Nonhomologous recombination in mammalian cells: role for short sequence homologies in the joining reaction. Mol. Cell Biol. 6:4295– 304 165. Schar P, Herrmann G, Daly G, Lindahl T. 1997. A newly identified DNA ligase of Saccharomyces cerevisiae involved in RAD52-independent repair of DNA double-strand breaks. Genes Dev. 11:1912–24 166. Schiestl RH, Petes TD. 1991. Integration of DNA fragments by illegitimate recombination in Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 88: 7585–89 167. Schiestl RH, Prakash S, Prakash L. 1990. The SRS2 suppressor of rad6 mutations of Saccharomyces cerevisiae acts by channeling DNA lesions into the RAD52 DNA repair pathway. Genetics 124:817– 31 168. Schwacha A, Kleckner N. 1995. Identification of double Holliday junctions as intermediates in meiotic recombination. Cell 83:783–91 169. Schwacha A, Kleckner N. 1997. Interhomolog bias during meiotic recombination: meiotic functions promote a highly differentiated interhomolog-only pathway. Cell 90:1123–35 169a. Sehorn MG, Sigurdsson S, Bussen W, Unger VM, Sung P. 2004. Human meiotic recombinase Dmc1 promotes 15 Oct 2004 19:50 268 170. Annu. Rev. Genet. 2004.38:233-271. Downloaded from arjournals.annualreviews.org by UNIVERSITY OF SOUTH CAROL on 02/10/05. For personal use only. 171. 172. 173. 174. 175. 176. 177. 178. 179. AR KROGH AR230-GE38-08.tex AR230-GE38-08.sgm LaTeX2e(2002/01/18) P1: GCE SYMINGTON ATP-dependent homologous DNA strand exchange. Nature 429:433–37 Seigneur M, Bidnenko V, Ehrlich SD, Michel B. 1998. RuvAB acts at arrested replication forks. Cell 95:419–30 Sharples GJ, Leach DR. 1995. Structural and functional similarities between the SbcCD proteins of Escherichia coli and the RAD50 and MRE11 (RAD32) recombination and repair proteins of yeast. Mol. Microbiol. 17:1215–17 Shin DS, Pellegrini L, Daniels DS, Yelent B, Craig L, et al. 2003. Full-length archaeal Rad51 structure and mutants: mechanisms for RAD51 assembly and control by BRCA2. EMBO J. 22:4566– 76 Shinohara A, Gasior S, Ogawa T, Kleckner N, Bishop DK. 1997. Saccharomyces cerevisiae recA homologues RAD51 and DMC1 have both distinct and overlapping roles in meiotic recombination. Genes Cells 2:615–29 Shinohara A, Ogawa H, Ogawa T. 1992. Rad51 protein involved in repair and recombination in S. cerevisiae is a RecAlike protein. Cell 69:457–70. Erratum. 1992. Cell 71(1):180 Shinohara A, Ogawa T. 1998. Stimulation by Rad52 of yeast Rad51-mediated recombination. Nature 391:404–7 Shinohara A, Shinohara M, Ohta T, Matsuda S, Ogawa T. 1998. Rad52 forms ring structures and co-operates with RPA in single-strand DNA annealing. Genes Cells 3:145–56 Singleton MR, Wentzell LM, Liu Y, West SC, Wigley DB. 2002. Structure of the single-strand annealing domain of human RAD52 protein. Proc. Natl. Acad. Sci. USA 99:13492–97 Smith GR, Boddy MN, Shanahan P, Russell P. 2003. Fission yeast Mus81.Eme1 Holliday junction resolvase is required for meiotic crossing over but not for gene conversion. Genetics 165:2289–93 Smith J, Rothstein R. 1999. An allele of RFA1 suppresses RAD52-dependent 180. 181. 182. 183. 184. 185. 186. 187. 188. 