Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
History of molecular evolution wikipedia , lookup
Genome evolution wikipedia , lookup
List of types of proteins wikipedia , lookup
Gel electrophoresis of nucleic acids wikipedia , lookup
Nucleic acid analogue wikipedia , lookup
Molecular cloning wikipedia , lookup
Cre-Lox recombination wikipedia , lookup
Endogenous retrovirus wikipedia , lookup
Deoxyribozyme wikipedia , lookup
Vectors in gene therapy wikipedia , lookup
Non-coding DNA wikipedia , lookup
Community fingerprinting wikipedia , lookup
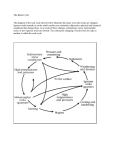
How We Know What Happened When Life began 3.8 billion years ago, and insects diversified 290 million years ago, but the human and chimpanzee lineages diverged only five million years ago. How have scientists figured out the dates of long past evolutionary events? Here are some of the methods and evidence that scientists use to put dates on events: 1. Radiometric dating relies on half-life decay of radioactive elements to allow scientists to date rocks and materials directly. 2. Stratigraphy provides a sequence of events from which relative dates can be extrapolated. 3. Molecular clocks allow scientists to use the amount of genetic divergence between organisms to extrapolate backwards to estimate dates. Radiometric Dating Geologists use radiometric dating to estimate how long ago rocks formed, and to infer the ages of fossils contained within those rocks. Radioactive elements decay. The universe is full of naturally occurring radioactive elements. Radioactive atoms are inherently unstable; over time, radioactive “parent atoms” decay into stable “daughter atoms.” When molten rock cools, forming what are called igneous rocks, radioactive atoms are trapped inside. Afterwards, they decay at a predictable rate. By measuring the quantity of unstable atoms left in a rock and comparing it to the quantity of stable daughter atoms in the rock, scientists can estimate the amount of time that has passed since that rock formed. Bracketing the fossils Fossils are generally found in sedimentary rock—not igneous rock. Sedimentary rocks can be dated using radioactive carbon, but because carbon decays relatively quickly, this only works for rocks younger than about 50 thousand years. So in order to date most older fossils, scientists look for layers of igneous rock or volcanic ash above and below the fossil. Scientists date igneous rock using elements that are slow to decay, such as uranium and potassium. By dating these surrounding layers, they can figure out the youngest and oldest that the fossil might be; this is known as “bracketing” the age of the sedimentary layer in which the fossils occur. Stratigraphy Fossils can be dated relative to one another by noting their positions in layers of rocks, known as strata. As shown in the animation (right), fossils found in lower strata were generally deposited earlier and are older. Sometimes geologic processes interrupt this straightforward, vertical pattern (left). For example, a mass of rock may cut across other strata, erosion may interrupt the regular pattern of deposition, or the rock layers may even be bent and turned upside-down. In the example at left, we can deduce that the oldest rocks are those that are cut through by other rocks. The next oldest rocks are those that are “doing the cutting” through the oldest rocks, and the youngest rocks lie on top of these layers and are not cut through at all. By making careful observations, we can detect these interruptions in the vertical pattern and use them to get more information about the relative ages of different layers. By studying and comparing strata from all over the world, we can date rocks relative to one another. Using numerical dating techniques, such as those based on the radioactive decay of atoms, we can assign probable ages to these layers and the fossils they contain. Venericardia planicosta, an Eocene bivalve during the Eocene. Certain fossils, referred to as index fossils, can be helpful as well. If an organism existed for a relatively short period of time and had a wide geographic distribution, then it can provide an index as to the age of the rocks in which it is preserved. For instance, Venericardia planicosta is known to have lived only during the Eocene, thus every time we find Venericardia planicosta, we can assume that the rocks containing this fossil must have been formed Molecular Clocks For the past 40 years, evolutionary biologists have been investigating the possibility that some evolutionary changes occur in a clock-like fashion. Over the course of millions of years, mutations may build up in any given stretch of DNA at a reliable rate. For example, the gene that codes for the protein alpha-globin (a component of hemoglobin) experiences base changes at a rate of .56 changes per base pair per billion years*. If this rate is reliable, the gene could be used as a molecular clock. When a stretch of DNA does indeed behave like a molecular clock, it becomes a powerful tool for estimating the dates of lineage-splitting events. For example, imagine that a length of DNA found in two species differs by four bases (as shown below) and we know that this entire length of DNA changes at a rate of approximately one base per 25 million years. That means that the two DNA versions differ by 100 million years of evolution and that their common ancestor lived 50 million years ago. Since each lineage experienced its own evolution, the two species must have descended from a common ancestor that lived at least 50 million years ago. This general technique has been used to investigate several important issues, including the origin of modern humans, the date of the human/chimpanzee divergence, and the date of the Cambrian “explosion.” Using molecular clocks to estimate divergence dates depends on other methods of dating. In order to calculate the rate at which a stretch of DNA changes, biologists must use dates estimated from other relative and absolute dating techniques. *This number is for changes that affect the structure of the protein Lesson Plan about Molecular Clocks THEORY Introduction Every human cell has a "second" genome, found in the cell's energy-generating organelle, the mitochondrion. In fact, each mitochondrion has several copies of its own genome, and there are several hundred to several thousand mitochondria per cell. This means that the mitochondrial (mt) genome is highly amplified. While each cell contains only two copies of a given nuclear gene (one on each of the paired chromosomes), there are thousands of copies of a given mt gene per cell. Because of this high copy number, it is possible to obtain a mt DNA type from the equivalent of a single cell's worth of mt DNA. Thus, mt DNA is the genetic system of choice in cases where tissue samples are very old, very small, or badly degraded by heat and humidity. Under good circumstances - working from fresh cell samples - mt DNA is the easiest human DNA to amplify by PCR. This experiment examines a 440-nucleotide sequence from the noncoding region of mt genome. Hand cycling is a realistic alternative to automated thermal cyclers, and the high yield of amplified product can be visualized in an agarose gel with a variety of stains. Because each student is amplifying the same region, the gel electrophoresis results will also be the same for each. However, amplified student samples may be submitted to our Sequencing Service, which will generate student mt DNA sequences and post the results on our Sequence Server. Comparison of control region sequences reveals that most people have a unique pattern of single nucleotide polymorphisms (SNPs). These sequence differences, in turn, are the basis for farranging investigations on human DNA diversity and the evolution of hominids.