Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
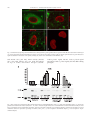
Cardiovascular Research 63 (2004) 176 – 185 www.elsevier.com/locate/cardiores Influence of the 4G/5G PAI-1 genotype on angiotensin II-stimulated human endothelial cells and in patients with hypertension C. Roncal a,b, J. Orbe a,b, J.A. Rodriguez a,b, M. Belzunce a,b, O. Beloqui c, J. Diez b, J.A. Páramo a,b,* a Atherosclerosis Research Laboratory, School of Medicine, University of Navarra, 31080-Pamplona, Spain b Division of Cardiovascular Pathophysiology, Foundation for Applied Medical Research (FIMA), School of Medicine, University of Navarra, Pamplona, Spain c University Hospital, University of Navarra, Pamplona, Spain Received 1 December 2003; received in revised form 11 March 2004; accepted 18 March 2004 Available online 14 May 2004 Time for primary review 28 days Abstract Background: We examined the influence of the 4G/5G PAI-1 (plasminogen activator inhibitor) genotype on Angiotensin II (Ang II)induced PAI-1 expression by human endothelial cells (HUVEC) in the presence and absence of AT1-receptor blocker losartan, and screened for this polymorphism in relation to plasma PAI-1 and arterial pressure in apparently healthy subjects. Methods and results: Genotyped cultured HUVEC were incubated with Ang II (10 8 M) with or without losartan up to 24 h. PAI-1 mRNA was determined in cell extracts and protein and activity assessed in supernatants and extracellular matrix (ECM). Ang II increased PAI-1 mRNA and activity in a genotypedependent manner, higher values observed for 4G/4G HUVEC compared with 4G/5G and 5G/5G genotypes ( p < 0.05). Laser confocal microscopy and Western blot analysis showed increased PAI-1 protein within ECM in Ang II-stimulated cultures. PAI-1 expression and protein secretion induced by Ang II in 4G/4G HUVEC was completely inhibited by preincubation with 0.05 AM losartan ( p < 0.01), indicating an AT1-mediated effect. In a group of hypertensives homozygous for the 4G allele, PAI-1 antigen was significantly increased (51.0 F 10.1 ng/ml) compared with normotensives (28.3 F 4.0 ng/ml) and hypertensives carrying the 5G allele ( p < 0.05). Conclusions: The 4G/5G PAI-1 polymorphism determines the endothelial PAI-1 upregulation by Ang II and the inhibitory response to losartan. Analysis of PAI-1 genotypes may help identifying subgroups of hypertensives at higher cardiovascular risk. D 2004 European Society of Cardiology. Published by Elsevier B.V. All rights reserved. Keywords: PAI-1; Atherosclerosis; Polymorphism; Endothelium; Angiotensin-II; Arterial hypertension 1. Introduction Activation of the renin – angiotensin– aldosterone system (RAAS) and defective fibrinolysis has been linked with an increased thrombotic risk and fibrosis [1,2]. There is strong evidence that the fibrinolytic balance is largely under the control of the RAAS [3]. Angiotensin II (Ang II), a key component of this system, plays a pivotal role in the * Corresponding author. Atherosclerosis Research Laboratory, School of Medicine, University of Navarra, 31080-Pamplona, Spain. Tel.: +34948296397; fax: +34-948296500. E-mail address: [email protected] (J.A. Páramo). homeostasis of systemic arterial pressure, and also modulates fibrinolysis by increasing type 1 plasminogen activator inhibitor (PAI-1) in vitro and in vivo [4 –6]. PAI-1 is the primary inhibitor of tissue-type plasminogen activator (tPA) and regulates fibrinolytic system [7]. A crucial role of PAI-1 in atherothrombosis has been demonstrated both at experimental and clinical levels [8– 10]. Endothelial cells in general, and human umbilical vein endothelial cells (HUVEC) in particular, express PAI-1 [11,12]. Transcriptional control of PAI-1 promoter in response to cytokine and hormone stimuli is an important regulatory mechanism to fibrinolysis. The PAI-1 4G/5G polymorphism located 675 bp upstream of the start of transcription has been associated with increased plasma 0008-6363/$ - see front matter D 2004 European Society of Cardiology. Published by Elsevier B.V. All rights reserved. doi:10.1016/j.cardiores.2004.03.023 C. Roncal et al. / Cardiovascular Research 63 (2004) 176–185 PAI-1 concentrations and with increased risk for coronary disease [13 – 15]. It has been shown that proatherogenic stimuli such as IL-1 and VLDL induced PAI-1 synthesis in cells containing the 4G sequence [13,16], this effect being explained through the binding of a transcriptional activator to both 4G and 5G alleles, while the 5G allele also binds a transcriptional repressor [13]. Whereas Ang II causes the release of PAI-1 by endothelial cells [5,6], no studies have addressed the issue of whether those effects differ according to the 4G/5G endothelial cell genotype. Recent reports have also demonstrated that the relationship between Ang II and PAI-1 would be mediated by the angiotensin II type 1 (AT1) receptor [17,18]; furthermore, AT1 receptor blockers have been shown to attenuate angiotensin-induced inhibition of