189. double-strand break repair in Saccharomyces cerevisiae. Genetics 151:447– 58 Solinger JA, Heyer WD. 2001. Rad54 protein stimulates the postsynaptic phase of Rad51 protein-mediated DNA strand exchange. Proc. Natl. Acad. Sci. USA 98:8447–53 Solinger JA, Kiianitsa K, Heyer WD. 2002. Rad54, a Swi2/Snf2-like recombinational repair protein, disassembles Rad51:dsDNA filaments. Mol. Cell 10:1175–88 Song B, Sung P. 2000. Functional interactions among yeast Rad51 recombinase, Rad52 mediator, and replication protein A in DNA strand exchange. J. Biol. Chem. 275:15895–904 Sonoda E, Sasaki MS, Buerstedde JM, Bezzubova O, Shinohara A, et al. 1998. Rad51-deficient vertebrate cells accumulate chromosomal breaks prior to cell death. EMBO J. 17:598–608 Spell RM, Jinks-Robertson S. 2003. Role of mismatch repair in the fidelity of RAD51- and RAD59-dependent recombination in Saccharomyces cerevisiae. Genetics 165:1733–44 Stasiak AZ, Larquet E, Stasiak A, Muller S, Engel A, et al. 2000. The human Rad52 protein exists as a heptameric ring. Curr. Biol. 10:337–40 Stellwagen AE, Haimberger ZW, Veatch JR, Gottschling DE. 2003. Ku interacts with telomerase RNA to promote telomere addition at native and broken chromosome ends. Genes Dev. 17:2384–95 Story RM, Weber IT, Steitz TA. 1992. The structure of the E. coli recA protein monomer and polymer. Nature 355:318– 25 Stracker TH, Carson CT, Weitzman MD. 2002. Adenovirus oncoproteins inactivate the Mre11-Rad50-Nbs1 DNA repair complex. Nature 418:348–52 Strathern JN, Klar AJ, Hicks JB, Abraham JA, Ivy JM, et al. 1982. Homothallic switching of yeast mating type cassettes 15 Oct 2004 19:50 AR AR230-GE38-08.tex AR230-GE38-08.sgm LaTeX2e(2002/01/18) RECOMBINATION PROTEINS IN YEAST 190. Annu. Rev. Genet. 2004.38:233-271. Downloaded from arjournals.annualreviews.org by UNIVERSITY OF SOUTH CAROL on 02/10/05. For personal use only. 191. 192. 193. 194. 195. 196. 197. 198. is initiated by a double-stranded cut in the MAT locus. Cell 31:183–92 Strunnikov AV, Jessberger R. 1999. Structural maintenance of chromosomes (SMC) proteins: conserved molecular properties for multiple biological functions. Eur. J. Biochem. 263:6–13 Sugawara N, Haber JE. 1992. Characterization of double-strand break-induced recombination: homology requirements and single-stranded DNA formation. Mol. Cell Biol. 12:563–75 Sugawara N, Ira G, Haber JE. 2000. DNA length dependence of the singlestrand annealing pathway and the role of Saccharomyces cerevisiae RAD59 in double-strand break repair. Mol. Cell Biol. 20:5300–9 Sugawara N, Wang X, Haber JE. 2003. In vivo roles of Rad52, Rad54, and Rad55 proteins in Rad51-mediated recombination. Mol. Cell 12:209–19 Sugiyama T, Kowalczykowski SC. 2002. Rad52 protein associates with replication protein A (RPA)-single-stranded DNA to accelerate Rad51-mediated displacement of RPA and presynaptic complex formation. J. Biol. Chem. 277:31663–72 Sugiyama T, New JH, Kowalczykowski SC. 1998. DNA annealing by RAD52 protein is stimulated by specific interaction with the complex of replication protein A and single-stranded DNA. Proc. Natl. Acad. Sci. USA 95:6049– 54 Sun H, Treco D, Schultes NP, Szostak JW. 1989. Double-strand breaks at an initiation site for meiotic gene conversion. Nature 338:87–90 Sun H, Treco D, Szostak JW. 1991. Extensive 3 -overhanging, single-stranded DNA associated with the meiosisspecific double-strand breaks at the ARG4 recombination initiation site. Cell 64:1155–61 Sung P. 1997. Function of yeast Rad52 protein as a mediator between replication 199. 200. 201. 202. 203. 204. 205. 206. 207. 