Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Drosophila melanogaster wikipedia , lookup
Immune system wikipedia , lookup
Lymphopoiesis wikipedia , lookup
Polyclonal B cell response wikipedia , lookup
Adaptive immune system wikipedia , lookup
Cancer immunotherapy wikipedia , lookup
Psychoneuroimmunology wikipedia , lookup
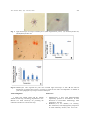
271 Advances in Environmental Biology, 4(2): 271-276, 2010 ISSN 1995-0756 © 2010, American-Eurasian Network for Scientific Information This is a refereed journal and all articles are professionally screened and reviewed ORIGINAL ARTICLE Effect of temperature on phagocytosis activity in garden snails Helix aspersa Ahmad. K. Ma'aqbeh Department of Biology Al-Hussein bin Talal University Ma'an, Jordan Ahmad. K. Ma'aqbeh, Effect of temperature on phagocytosis activity in garden snails Helix aspersa; Adv. Environ. Biol., C(): CC-CC, 2010 ABSTRACT Helix aspersa, known as garden snail, is a species of land snail, a pulmonate gastropod. This snail is very common and widespread in Mediterranean region and western Europe. This species is characterized by a spherical in shape shell with a short spire and a rough surface. The shell color is pale brown, or yellow with a number of broken dark bands, which give the shell it, is blotchy appearance. This study investigates the effect of temperature on phagocytosis activity in garden snails H. aspersa. Low and high temperatures depressed phagocytosis activity and decrease the number of yeast cells phagocyted by H.aspersa hemocytes. The phagocytosis activity at 25Co is (50.8±4.5), 35 Co (19.2 ± 3.9), 40 Co (3.6±0.89), and 15 Co (4.4±1.1).Two types of hemocytes cell GLC (Granulocyte like Cell), and HLC (Hyalinocyte like Cell) are diagnosed Key words: phagocytosis, HLC, GLC, H. aspersa, stress, yeast Introduction Invertebrates are widely distributed and can be found in almost any kind of habitat. Their dispersal and survival depend on successful defenses mechanism against various kinds of microorganisms and parasites, especially that they live in the same environmental conditions [1]. Invertebrates lack immunoglobulin antibodies and their associated adaptive immune responses; instead, they have highly effective innate immune reactions that defend them against a broad spectrum of microorganisms [3]. However, previous study has shown that a specific memory that consider as acquired immunity in defense systems of invertebrates [4]. Invertebrate innate immune system is composed of both cellular and humoral immune responses. hemocytes-mediated immune responses such as phagocytosis, nodule formation and melanotic encapsulation are major cellular responses [5, 6] Hemocytes from bivalve mollusks have most often been divided into two types, hyalinocytes and granulocytes, both of which are phagocytic [7]. These two cell types have been found in many bivalve species [7, 8]. In many of these species, granulocytes have been further subclassified as eosinophilic, basophilic and neutrophilic cells [9]. Granulocytes have the ability to phagocytose microbial pathogens and they contain a mixture of hydrolytic enzymes that contribute to intracellular killing. They are often more active phagocytes than hyalinocytes. Hyalinocytes are smaller than granulocytes. Some contain few granules and are usually more morphologically heterogeneous than granulocytes. Hyalinocytes have been divided into two types, small hyalinocytes, with large nuclei and scanty cytoplasm, and large hyalinocytes with low nucleus: cytoplasm ratios [10]. Phagocytic blood cells are major component of innate defense in both invertebrates and vertebrates, where they are also critical for linking innate to adaptive immunity. Moreover, phagocytosis seems relevant for one of the most promising candidates for specific immunity in insects and crustaceans,the alternatively spliced Down syndrome cell adhesion Corresponding Author Ahmad. K. Ma'aqbeh, Department of Biology Al-Hussein bin Talal University Ma'an, Jordan Tel. +962785656776, E-mail: [email protected] [email protected] Adv. Environ. Biol., 4(2): 271-276, 2010 molecule (Dscam).previous study demonstrate that, enhanced phagocytosis resulting from prior exposure to endotoxin or live pathogenic bacteria in invertebrates [11]. Both the density and functions of mollusc hemocytes can be affected by changes in environmental conditions. Different types of stressors, including pathogens, parasites and xenobiotic, have been identified as hemocytes effectors. Hemocytes response patterns may depend on the nature of stressor [12, 13]. Hemocytes also serve a role in the molluscan stress response involving endocrine molecules shared with the vertebrates [14, 15]. Recent studies have shown that molluscs possess a primitive form of neuroendocrine response to stress involving catecholamine and neuropeptides such as adrenocorticotropic hormone [16]. The objective of this study to investigated the effect of different temperature on phagocytosis activity in Helix aspersa 2. Materials and method 2.1. Snails: Adult specimens of Helix aspersa were obtained from the northern area of AL-Karak city.it is governorate lies 140 km to the south of Amman on the King's Highway. It is situated on a hilltop about 1000 meters above sea level and is surrounded on three sides by a valley and from Karak can be seen wonder view of the Dead Sea, and the geographical coordinates to karak Governorate are 31° 11' 5" North, 35° 42' 17" East. The snails were kept in plastic container, at room temperature. The water and soil was changed every week and the snails were fed lettuce twice weekly. Treated snails were kept and incubated in plastic container at different temperature (25Co, 35 Co, 40 Co, and 15 Co) for two days. 2.2. Hemolymph sampling and counts Hemolymph was collected directly from the heart using 22-gauge needles fitted to 3-ml syringes When the snail retracts into its shell, a drop of hemolymph was extruded through the hemal pore, and can be collected using a micropipette. Hemolymph (10 µl) was directly dropped in hemocytometer. Total number of living hemocytes and the number of hemocytes in each cell population identified according to, Van der Knaap [17] using Optika B350 (Italy) light microscopy. 2.3. Cells analysis and morphology 2.3.1. Live cell analysis 272 The analysis of live hemocytes was carried out by placing a 30 µl of whole hemolymph on acid alcohol cleaned microscope slides and allowing the cells to adhere for 10 min. The slides were then covered with a glass cover slip and viewed at high magnification with an Inverted phase microscope (Leica, Germany) 2.3.2. Staining method After adhesion to slides, hemocytes were fixed for 20 min in methanol (80%) and then immersed in Giemsa stain for 30 min before being washed in distilled water and air dried. Slides were observed by Optika B350 (Italy) light microscopy. 2.4. Phagocytic activity Phagocytic activity was assessed using baker’s yeast, Saccharomyces cerevisiae (Sigma Aldrich), as target cells. Five milligrams of yeast was suspended in 5ml distilled water and mixed with an equal volume of Congo red. The suspension was autoclaved at 120 Co for 15 min before being washed twice by centrifugation at 1300g for 5 min and resuspended in 10 ml distilled water .To measure phagocytosis; 100 ml of whole hemolymph (adjusted to 1×106 hemocytes ml-1) was placed on glass coverslips (22×22 mm) and incubated in a moist chamber. Hemocytes were allowed to settle and adhere for 20 min at room temperature (25 Co). The supernatants were then removed and the adherent hemocytes were rinsed 5 times with distilled water. The coverslips were overlaid with 100 ml of Congo red stained yeast (0.7×106 ml-1) and incubated for 30 min at room temperature. The coverslips were washed 10 times with distilled water to remove nonphagocytosed yeast cells. They were then inverted over clean microscope slides and sealed with nail polish. A minimum of 100 hemocytes on each coverslip were examined and the number of hemocytes that had phagocytosed one or more yeast was recorded so that the percentage of phagocytic cells could be calculated [10]. 