fibrinolysis and this may be one of the mechanisms in the prevention of ischemic events [19]. In addition, AT1 receptor blockade has been shown to slow the progression of vascular sclerosis via its effect on PAI-1 [20]. Because endothelial PAI-1 is an important source for PAI1 levels in plasma and there is evidence that genetic factors may be important in determining inhibitor levels, we investigated whether the PAI-1 response by endothelial cells challenged with Ang II is related to the 4G/5G genotype, and whether selective AT1 receptor blockade would lower endothelial cell production of PAI-1 in a genotype-specific manner. In addition, the 4G/5G polymorphism and plasma PAI-1 levels were also determined in apparently healthy subjects in relation to their systemic arterial pressure. 2. Materials and methods 2.1. Endothelial cell culture Endothelial cells (HUVEC) were isolated from human umbilical cords obtained less than 8 h after delivery, by digestion with collagenase A (Gibco) as previously described [11]. 2.2. Experimental design All studies were performed with confluent cultures in the second passage. Cultures were washed with HBSS (Gibco) after which fresh serum-free media (Gibco) supplemented with penicillin/streptomycin was added. Cultures were challenged with Angiotensin II (Ang II, Sigma) 10 8 M alone, or in the presence of AT1 receptor blocker losartan (0.01 and 0.05 AM, kindly provided by Merck, USA), preincubated 18 h, and supernatants and cells were harvested at 0, 2, 6 and 24 h after the start of the experiment. Dose of Ang II was chosen according to previous dose –response assays (10 6 to 10 10 M, data not shown). Unstimulated cultures were used as controls. Endothelial cells derived from three individuals per genotype were used for each experimental condition. 177 2.3. Subjects The population studied consisted of 122 consecutive subjects attending the Vascular Risk Area of a single institution (University Clinic of Navarra) for global vascular risk assessment. Blood pressure was measured twice on the right upper arm with a random-zero mercury sphygmomanometer in patients in the sitting position (average of two measurements). Forty six untreated subjects had elevated systolic and/or diastolic blood pressure (>139/89 mm Hg) (hypertensive group) and 76 age-matched normotensive subjects were used as control group. Body mass index (BMI) was calculated as weight/height2 (kg/m2) and used as an estimate of overall adiposity. Written informed consent was obtained before participation in the study, and the local committee on human research approved the study protocol. The study was performed in accordance with the principles of the Declaration of Helsinki. 2.4. Laboratory analysis Blood samples were collected on ice and centrifuged immediately at 4 jC for 20 min. All plasma or serum was separated and stored at 80 jC until assayed. Serum C-reactive protein was assayed by hs-CRP immunoassay system (Immulyte hs-CRP, Diagnostic Product). Serum aldosterone was measured using a commercially available radioimmunoassay kit (Aldoctk-2, DiaSorin). The intra- and inter-assay variation coefficients were 4% and 6%, respectively. Serum cholesterol, HDL-cholesterol, LDL-cholesterol, triglycerides and glucose were measured on fasting blood samples by standard laboratory techniques. 2.5. PAI-1 promoter 4G/5G genotype Genomic DNA was extracted from cord tissue samples or subject blood samples using the TriPure Isolation Reagent (Roche) following the manufacturer’s instructions. The PAI1 promoter 4G/5G polymorphism was analyzed with an allele-specific PCR [21]. 2.6. RNA isolation and real-time quantitative RT-PCR Total RNA was extracted from cells with TriPure (Roche) and quantified with ultraviolet spectroscopy. Total RNA (0.5 Ag) was reverse transcribed with random primers with MMLV reverse transcriptase (Invitrogen), in the presence of RNase Outk (Invitrogen). One hundred nanograms of reverse-transcribed RNA was primed with specific oligonucleotides for PAI-1 (5VCCGTCTGATTTGTGGAAGAGG-3V and 5V-ACAGGAGGAGAAACCCAGCAG-3V), and h-actin (5V-AGCCTCGCCTTTGCCGA-3V and 5V-CTGGTGCCTGGGGCG-3V). PCR was performed as previously described [22] using 178 C. Roncal et al. / Cardiovascular Research 63 (2004) 176–185 SYBR Green PCR Master Mix (Applied Biosystems) on the ABI PRISM 7000 Detection System (Applied Biosystems). Potential genomic DNA contamination was excluded by using intron-encompassing primers. All samples were assayed in triplicate and normalized on the basis of their h-actin content. A melting curve analysis and length verification by gel electroforesis were carried out to confirm the specificity of PCR products. 