208. P1: GCE 269 protein A and the Rad51 recombinase. J. Biol. Chem. 272:28194–97 Sung P. 1997. Yeast Rad55 and Rad57 proteins form a heterodimer that functions with replication protein A to promote DNA strand exchange by Rad51 recombinase. Genes Dev. 11:1111–21 Sung P, Robberson DL. 1995. DNA strand exchange mediated by a RAD51ssDNA nucleoprotein filament with polarity opposite to that of RecA. Cell 82:453–61 Sung P, Stratton SA. 1996. Yeast Rad51 recombinase mediates polar DNA strand exchange in the absence of ATP hydrolysis. J. Biol. Chem. 271:27983–86 Symington LS. 2002. Role of RAD52 epistasis group genes in homologous recombination and double-strand break repair. Microbiol. Mol. Biol. Rev. 66:630– 70 Szostak JW, Orr-Weaver TL, Rothstein RJ, Stahl FW. 1983. The double-strandbreak repair model for recombination. Cell 33:25–35 Teo SH, Jackson SP. 1997. Identification of Saccharomyces cerevisiae DNA ligase IV: involvement in DNA double-strand break repair. EMBO J. 16:4788–95 Teo SH, Jackson SP. 2000. Lif1p targets the DNA ligase Lig4p to sites of DNA double-strand breaks. Curr. Biol. 10:165–68 Trujillo KM, Roh DH, Chen L, Van Komen S, Tomkinson A, Sung P. 2003. Yeast Xrs2 binds DNA and helps target Rad50 and Mre11 to DNA ends. J. Biol. Chem. 278:48957–64 Trujillo KM, Sung P. 2001. DNA structure-specific nuclease activities in the Saccharomyces cerevisiae Rad50 Mre11 complex. J. Biol. Chem. 276: 35458–64 Trujillo KM, Yuan SS, Lee EY, Sung P. 1998. Nuclease activities in a complex of human recombination and DNA repair factors Rad50, Mre11, and p95. J. Biol. Chem. 273:21447–50 15 Oct 2004 19:50 Annu. Rev. Genet. 2004.38:233-271. Downloaded from arjournals.annualreviews.org by UNIVERSITY OF SOUTH CAROL on 02/10/05. For personal use only. 270 AR KROGH AR230-GE38-08.tex AR230-GE38-08.sgm LaTeX2e(2002/01/18) P1: GCE SYMINGTON 209. Tsubouchi H, Ogawa H. 1998. A novel mre11 mutation impairs processing of double-strand breaks of DNA during both mitosis and meiosis. Mol. Cell Biol. 18:260–68 210. Tsubouchi H, Ogawa H. 2000. Exo1 roles for repair of DNA double-strand breaks and meiotic crossing over in Saccharomyces cerevisiae. Mol. Biol. Cell 11:2221–33 211. Tsubouchi H, Roeder GS. 2003. The importance of genetic recombination for fidelity of chromosome pairing in meiosis. Dev. Cell 5:915–25 212. Tsukamoto M, Yamashita K, Miyazaki T, Shinohara M, Shinohara A. 2003. The N-terminal DNA-binding domain of Rad52 promotes RAD51-independent recombination in Saccharomyces cerevisiae. Genetics 165:1703–15 213. Tsukamoto Y, Taggart AK, Zakian VA. 2001. The role of the Mre11-Rad50Xrs2 complex in telomerase-mediated lengthening of Saccharomyces cerevisiae telomeres. Curr. Biol. 11:1328–35 214. Tsuzuki T, Fujii Y, Sakumi K, Tominaga Y, Nakao K, et al. 1996. Targeted disruption of the Rad51 gene leads to lethality in embryonic mice. Proc. Natl. Acad. Sci. USA 93:6236–40 214a. Usui T, Ogawa H, Petrini JH. 2001. A DNA damage response pathway controlled by Tel1 and the Mre11 complex. Mol. Cell 7:1255–66 215. Usui T, Ohta T, Oshiumi H, Tomizawa J, Ogawa H, Ogawa T. 1998. Complex formation and functional versatility of Mre11 of budding yeast in recombination. Cell 95:705–16 216. Valencia M, Bentele M, Vaze MB, Herrmann G, Kraus E, et al. 2001. NEJ1 controls non-homologous end joining in Saccharomyces cerevisiae. Nature 414:666–69 217. Van Komen S, Petukhova G, Sigurdsson S, Stratton S, Sung P. 2000. Superhelicity-driven homologous DNA pairing by yeast recombination factors 218. 219. 220. 221. 222. 223. 224. 225. 