2.5. Statistical analysis All data were presented as means±standard errors. All experiments were conducted three times. Data are shown for one representative experiment from each trial. In each experiment, different groups of five snails were tested at each time point. Data were analyzed using the Microsoft Excel package. Statistical analysis was performed by Student’s t-test to determine the significance of differences between mean values. Differences were considered to be significant if P<0.05[10] Adv. Environ. Biol., 4(2): 271-276, 2010 3. Results 3.1. Hemocyte morphology Two major hemocytes types were identified by light microscopy according to presence or absence of granules in their cytoplasm, hyalinocytes-like cells (HLC) and granulocytes-like cells (GLC). HCL were the first type of hemocytes identify in H. aspersa (Fig. 1B) these cells comprised 57.6 ±2.6% of the total hemocytes population. They varied in size within a range of 6–10µm, with an average diameter of 9 ± 0.9 µm. HLC had an amoeboid movement and they formed filopodia. HLC were also characterized by a cytoplasm containing no granules. Some HLC maintained a round shape when attached to glass slides and spread only slightly, forming some small filopodia. GLC were the second type of hemocytes identified in H. aspersa (Fig.1A). These cells comprised 42.4 ± 2.6% of the total hemocytes population. They varied in size from 5–12 µm with an average diameter of 9.6 ± 1.4 µm. GLC adhered to glass slides within 15–20 min and they were highly ameboid after they adhered. They also had the ability to form many long filopodia, and were characterized by numerous cytoplasmic granules. 3.2. Phagocytosis activity H.aspersa hemocytes were capable of ingesting yeast cells (Fig.2A). The phagocytic activity was measured at different temperature (Fig.2B), and a significant decreased in activity of H.aspersa held at 35 Co (19.2 ± 1.95) as compared to those treated at 25 Co as control 50.8±2.3,( P < 0.05),and the activity represent more decreasing at 40 Co (3.6±0.4) (P < 0.05) than the activity at 35 Co as compared to the activity at 25Co . When the H.aspersa kept in low temperature (15Co), the phagocytic activity dropped to (4.4±0.6) with compared to those at 25Co. very little difference in phagocytic activity was found between H.aspersa kept at 40 Co and those kept at 15 Co. The average number of yeast cells which was detected in haemocyte of H.aspersa kept at 25Co is (4cells±0.4), 35Co (2cells±0.4), 40 Co (1.2cells±0.2), and 15 Co (1.2cells±0.2) (Fig.2 C). 4. Discussion Hemocyte classification in invertebrates is often a controversial issue. Simply because there is no universal procedure to classify hemocytes [10]. In bivalves mollusks, hemocytes have been classified in many species, such as R. philippinarum, M. lusoria, C.gigas, C. farreri, T. crocea, D. polymorpha, and M. galloprovincialis [18, 19]. Hemocytes classification depends mainly on host defense, including the types of granules and enzymes found 273 within cells [10]. Blood cells are differentiated into two types: granulocytes and hyalinocytes, [7, 8]. The results of the present study confirm that there are two types of hemocytes (type I, type II), this result are similar to that reported in helix aspersa maxima into(type I, type II). [20] Similar finding has been reported earlier in P. corneus [21, 22]. Haemocytes type I in H. aspersa morphologically similar to granulocytes, and haemocytes II to hyalinocytes therefor it is possible to suggest that hemocyte type I is Granulocyte like Cell (GLC), and hemocyte type II is Hyalinocyte like Cell (HLC). Several studies have demonstrated that defence mechanisms in mollusks are susceptible to changes in environmental factors, such as temperature, salinity, dissolved oxygen, food availability and presence of pollutants [23, 24, 31, 32, 33]. In particular, increases in temperature have been shown to induce mortality in bivalves, probably due to fact that high temperature induced suppression to the immune system in these animals [25, 34]. Furthermore, the data obtained of the present study demonstrated that H.aspersa hemocytes are sensitive to temperature and there is a good correlation between temperature and phagocytic activity in this species. Phagocytosis is a temperature-dependent process in bivalve molluscs, so it is the main mechanism against foreign materials in snail's internal defence system [26]. In the present study, a significant decrease of phagocytic activity was found in snails kept at (15 Co, 35Co, 40 Co) with respect to those kept at 25 Co. Similar finding has been reported in C. virginica when exposed to a temperature variety for 1 