2.10. Statistical analysis Results are expressed as mean F S.E.M. of three independent experiments. For statistical purposes, all data at 2, 6 and 24 h were expressed as percentages of their baseline value, which was considered 100%. Statistical analysis was performed by ANOVA or Kruskal – Wallis followed by Tukey-b post-hoc test or Mann – Whitney U-test for comparisons between genotypes. Differences between stimulated and 2.7. Determination of PAI-1 activity and antigen Active PAI-1 was measured in HUVEC supernatants by an amidolytic assay (Coatest-PAI, Chromogenix) as previously described [23]. PAI-1 antigen levels in culture supernatants and plasma samples were measured, by ELISA (Asserachrom-PAI-1, Diagnostica Stago) according to manufacturer instructions. 2.8. Laser confocal immunofluorescence microscopy of HUVEC Cells grown on slides (Nalgene Nunc International) precoated with gelatin 0.1% were fixed with 4% paraformaldehyde and permeabilized with cold methanol/acetone (v/v) before immunocytochemistry. Briefly, after incubating HUVEC with monoclonal anti-PAI-1 (American Diagnostica, 20 Ag/ml) overnight at 4 jC, the slides were washed and incubated for 1 h at room temperature with 10 Ag/ml Alexa Fluor 488R anti-mouse IgG (Molecular Probes). TO-PROR3 (1.25 AM; Molecular Probes) was used for nuclear counterstaining. Finally, cells were mounted in SlowFadeR Antifade reagent (Molecular Probes) and visualized under confocal laser scan microscope (Zeiss LSM-510 Meta; 63 objective). Forty image stacks (512 512 pixels each) were captured from representative areas. The specificity of the fluorescent signal was checked in samples prepared by omitting the first antibody. 2.9. PAI-1 protein in ECM Genotyped HUVEC monolayers (n = 3) were washed with cold PBS and incubated at 37 jC with 0.5% Triton X-100 in PBS for 10 min. The supernatant was discarded and the remaining ECM was solubilized with electrophoresis (PAGE) sample buffer as previously described [24]. Samples from ECM were separated by SDS-PAGE under reducing conditions in Tris – glycine –SDS buffer (pH 8.3). Electrotransfer was performed in Tris – glycine – methanol buffer for 1 h at 300 mA. Nitrocellulose membrane was blocked for 1 h, then incubated with murine anti-PAI-1 (1 Ag/ ml, American Diagnostica) or rabbit anti-vitronectin (8 Ag/ ml, American Diagnostica) in PBS – Tween (0.1%). The membrane was rinsed and incubated with a secondary antibody (anti IgG labeled with peroxidase, Amersham) and developed by chemiluminiscence system (ECL, Amersham) on X-ray film. Fig. 1. Effect of Ang II on PAI-1 mRNA by cultured HUVEC (n = 3) genotyped for the 4G/5G PAI-1 polymorphism. The time course of Ang IIinduced PAI-1 mRNA expression determined by real-time PCR in 4G/4G (panel A), 4G/5G (panel B) and 5G/5G (panel C) cultures, is shown. Values were normalized with h-actin and expressed as percentage of baseline. y p < 0.05 compared to basal; *p < 0.05 compared to unstimulated cultures. C. Roncal et al. / Cardiovascular Research 63 (2004) 176–185 179 3. Results 3.1. Influence of the 4G/5G PAI-1 genotype on Ang II induced PAI-1 expression by HUVEC PAI-1 genotyped cultured HUVEC were stimulated with 10 8 M Ang II, and the PAI-1 mRNA, as well as activity and protein were measured before and 2, 6, and 24 h after stimulation. The basal levels of PAI-1 mRNA (4G/4G: 36.6 F 0.6 AU, 4G/5G: 36.7 F 0.7 AU Fig. 2. Effect of Ang II on PAI-1 activity by cultured HUVEC (n = 3) genotyped for the 4G/5G PAI-1 promoter polymorphism. The time course of Ang II-induced PAI-1 amidolytic activity in 4G/4G (panel A), 4G/5G (panel B) and 5G/5G (panel C) endothelial cultures, is shown. Values are expressed as percentage of baseline (mean F S.E.). yp < 0.05 compared to basal; *p < 0.05 compared to unstimulated cultures. unstimulated cultures of the same genotype were assessed by Mann – Whitney U-test. Wilcoxon test was used to find differences of paired data. Pearson or Spearman tests were used for univariate associations of continuous variables. Multiple regression analysis was performed to determine whether PAI-1 genotype was an independent predictor of plasma PAI-1 antigen. The statistical analysis was performed with SPSS for Windows software package version 11.0. Fig. 3. Effect of Ang II on PAI-1 antigen secretion by cultured HUVEC (n = 3) genotyped for the 4G/5G PAI-1 polymorphism. The time course of Ang II-induced PAI-1 antigen in 4G/4G (panel A), 4G/5G (panel B) and 5G/5G (panel C) cultures, is shown. Values are expressed as percentage of baseline (mean F S.E.). yp < 0.05 compared to basal. 180 C. Roncal et al. / Cardiovascular Research 63 (2004) 176–185 Fig. 4. Confocal microscope images of immunofluorescence staining for PAI-1 protein (pseudocolored in green) in 4G/4G HUVEC treated for 24 h with Ang II (A), Ang II plus losartan (B) and unstimulated (C) cultures. A Z-section of each capture is shown at the top and the right hand side. No green fluorescence signal was detected in the absence of the primary antibody (D). Nuclei were counterstained with TOPRO-3 (red). and 5G/5G: 36.4 F 0.9 AU), PAI-1 activity (4G/4G: 30.3 F 4.6 U/ml, 4G/5G: 27.2 F 5.2 U/ml and 5G/5G: 2 1 . 4 F 3 . 