226. Rad51 and Rad54. Mol. Cell. 6:563– 72 Van Komen S, Reddy MS, Krejci L, Klein H, Sung P. 2003. ATPase and DNA helicase activities of the Saccharomyces cerevisiae anti-recombinase Srs2. J. Biol. Chem. 278:44331–37 van Noort J, van Der Heijden T, de Jager M, Wyman C, Kanaar R, Dekker C. 2003. The coiled-coil of the human Rad50 DNA repair protein contains specific segments of increased flexibility. Proc. Natl. Acad. Sci. USA 100:7581–86 Varon R, Vissinga C, Platzer M, Cerosaletti KM, Chrzanowska KH, et al. 1998. Nibrin, a novel DNA doublestrand break repair protein, is mutated in Nijmegen breakage syndrome. Cell 93:467–76 Veaute X, Jeusset J, Soustelle C, Kowalczykowski SC, Le Cam E, Fabre F. 2003. The Srs2 helicase prevents recombination by disrupting Rad51 nucleoprotein filaments. Nature 423:309–12 Wallis JW, Chrebet G, Brodsky G, Rolfe M, Rothstein R. 1989. A hyperrecombination mutation in S. cerevisiae identifies a novel eukaryotic topoisomerase. Cell 58:409–19 Wan L, de los Santos T, Zhang C, Shokat K, Hollingsworth NM. 2004. Mek1 kinase activity functions downstream of RED1 in the regulation of meiotic double-strand break repair in budding yeast. Mol. Biol. Cell 15:11–23 Wang H, Perrault AR, Takeda Y, Qin W, Iliakis G. 2003. Biochemical evidence for Ku-independent backup pathways of NHEJ. Nucleic Acids Res. 31:5377–88 Wang X, Haber JE. 2004. Role of Saccharomyces single-stranded DNAbinding protein RPA in the strand invasion step of double-strand break repair. PLoS Biol. 2:E21 Weiden MD, Ginsberg HS. 1994. Deletion of the E4 region of the genome produces adenovirus DNA concatemers. Proc. Natl. Acad. Sci. USA 91:153–57 15 Oct 2004 19:50 AR AR230-GE38-08.tex AR230-GE38-08.sgm LaTeX2e(2002/01/18) Annu. Rev. Genet. 2004.38:233-271. Downloaded from arjournals.annualreviews.org by UNIVERSITY OF SOUTH CAROL on 02/10/05. For personal use only. RECOMBINATION PROTEINS IN YEAST 227. White CI, Haber JE. 1990. Intermediates of recombination during mating type switching in Saccharomyces cerevisiae. EMBO J. 9:663–73 228. Wilson S, Warr N, Taylor DL, Watts FZ. 1999. The role of Schizosaccharomyces pombe Rad32, the Mre11 homologue, and other DNA damage response proteins in non-homologous end joining and telomere length maintenance. Nucleic Acids Res. 27:2655–61 229. Wilson TE, Grawunder U, Lieber MR. 1997. Yeast DNA ligase IV mediates non-homologous DNA end joining. Nature 388:495–98 230. Wolner B, van Komen S, Sung P, Peterson CL. 2003. Recruitment of the recombinational repair machinery to a DNA double-strand break in yeast. Mol. Cell. 12:221–32 231. Wu L, Hickson ID. 2003. The Bloom’s syndrome helicase suppresses crossing over during homologous recombination. Nature 426:870–74 232. Wu TC, Lichten M. 1994. Meiosis- 233. 234. 235. 236. P1: GCE 271 induced double-strand break sites determined by yeast chromatin structure. Science 263:515–18 Wu X, Ranganathan V, Weisman DS, Heine WF, Ciccone DN, et al. 2000. ATM phosphorylation of Nijmegen breakage syndrome protein is required in a DNA damage response. Nature 405:477–82 Yang S, Yu X, Seitz EM, Kowalczykowski SC, Egelman EH. 2001. Archaeal RadA protein binds DNA as both helical filaments and octameric rings. J. Mol. Biol. 314:1077–85 Yu J, Marshall K, Yamaguchi M, Haber JE, Weil CF. 2004. Microhomologydependent end joining and repair of transposon-induced DNA hairpins by host factors in Saccharomyces cerevisiae. Mol. Cell Biol. 24:1351–64 Zhao S, Renthal W, Lee EY. 2002. Functional analysis of FHA and BRCT domains of NBS1 in chromatin association and DNA damage responses. Nucleic Acids Res. 30:4815–22 P1: JRX October 18, 2004 14:55 Annual Reviews AR230-FM Annual Review of Genetics Volume 38, 2004 Annu. Rev. Genet. 