week [27]. Conversely, Carballal MJ [28] found that the percentage of phagocytic haemocytes from M. galloprovincialis was lower at 10 Co than at 20 Co and 30 Co. [29] observed no differences between phagocytosis at 15 Co and 21 Co in both, R. decussatus, and M. galloprovincialis. In the present study, the efficiency of hemocyte for phagocyted yeast cell decreased at 15 Co to (1.2 cells), 35 Co to (2 cells), and 40 Co to (1.2 cells) respectively when compared with 25 Co (4 cells), a previous study [30] indicate that the max number zymosan particles was phagocyted by hemocytes cells is (3 particles) at room temperature. In conclusion, the influence of temperature on activity of phagocytosis has been investigated, and showed that both high temperature and low temperature have a clear effect on phagocytosis. As the temperature increased, the activity is clearly reduced. A similar pattern has been recorded with low temperature. Two types of cells (GLC, HLC) seem to have role in phagocytosis in the present investigation. Adv. Environ. Biol., 4(2): 271-276, 2010 274 Fig. 1. Light microscope of H.aspersa hemocytes at (40X magnification power). (A) Granulocyte-like cell, (B) Hyalinocyte-like cell. Fig 2A: Hematocytes cells ingested the yeast cells scanned light microscope at 40X. B: the effect of temperature in phagocytosis activity in H.aspersa snails C: the effect of temperature in number of yeast cells phagocytic by H.aspersa snail's hemocytes Acknowledgments References I thank prof. Khalil Altaif and Dr. Saleem Adaileh for her help, and biology department in ALHussein bin Talal University for providing the chemicals and devices and for her help. 1. 2. Mendoza H.L., I. Faye, 1996 Immunoglobulin superfamily proteins in invertebrates. In New Directions in Invertebrate Immunology. SOS publication, 285-302. Hoffmann, J.A., F.C. Kafatos, C.A. Janeway, R.A. Ezekowitz, 1999. Phylogenetic perspectives in innate immunity. Science, 284: 1313-1318. Adv. Environ. Biol., 4(2): 271-276, 2010 3. 4. 5. 6. 7. 8. 9. 10. 11. 12. 13. 14. 15. Perdomo Morales, R., V. Montero Alejo, E. Perera, 2007. Phenoloxidase activity in the hemolymph of the spiny lobster Panulirus argus. Fish Shellfish Immunol., 23: 1187-1195. Kurtz, J., 2005. Specific memory within innate immune systems. Trends Immunol., 26: 186-192. Lemaitre, B., J. Hoffmann, 2007. The host defense of Drosophila melanogaster. Annu Rev Immunol., 25: 697-743. Williams, M.J., 2007. Drosophila hemopoiesis and cellular immunity. J Immunol., 178: 4711-6. Wootton, E.C., R.K. Pipe, 2003. Structural and functional characterisation of the blood cells of the bivalve mollusc, Scrobicularia plana. Fish Shellfish Immunol., 15: 249–262. Chang, S.J., S.M. Tseng, H.Y. Chou, 2005. Morphological characterization via light and electron microscopy of the hemocytes of two cultured bivalves: A comparison study between the hard clam (Meretrix lusoria) and Pacific oyster (Crassostrea gigas). Zool. Stud., 44: 144–152. Lopez, C., M.J. Carballal, C. Azevedo, A. Villalba, 1997c. Morphological characterization of the Hemocytes of the Clam, Ruditapes decussatus (Mollusca: Bivalvia). J. Invertebr. Pathol., 69: 51–57. Aladaileh, S., S.V. Nair, D. Birch, D. Raftos, 2007. Sydney rock oyster (Saccostrea glomerata) hemocytes: morphology and function. J. Invert. Pathol., 96: 48-53. Roth, O., J. Kurtz, 2009. Phagocytosis mediates specificity in the immune defence of an invertebrate, the woodlouse Porcellio scaber (Crustacea: Isopoda). Developmental and Comparative Immunology, 33: 1151–1155. Galloway, T.S., M.H. Depledge, 2001. Immunotoxicity in invertebrates: Measurement and ecotoxicological relevance. Ecotoxicology, 10: 5–23. Livingstone, D.R., J.K. Chipman, D.M. Lowe, C. Minier, C.L. Mitchelmore, M.N. Moore, L.D. Peters, R.K. Pipe, 2000. Development of biomarkers to detect the effects of organic pollution on aquatic invertebrates: Recent molecular, genotoxic, cellular and immunological studies on the common mussel (Mytilus edulis L.) and other mytilids. Int. J. Environ. Pollut., 13: 56–91. Franchini, A., D. Kletsas and E. Ottaviani, 1996. Immunocytochemical evidence of PGF- and TGF-b-like molecules in invertebrate and vertebrate immunocytes: an evolutionary approach. Histochemical Journal, 28: 599–605. Ottaviani, E., A. Franchini and P. Fontanili, 1992. The presence of immunoreactive vertebrate bioactive peptide substances in hemocytes of the freshwater snail Viviparus ater (Gastropoda, Prosobranchia). Cellular and Molecular Neurobiology, 12: 455–462. 