8 U / m l ) a n d PA I - 1 a n t i g e n ( 4 G / 4 G : 1780.3 F 150.1 ng/ml, 4G/5G: 1553.5 F 167.0 ng/ml and 5G/5G: 1668.7 F 130.6 ng/ml), did not differ among genotypes. Fig. 5. PAI-1 antigen was measured in ECM by Western blot analysis in unstimulated HUVEC (lanes 3 – 5), after Ang II stimulation (lanes 6 – 8), and in cells pretreated with losartan 0.01 and 0.05 AM (lanes 9 – 14). Lane 1: molecular weight marker; lane 2: recombinant PAI-1. Panel B: Densitometric analysis of free PAI-1 corresponding to three independent experiments. Free PAI-1 plus PAI-1/vitronectin were completely blocked by losartan 0.05 AM after 24 h Ang II stimulation. Values are expressed as percentage of baseline (mean F S.E.). *p < 0.05 compared to unstimulated samples; zp < 0.05 compared to Ang IIstimulated samples. C. Roncal et al. / Cardiovascular Research 63 (2004) 176–185 181 As shown in Fig. 1, Ang II induced a significant increase of PAI-1 expression in 4G/4G HUVEC compared to either baseline or unstimulated cultures. PAI-mRNA was significantly elevated 2 h after stimulation (1.8-fold increase), the effect being maximal ( p < 0.05) at 6 h (2-fold increase), to return to basal values after 24 h. However, the addition of Ang II to 4G/5G and 5G/5G HUVEC was followed by a slight non-significant increase in PAI-1 mRNA. As shown in Fig. 2A, changes in 4G/4G mRNA paralleled those in PAI-1 activity (2.5-fold increase, p < 0.05) at 2 (67.8 F 6.2 U/ml) and 6 h (59.8 F 5.1 U/ ml) compared to baseline (24.3 F 3.3 U/ml), activity levels still remaining elevated (3-fold increase) at 24 h (72.8 F 5.7 U/ml). The addition of Ang II to 4G/5G HUVEC was followed by a lower increase in PAI-1 activity at 2 and 6 h ( p = 0.07), without differences in the activity for the 5G/ 5G cultures. Finally, no significant increase in the amount of PAI-1 released into medium was observed (Fig. 3) for all tested endothelial genotypes as compared with unstimulated cultures, further confirmed by Western blot analysis of supernatants (data not shown). The influence of genotype on the magnitude of response to Ang II was investigated by studying the ratio of Ang II stimulated/basal levels in the different HUVEC genotyped cultures. Mean PAI-1 mRNA and activity were respectively 1.5- and 1.9-fold higher in 4G/4G cultures compared to the remaining genotypes. Since no significant increase in the PAI-1 protein secreted into the medium could be detected after Ang II stimulation, further experiments were carried out to assess the amount of PAI-1 present in the cells and ECM. As shown in Fig. 4, laser confocal microscopy revealed a significant increase of PAI-1 protein in the basal surface of the cells after Ang II stimulation. Western blot analysis of ECM samples showed a significant increase of free PAI-1 (50 kDa) and PAI-1/vitronectin complexes (112 kDa) in 4G/4G HUVEC, 6 and 24 h after Ang II stimulation (Fig. 5). 3.2. Effect of losartan on Ang II-induced PAI-1 response by 4G/4G HUVEC cultures In a further step, the involvement of AT1 receptor on Ang II-induced PAI-1 expression in 4G/4G genotyped cultures was assessed by preincubation (18 h) with different losartan concentrations (0.01 – 0.05 AM). As shown in Fig. 6, the Ang II-induced PAI-1 mRNA was completely neutralized by the higher losartan dose (0.05 AM) after 6-h Ang II addition (220.9 F 10.7% vs. 116.2 F 20.7%, p < 0.05). This dose of losartan also reduced significantly the PAI-1 activity into the medium throughout the experiment. Moreover, a marked reduction in PAI-1 protein in the basal surface of cells (Fig. 4) and in the amount of free plus complexed PAI-1 within ECM was found in losartantreated HUVEC 6 h after Ang II stimulation, with complete inhibition after 24 h (Fig. 5). Fig. 6. Effect of losartan on Ang II-induced PAI-1 expression by 4G/4G genotyped cultures (n = 3). The Ang II-induced PAI-1 mRNA (A), PAI-1 activity (B) and antigen (C) in HUVEC pretreated with losartan (0.01 – 0.05 AM) is shown. Values are expressed as percentage compared to baseline (mean F S.E.). The high losartan dose (0.05 AM) completely blocked the Ang II-induced PAI-1 mRNA at 6 h and activity at 6 and 24 h. yp < 0.05 compared to basal; zp < 0.05 compared to Ang II. 3.3. 4G/5G PAI-1 polymorphism and protein levels in hypertensive subjects The possible interactions among the 4G/5G PAI-1 polymorphism, plasma PAI-1 antigen levels and arterial blood pressure was assessed in 122 apparently healthy subjects (mean age 50.5 F 0.8 years, 84% men) referred to our 182 C. Roncal et al. / Cardiovascular Research 63 (2004) 176–185 Table 1 Partial correlation between PAI-1 levels and the clinical and biochemical parameters analyzed after controlling for age and sex BMI (kg/m2) SBP (mm Hg) DBP (mm Hg) Total cholesterol (mg/dl) HDL-cholesterol (mg/dl) LDL-cholesterol (mg/dl) Triglycerides (mg/dl)a CRP (mg/l)a Glucose levels (mg/dl) Aldosterone (pg/ml) R P 0.25 0.07 0.10 0.11 0.18 0.13 0.20 0.19 0.31 0.25 0.01 0.38 0.25 0.20 0.04 0.15 0.02 0.02 0.01 0.01 BMI: body mass index; SBP: systolic blood pressure; DBP: diastolic blood pressure; CRP: C-reactive protein. a Logaritmically transformed variables to normalized their distribution. 