2004.38:233-271. Downloaded from arjournals.annualreviews.org by UNIVERSITY OF SOUTH CAROL on 02/10/05. For personal use only. CONTENTS MOBILE GROUP II INTRONS, Alan M. Lambowitz and Steven Zimmerly THE GENETICS OF MAIZE EVOLUTION, John Doebley GENETIC CONTROL OF RETROVIRUS SUSCEPTIBILITY IN MAMMALIAN CELLS, Stephen P. Goff LIGHT SIGNAL TRANSDUCTION IN HIGHER PLANTS, Meng Chen, Joanne Chory, and Christian Fankhauser 1 37 61 87 CHLAMYDOMONAS REINHARDTII IN THE LANDSCAPE OF PIGMENTS, Arthur R. Grossman, Martin Lohr, and Chung Soon Im THE GENETICS OF GEOCHEMISTRY, Laura R. Croal, Jeffrey A. Gralnick, Davin Malasarn, and Dianne K. Newman CLOSING MITOSIS: THE FUNCTIONS OF THE CDC14 PHOSPHATASE AND ITS REGULATION, Frank Stegmeier and Angelika Amon RECOMBINATION PROTEINS IN YEAST, Berit Olsen Krogh and Lorraine S. Symington 119 175 203 233 DEVELOPMENTAL GENE AMPLIFICATION AND ORIGIN REGULATION, John Tower 273 THE FUNCTION OF NUCLEAR ARCHITECTURE: A GENETIC APPROACH, Angela Taddei, Florence Hediger, Frank R. Neumann, and Susan M. Gasser 305 GENETIC MODELS IN PATHOGENESIS, Elizabeth Pradel and Jonathan J. Ewbank 347 MELANOCYTES AND THE MICROPHTHALMIA TRANSCRIPTION FACTOR NETWORK, Eirı́kur Steingrı́msson, Neal G. Copeland, and Nancy A. Jenkins EPIGENETIC REGULATION OF CELLULAR MEMORY BY THE POLYCOMB AND TRITHORAX GROUP PROTEINS, Leonie Ringrose and Renato Paro REPAIR AND GENETIC CONSEQUENCES OF ENDOGENOUS DNA BASE DAMAGE IN MAMMALIAN CELLS, Deborah E. Barnes and Tomas Lindahl 365 413 445 MITOCHONDRIA OF PROTISTS, Michael W. Gray, B. Franz Lang, and Gertraud Burger 477 v P1: JRX October 18, 2004 vi 14:55 Annual Reviews AR230-FM CONTENTS METAGENOMICS: GENOMIC ANALYSIS OF MICROBIAL COMMUNITIES, Christian S. Riesenfeld, Patrick D. Schloss, and Jo Handelsman 525 GENOMIC IMPRINTING AND KINSHIP: HOW GOOD IS THE EVIDENCE?, David Haig 553 MECHANISMS OF PATTERN FORMATION IN PLANT EMBRYOGENESIS, Viola Willemsen and Ben Scheres Annu. Rev. Genet. 2004.38:233-271. Downloaded from arjournals.annualreviews.org by UNIVERSITY OF SOUTH CAROL on 02/10/05. For personal use only. DUPLICATION AND DIVERGENCE: THE EVOLUTION OF NEW GENES AND OLD IDEAS, John S. Taylor and Jeroen Raes GENETIC ANALYSES FROM ANCIENT DNA, Svante Pääbo, Hendrik Poinar, David Serre, Viviane Jaenicke-Despres, Juliane Hebler, Nadin Rohland, Melanie Kuch, Johannes Krause, Linda Vigilant, and Michael Hofreiter 587 615 645 PRION GENETICS: NEW RULES FOR A NEW KIND OF GENE, Reed B. Wickner, Herman K. Edskes, Eric D. Ross, Michael M. Pierce, Ulrich Baxa, Andreas Brachmann, and Frank Shewmaker 681 PROTEOLYSIS AS A REGULATORY MECHANISM, Michael Ehrmann and Tim Clausen 709 MECHANISMS OF MAP KINASE SIGNALING SPECIFICITY IN SACCHAROMYCES CEREVISIAE, Monica A. Schwartz and Hiten D. Madhani 725 rRNA TRANSCRIPTION IN ESCHERICHIA COLI, Brian J. Paul, Wilma Ross, Tamas Gaal, and Richard L. Gourse 749 COMPARATIVE GENOMIC STRUCTURE OF PROKARYOTES, Stephen D. Bentley and Julian Parkhill 771 SPECIES SPECIFICITY IN POLLEN-PISTIL INTERACTIONS, Robert Swanson, Anna F. Edlund, and Daphne Preuss 793 INTEGRATION OF ADENO-ASSOCIATED VIRUS (AAV) AND RECOMBINANT AAV VECTORS, Douglas M. McCarty, Samuel M. Young Jr., and Richard J. Samulski 819 INDEXES Subject Index ERRATA An online log of corrections to Annual Review of Genetics chapters may be found at http://genet.annualreviews.org/errata.shtml 847