275 16. Lacoste, A., S.K. Malham, A. Cueff, S.A. Poulet, 2001d. Stress-induced catecholamine changes in the hemolymph of the oyster Crassostrea gigas. Gen. Comp. Endocrinol., 122: 181-188. 17. Van der Knaap, W.P.W., C.M. Adema, T. Sminia, 1993. Invertebrate blood cells: morphological and functional aspects of the haemocytes in the pond snail Lymnaea stagnalis. Comp. Haematol. Int., 3: 20-26. 18. Allam, B., K.A. Ashton-Alcox, S.E. Ford, 2002b. Flow cytometric measurement of hemocyte viability and phagocytic activity in the clam, Ruditapes philippinarum. J. Shellfish Res., 21: 13–19. 19. Jing, X., Z. Wenbin, 2005. Characterisation of monoclonal antibodies to haemocyte types of scallop (Chlamys farreri). Fish Shellfish Immunol., 19: 17–25. 20. Adamowicz, A., M. Bolaczek, 2003. blood cells morphology of the snail helix aspersa maxima (Helicidae). J. 9Zoologica Poloniae.48/1-4: 93101((26)). 21. FranceschI, C., A. Cossarizza, D. Monti, E. Ottaviani, 1991. Cytotoxicity and immunocyte markers in cells from the freshwater snail Planorbarius corneus (L.) (Gastropoda Pulmonata): implications for the evolution of natural killer cells. Eur. J. Immunol., 21: 489493. 22. Ottaviani, E., 1989. Selected decrease of haemocytes of the freshwater snail Planorbarius corneus (L.) (Gastropoda, Pulmonata) after bacteria injection. Experientia, 45: 368-369. 23. Gagnaire, B., H. Frouin, K. Moreau, H. ThomasGuyon, T. Renault, 2006. Effects of temperature and salinity on haemocyte activities of the Pacific oyster, Crassostrea gigas (Thunberg). Fish and Shellfish Immunology, 20: 536e47. 24. Matozzo, V., M. Monari, J. Foschi, T. Papi, O. Cattani, M.G. Marin, 2005. Exposure to anoxia of the clam Chamelea gallina. I: effects on immune responses. Journal of Experimental Marine Biology and Ecology, 325: 163e74. 25. Shumway, S.E., 1996. Natural environmental factors. In: Kennedy VS, Newell RIE, Eble AF, editors. The Eastern oyster Crassostrea virginica. College Park (MD) USA: Maryland Sea Grant., pp: 467-513. 26. Oliver, L.M., W.S. Fisher, 1999. Appraisal of perspective bivalve immunomarkers. Biomarkers, 4: 510-530. 27. He´garet, H., G.H. Wikfors, P. Soudant, 2003. Flow-cytometric analysis of hemocytes from eastern oysters, Crassostrea virginica, subjected to a sudden temperature elevation: II. Hemocyte functions: aggregation, viability, phagocytosis and respiratory burst. Journal of Experimental Marine Biology and Ecology, 293: 249-265. Adv. Environ. Biol., 4(2): 271-276, 2010 28. Carballal, M.J., C. Lo´pez, C. Azevedo, A. Villalba, 1997. In vitro study of phagocytic ability of Mytilus galloprovincialis Lmk. haemocytes. Fish and Shellfish Immunology, 7: 403-416. 29. Ordas, M.C., B. Novoa, A. Figueras, 1999. Phagocytosis inhibition of clam and mussel haemocytes by Perkinsus atlanticus secretion products. Fish and Shellfish Immunology, 9: 491-503. 30. Coen M. Adema, Elly C. van Deutekom-Mulder, Wil P.W. van der Knaap, Elisabeth A. Meuleman and Taede Sminia, 1991. GENERATION OF OXYGEN Radicals in hemocytes of the snail lymnaea stagnalis in relation to the rate of phagocytosis. Developmental and Comparative Immunology, 15: 17- 26. 31. Camus, L., B.E. Grøsvikb, J.F. Børseth, M.B. Jones, M.H. Depledge, 2000. Stability of lysosomal and cell membranes in haemocytes of the common mussel (Mytilus edulis): effect of low temperatures. Marine Environmental Research, 50: 325-329. 276 32. Matozzo, V., M.G. Marin, 2005. 4-Nonylphenol induces immunomodulation and apoptotic events in the clam Tapes philippinarum. Marine Ecology Progress Series, 285: 97-106. 33. Monari, M., V. Matozzo, J. Foschi, M.G. Marin, O. Cattani, 2005. Exposure to anoxia of the clam, Chamelea gallina. II: modulation of superoxide dismutase activity and expression in haemocytes. Journal of Experimental Marine Biology and Ecology, 325: 175-188. 34. Ford, S.E., M.R. Tripp, 1996. Diseases and defense mechanisms. In: Kennedy VS, Newell RIE, Eble AF, editors. The Eastern oyster Crassostrea virginica. College Park (MD) USA: Maryland Sea Grant., pp: 581-660.