28.6%), consistent with the Hardy –Weinberg equilibrium, and did not differ from other series [25]. The 4G/5G PAI-1 genotypes percentage and the presence of other cardiovascular risk factors were similar between groups (Table 2). Moreover, the 4G/4G hypertensive group had significantly higher plasma PAI-1 (51.0 F 10.1 ng/ml) than either 4G/4G normotensives (28.3 F 4.0 ng/ml) or hypertensives with the 5G allele ( p < 0.05). Multiple linear regression analysis showed that the PAI-1 genotype (4G/4G vs. 4G/5G plus 5G/5G) was a significant predictor of plasma PAI-1 antigen in hypertensives after adjustment for age, sex, glucose levels, BMI, CRP, dyslipidemia and smoking habits (r = 0.46, p < 0.01). 4. Discussion institution for global vascular risk assessment. In the overall population, hypertensive patients (n = 46) had significantly higher BMI (29.2 F 4.2 vs. 26.2 F 3.6, p < 0.01) and PAI-1 antigen (39.0 F 3.8 vs. 30.7 F 2.4 ng/ml, p < 0.05) than normotensive subjects (n = 76). Moreover, serum aldosterone levels were also significantly increased in hypertensives (176.9 F 78.1 vs. 150.0 F 59.0 pg/ml, p < 0.03). As shown in Table 1, there was a significant positive correlation between plasma PAI-1 levels, several biochemical parameters and markers of atherosclerosis. After controlling for age and sex, the associations remained for BMI ( p < 0.01), glucose ( p < 0.01), HDL-cholesterol ( p < 0.04), triglycerides ( p < 0.02), CRP ( p < 0.02) and serum aldosterone ( p < 0.01). PAI-1 genotype frequencies were: 4G/4G (n = 36, 29.5%), 4G/5G (n = 51, 41.8%) and 5G/5G (n = 35, This study demonstrates a genotype-specific regulation of PAI-1 in Ang II-stimulated HUVEC, the 4G/4G PAI-1 genotype being critical in endothelial PAI-1 response through the AT1 receptor, since the effect was completely prevented by losartan. In addition, higher PAI-1 antigen levels were found in hypertensive subjects homozygous for the 4G allele compared to the remaining genotypes. Taken together, our data strongly support a significant role for the 4G/5G PAI-1 genotype in determining PAI-1 response in arterial hypertension. The proatherogenic effects of Ang II appear to be partially related to reduced fibrinolysis [3,26]. PAI-1 is considered to be an important regulator of fibrinolysis and ECM turnover [7]. In addition, PAI-1 plays a crucial role in the development of atherosclerosis and neointimal forma- Table 2 Clinical characteristics of the studied population according to 4G/5G PAI-1 polymorphism (mean F S.E.M. is shown) 4G/4G Age, years Sex (M/F) BMI, kg/m2 Smokers (%) Diabetes (%) Systolic blood pressure, mm Hg Diastolic blood pressure, mm Hg Glucose, mg/dl Total cholesterol, mg/dl HDL cholesterol, mg/dl Triglycerides, mg/dl CRP, mg/l PAI-1 antigen, ng/ml 4G/5G 5G/5G Normotensives (n = 24) Hypertensives (n = 12) Normotensives (n = 27) Hypertensives (n = 24) Normotensives (n = 25) Hypertensives (n = 10) 47.7 F 2.9 22/2 26.2 F 0.9 50 5 117.5 F 2.5 53.3 F 1.7 11/1 29.3 F 1.1* 42 16 138.8 F 2.9** 48.6 F 1.8 22/5 26.8 F 0.6 48 4 115.6 F 1.8 52.7 F 1.7 20/4 28.7 F 0.9* 42 12 150.0 F 3.3** 48.0 F 1.9 19/6 24.7 F 0.6 40 2 113.0 F 2.2 54.0 F 2.9 7/3 29.1 F 1.1** 40 8 148.0 F 3.5** 76.1 F 1.5 88.7 F 2.1** 76.1 F 1.3 92.5 F 1.5** 76.5 F 1.3 91.5 F 3.6** 98.6 F 2.7 235.4 F 8.7 108.7 F 11.5 240.1 F 7.7 103.6 F 5.4 240.0 F 6.3 106.4 F 6.2 258.2 F 8.8 94.8 F 2.1 233.4 F 8.9 95.3 F 1.9 246.8 F 13.3 54.9 F 2.8 52.2 F 3.8 50.9 F 2.2 47.4 F 2.3 58.1 F 3.4 55.9 F 2.1 109.0 F 10.8 3.1 F 2.5 28.3 F 4.0 107.2 F 8.9 3.1 F 1.7 51.0 F 10.1* 118.4 F 15.4 2.4 F 1.6 30.9 F 4.3 141.8 F 16.4 4.4 F 4.1 29.6 F 2.9y 111.5 F 11.6 2.5 F 3.2 31.5 F 5.1 147.5 F 22.9 3.4 F 2.6 26.6 F 5.1y * p < 0.05 compared to normotensive. ** p < 0.01 compared to normotensive. y p < 0.05 compared to 4G/4G hypertensives. C. Roncal et al. / Cardiovascular Research 63 (2004) 176–185 tion after vascular injury [27,28]. Because of the association of PAI-1 gene 4G/5G polymorphism with circulating PAI-1 levels [14,15], we first tested the hypothesis that Ang IIinduced PAI-1 by HUVEC would be dependent on the endothelial 4G/5G genotype. Ang II significantly increased the PAI-1 gene expression in cultures with 4G/4G genotype compared to 5G allele. The exact mechanism by which Ang II might regulate endothelial PAI-1 expression in a genotype-specific manner has not been clearly defined. It can be speculated that the 4G/4G genotype, in the absence of repressor binding site, would elicit increased response to Ang II through nuclear transcription factor kB (NF-kB) pathway [13]. Ang II response sites have been identified upstream the PAI-1 promoter [29] involving signaling cascades depending on PKC activation, which in turn can activate NF-kB in endothelial cells [30]. The competition between the transcriptional factor and the 5G allele-specific transcriptional repressor protein might explain the increased PAI-1 response by HUVEC 4G/4G upon Ang II stimulation. Additional studies are, however, needed to examine how this factor might influence Ang II-induced PAI-1 at the promoter level. Previous studies have associated the homozygosity for the 4G allele with PAI-1 after RAAS activation by salt depletion [31], with triglycerides [14] and with circadian variations in PAI-1 [15]. The PAI-1 mRNA expression paralleled an increase in PAI-1 activity, but not in the amount of protein released into the medium suggesting post-translational modifications. Previous studies reporting elevated PAI-1 antigen after Ang II stimulation could be related to different endothelial origin [32] or experimental conditions. In fact, a slight but significant increase in Ang II-stimulated HUVEC was only observed with 100 times higher Ang II concentrations [33]. However, our data would agree with a previous study showing that calcium mobilizing agents (as Ang II) may indeed suppress the proinflammatory cytokines-induced increase in PAI-1 synthesis due to impaired protein translation [34]. Quite unexpectedly, we found that the majority of Ang II-induced PAI-1 antigen was localized within ECM. The relationship between Ang II and cardiovascular remodeling has not been clearly defined. Several studies have demonstrated that Ang II can modulate synthesis and degradation of ECM components [35,36]. Our results indicate that it may also favor the endothelial secretion of PAI-1 towards the subendothelial space, as demonstrated by the increased presence of free PAI-1 and PAI-1/vitronectin (Vn) complexes in ECM. PAI-1 and Vn are known to participate in the early stages of the vascular response to injury by stabilizing the initial thrombus and preventing early fibrinolysis [37] but are also involved in pathological fibrosis [38]. The presence of free and Vn-complexed PAI-1 within ECM in Ang II-stimulated HUVEC might reflect the limitation of excessive proteolysis favoring fibrous tissue deposition [39]. 183 We found that Ang II-induced PAI-1 by HUVEC in 4G/4G could be prevented by losartan. Whereas it has been previously reported that the 4G/5G genotype influenced the change in PAI-1 during ACE inhibition [40], no studies have demonstrated a similar effect of AT1 receptor blockers. Interestingly, a dose of losartan lower than required to reduce blood pressure in humans (0.05 AM) completely blocked Ang II-induced PAI-1 expression and this may be one of the mechanisms by which losartan may decrease the thrombotic potential of the vascular wall and increase fibrinolysis [41]. Controversy still exists, however, as to the effects of short- and long-term interruption of the RAAS system by either ACE inhibition or AT1 receptor antagonists on plasma fibrinolytic balance [41 –44]. The reduced amount of PAI-1 and PAI-1/Vn complexes within ECM in losartan-treated 4G/4G HUVEC would support a role for AT1 receptor blockade in the modulation of vascular fibrotic processes, as suggested by in vivo experiments [20]. In a next step, we investigated the relationship between PAI-1 polymorphism and PAI-1 antigen in relation to arterial hypertension, in subjects free of clinical cardiovascular disease. PAI-1 antigen was significantly increased in 4G/4G hypertensives compared to normotensives and hypertensives carrying the 5G allele. Results remained significant after adjusting for important confounding variables, such as BMI, triglycerides, glucose and CRP, thus emphasizing the pathophysiological relation between PAI-1 polymorphism, PAI-1 antigen and arterial hypertension. The observed correlation between the PAI-1 and aldosterone agrees with a previous report indicating that aldosterone is a major contributor of variability in PAI-1 levels in hypertensive subjects [26]. Impaired fibrinolysis due to elevated PAI-1 has been reported as a marker of early atherothrombosis in hypertensives [45]. Moreover, it has been recently pointed out a positive association between 4G/5G PAI-1 genotype and plasma PAI-1 activity in Chinese patients with arterial hypertension [46]. Some metabolic factors, such as insulin resistance and obesity, may also contribute to the high level of PAI-1 [47]. However, in the series analyzed, an independent association of PAI-1 and the 4G/5G polymorphism was observed in hypertensives, after adjustment for BMI and other metabolic factors. In conclusion, this study shows that Ang II-induced PAI-1 expression by HUVEC is dependent on the 4G/5G cell genotype and can be effectively prevented by losartan in 4G/4G cultures. The common 675 4G/5G polymorphism in the PAI-1 gene was associated with PAI-1 levels in hypertension, since hypertensive carriers of the 4G allele in its homozygous form had greater circulating PAI-1 antigen levels. Reduced fibrinolytic potential related to high PAI-1 might be an additional cardiovascular risk factor in 4G/4G homozygous with arterial hypertension. 184 C. Roncal et al. / Cardiovascular Research 63 (2004) 176–185 Acknowledgements This project was funded through the agreement between FIMA and the ‘‘UTE project CIMA’’. [18] [19] References [20] [1] Brasier AR, Recinos III A, Eledrisi MS. Vascular inflammation and the renin – angiotensin system. Arterioscler Thromb Vasc Biol 2002; 22:1257 – 66. [2] Kim S, Iwao H. Molecular and cellular mechanisms of angiotensin IImediated cardiovascular and renal diseases. Pharmacol Rev 2000;52: 11 – 34. [3] Vaughan DE. Angiotensin and vascular fibrinolytic balance. Am J Hypertens 2002;15:3S – 8S. [4] Ridker PM, Gaboury CL, Conlin PR, Seely EW, Williams GH, Vaughan DE. Stimulation of plasminogen activator inhibitor in vivo by infusion of angiotensin II. Evidence of a potential interaction between the renin – angiotensin system and fibrinolytic function. Circulation 1993;87:1969 – 73. [5] Vaughan DE, Lazos SA, Tong K. Angiotensin II regulates the expression of plasminogen activator inhibitor-1 in cultured endothelial cells. A potential link between the renin – angiotensin system and thrombosis. J Clin Invest 1995;95:995 – 1001. [6] Nishimura H, Tsuji H, Masuda H, et al. Angiotensin II increases plasminogen activator inhibitor-1 and tissue factor mRNA expression without changing that of tissue type plasminogen activator or tissue factor pathway inhibitor in cultured rat aortic endothelial cells. Thromb Haemost 1997;77:1189 – 95. [7] Podor TJ, Peterson CB, Lawrence DA, et al. Type 1 plasminogen activator inhibitor binds to fibrin via vitronectin. J Biol Chem 2000; 275:19788 – 94. [8] Paramo JA, Colucci M, Collen D, van de Werf F. Plasminogen activator inhibitor in the blood of patients with coronary artery disease. Br Med J 1985;291:573 – 4. [9] Hamsten A, de Faire U, Walldius G, et al. Plasminogen activator inhibitor in plasma: risk factor for recurrent myocardial infarction. Lancet 1987;2:3 – 9. [10] Eitzman DT, Westrick RJ, Xu Z, Tyson J, Ginsburg D. Plasminogen activator inhibitor-1 deficiency protects against atherosclerosis progression in the mouse carotid artery. Blood 2000;96:4212 – 5. [11] Orbe J, Chorda C, Montes R, Paramo JA. Changes in the fibrinolytic components of cultured human umbilical vein endothelial cells induced by endotoxin, tumor necrosis factor-alpha and interleukin-1alpha. Haematologica 1999;84:306 – 11. [12] Perez-Ruiz A, Montes R, Velasco F, et al. Regulation by nitric oxide of endotoxin-induced tissue factor and plasminogen activator inhibitor-1 in endothelial cells. Thromb Haemost 2002;88:1060 – 5. [13] Dawson SJ, Wiman B, Hamsten A, Green F, Humphries S, Henney AM. The two allele sequences of a common polymorphism in the promoter of the plasminogen activator inhibitor-1 (PAI-1) gene respond differently to interleukin-1 in HepG2 cells. J Biol Chem 1993;268:10739 – 45. [14] Eriksson P, Kallin B, van’t Hooft FM, Bavenholm P, Hamsten A. Allele-specific increase in basal transcription of the plasminogen-activator inhibitor 1 gene is associated with myocardial infarction. Proc Natl Acad Sci U S A 1995;92:1851 – 5. [15] van der Bom JG, Bots ML, Haverkate F, Kluft C, Grobbee DE. The 4G/5G polymorphism in the gene for PAI-1 and the circadian oscillation of plasma PAI-1. Blood 2003;101:1841 – 4. [16] Grenett HE, Benza RL, Fless GM, Li XN, Davis GC, Booyse FM. Genotype-specific transcriptional regulation of PAI-1 gene by insulin, hypertriglyceridemic VLDL, and Lp(a) in transfected, cultured human endothelial cells. Arterioscler Thromb Vasc Biol 1998;18:1803 – 9. [17] Nakamura S, Nakamura I, Ma L, Vaughan DE, Fogo AB. Plasmino- [21] [22] [23] [24] [25] [26] [27] [28] [29] [30] [31] [32] [33] [34] gen activator inhibitor-1 expression is regulated by the angiotensin type 1 receptor in vivo. Kidney Int 2000;58:251 – 9. Chen HC, Bouchie JL, Perez AS, et al. Role of the angiotensin AT(1) receptor in rat aortic and cardiac PAI-1 gene expression. Arterioscler Thromb Vasc Biol 2000;20:2297 – 302. Oubina MP, de Las Heras N, Vazquez-Perez S, et al. Valsartan improves fibrinolytic balance in atherosclerotic rabbits. J Hypertens 2002;20:303 – 10. Ma LJ, Nakamura S, Whitsitt JS, Marcantoni C, Davidson JM, Fogo AB. Regression of sclerosis in aging by an angiotensin inhibitioninduced decrease in PAI-1. Kidney Int 2000;58:2425 – 36. Grancha S, Estelles A, Tormo G, et al. Plasminogen activator inhibitor-1 (PAI-1) promoter 4G/5G genotype and increased PAI-1 circulating levels in postmenopausal women with coronary artery disease. Thromb Haemost 1999;81:516 – 21. Castello R, Estelles A, Vazquez C, et al. Quantitative real-time reverse transcription-PCR assay for urokinase plasminogen activator, plasminogen activator inhibitor type 1, and tissue metalloproteinase inhibitor type 1 gene expressions in primary breast cancer. Clin Chem 2002;48:1288 – 95. Paramo JA, Alfaro MJ, Rocha E. Postoperative changes in the plasmatic levels of tissue-type plasminogen activator and its fast-acting inhibitor-relationship to deep vein thrombosis and influence of prophylaxis. Thromb Haemost 1985;54:713 – 6. Chavakis T, Pixley RA, Isordia-Salas I, Colman RW, Preissner KT. A novel antithrombotic role for high molecular weight kininogen as inhibitor of plasminogen activator inhibitor-1 function. J Biol Chem 2002;277:32677 – 82. Festa A, D’Agostino Jr. R, Rich SS, Jenny NS, Tracy RP, Haffner SM. Promoter (4G/5G) plasminogen activator inhibitor-1 genotype and plasminogen activator inhibitor-1 levels in blacks, Hispanics, and non-Hispanic whites: the Insulin Resistance Atherosclerosis Study. Circulation 2003;107:2422 – 7. Srikumar N, Brown NJ, Hopkins PN, et al. PAI-1 in human hypertension: relation to hypertensive groups. Am J Hypertens 2002; 15:683 – 90. Carmeliet P, Moons L, Lijnen R, et al. Inhibitory role of plasminogen activator inhibitor-1 in arterial wound healing and neointima formation: a gene targeting and gene transfer study in mice. Circulation 1997;96:3180 – 91. Peng L, Bhatia N, Parker AC, Zhu Y, Fay WP. Endogenous vitronectin and plasminogen activator inhibitor-1 promote neointima formation in murine carotid arteries. Arterioscler Thromb Vasc Biol 2002; 22:934 – 9. Motojima M, Ando T, Yoshioka T. Sp1-like activity mediates angiotensin-II-induced plasminogen-activator inhibitor type-1 (PAI-1) gene expression in mesangial cells. Biochem J 2000;349:435 – 41. Kobayashi N, Nakano S, Mita S, et al. Involvement of Rho-kinase pathway for angiotensin II-induced plasminogen activator inhibitor-1 gene expression and cardiovascular remodeling in hypertensive rats. J Pharmacol Exp Ther 2002;301:459 – 66. Brown NJ, Murphey LJ, Srikumar N, Koschachuhanan N, Williams GH, Vaughan DE. Interactive effect of PAI-1 4G/5G genotype and salt intake on PAI-1 antigen. Arterioscler Thromb Vasc Biol 2001;21: 1071 – 7. Yoshizumi M, Tsuji H, Nishimura H, et al. Atrial natriuretic peptide inhibits the expression of tissue factor and plasminogen activator inhibitor 1 induced by angiotensin II in cultured rat aortic endothelial cells. Thromb Haemost 1998;79:631 – 4. Yoshida M, Naito Y, Urano T, Takada A, Takada Y. L-158,809 and (DAla(7))-angiotensin I/II (1 – 7) decrease PAI-1 release from human umbilical vein endothelial cells. Thromb Res 2002;105:531 – 6. Peiretti F, Alessi MC, Henry M, Anfosso F, Juhan-Vague I, Nalbone G. Intracellular calcium mobilization suppresses the TNF-alpha-stimulated synthesis of PAI-1 in human endothelial cells. Indications that calcium acts at a translational level. Arterioscler Thromb Vasc Biol 1997;17:1550 – 60. C. Roncal et al. / Cardiovascular Research 63 (2004) 176–185 [35] Masutomo K, Makino N, Fushiki MS. Effects of losartan on the collagen degradative enzymes in hypertrophic and congestive types of cardiomyopathic hamsters. Mol Cell Biochem 2001;224:19 – 27. [36] Papakonstantinou E, Roth M, Kokkas B, Papadopoulos C, Karakiulakis G. Losartan inhibits the angiotensin II-induced modifications on fibrinolysis and matrix deposition by primary human vascular smooth muscle cells. J Cardiovasc Pharmacol 2001;38:715 – 28. [37] Konstantinides S, Schafer K, Thinnes T, Loskutoff DJ. Plasminogen activator inhibitor-1 and its cofactor vitronectin stabilize arterial thrombi after vascular injury in mice. Circulation 2001;103:576 – 83. [38] Eitzman DT, Westrick RJ, Nabel EG, Ginsburg D. Plasminogen activator inhibitor-1 and vitronectin promote vascular thrombosis in mice. Blood 2000;95:577 – 80. [39] Sobel BE, Taatjes DJ, Schneider DJ. Intramural plasminogen activator inhibitor type-1 and coronary atherosclerosis. Arterioscler Thromb Vasc Biol 2003;23:1979 – 89. [40] Brown NJ, Abbas A, Byrne D, Schoenhard JA, Vaughan DE. Comparative effects of estrogen and angiotensin-converting enzyme inhibition on plasminogen activator inhibitor-1 in healthy postmenopausal women. Circulation 2002;105:304 – 9. [41] Brown NJ, Kumar S, Painter CA, Vaughan DE. ACE inhibition versus angiotensin type 1 receptor antagonism: differential effects on PAI-1 over time. Hypertension 2002;40:859 – 65. 185 [42] Vaughan DE, Rouleau JL, Ridker PM, Arnold JM, Menapace FJ, Pfeffer MA. Effects of ramipril on plasma fibrinolytic balance in patients with acute anterior myocardial infarction. HEART Study Investigators. Circulation 1997;96:442 – 7. [43] Brown NJ, Agirbasli M, Vaughan DE. Comparative effect of angiotensin-converting enzyme inhibition and angiotensin II type 1 receptor antagonism on plasma fibrinolytic balance in humans. Hypertension 1999;34:285 – 90. [44] Fogari R, Mugellini A, Zoppi A, et al. Losartan and perindopril effects on plasma plasminogen activator inhibitor-1 and fibrinogen in hypertensive type 2 diabetic patients. Am J Hypertens 2002; 15:316 – 20. [45] Diamantopoulos EJ, Andreadis EA, Vassilopoulos CV, et al. Increased plasma plasminogen activator inhibitor-1 levels: a possible marker of hypertensive target organ damage. Clin Exp Hypertens 2003;25:1 – 9. [46] Jeng JR. Association of PAI-1 gene promoter 4G/5G polymorphism with plasma PAI-1 activity in Chinese patients with and without hypertension. Am J Hypertens 2003;16:290 – 6. [47] Hoffstedt J, Andersson IL, Persson L, Isaksson B, Arner P. The common 675 4G/5G polymorphism in the plasminogen activator inhibitor-1 gene is strongly associated with obesity. Diabetologia 2002;45: 584 – 7.