Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
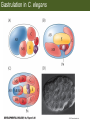
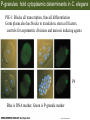
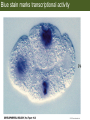
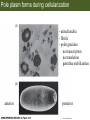
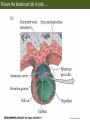
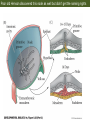
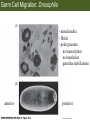
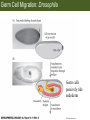
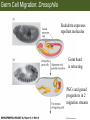
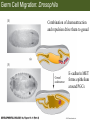
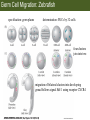
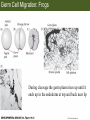
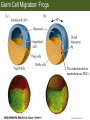
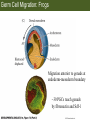
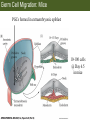
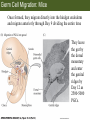
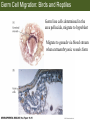
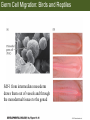
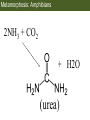
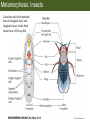
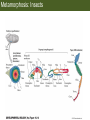
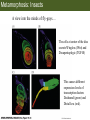
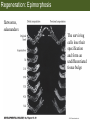
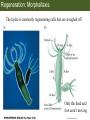
Bio 127 - Section III Late Development Germ Line Development Gilbert 9e – Chapter 16 Section 4 Encompasses : • • • • Development of the Tetrapod Limb Sex Determination The Saga of the Germ Line Post-Embryonic Development Student Learning Objectives 1. You should understand that sexual reproduction requiring the fusion of gametes from male and female gonads occurs in specific organisms. 2. You should understand that the primordial germ cells that give rise to gametes arise outside of the gonads and must migrate to them. 3. You should understand that in most organisms the primordial germ cells are specified conditionally, while in some they are specified autonomously by cytoplasmic determinants in the egg. 4. You should understand that migration of the germ cells from their site of origin to the gonads is an essential part of reproductive success . • In all plants and some animals, somatic cells can readily form new organisms – Cnidarians, flatworms, tunicates • In many animals, there is an early division between somatic and germ cells – Insects, roundworms, vertebrates • Two step process; – Primordial germ cells (PGCs) are determined in a specific location in the embryo – PGCs migrate to the gonad and become the progenitor population for eggs and sperm Two Methods of Germ Cell Determination • Autonomous Specification – Egg cytoplasmic determinants – Called ‘Germ Plasm’ – Nematodes, flies, frogs • Conditional Specification – Signals from surrounding cells – Majority of sexually reproducing organisms – Including mammals The nematode Caenorhabditis elegans Remember cleavage and gastrulation: Asymmetrical divisions produce a stem cell (P-lineage), “founder” cell. Stem cell divisions are meridional Founder cell divisions are equatorial Gastrulation in C. elegans P-granules hold cytoplasmic determinants in C. elegans PIE-1: Blocks all transcription, thus all differentiation Germ plasm also has blocks to translation, stem cell factors, controls for asymmetric divisions and meiosis inducing agents. P4 Blue is DNA marker, Green is P-granule marker Blue stain marks transcriptional activity P4 Synctitial cleavage in Drosophila is followed by cellularization Pole plasm forms during cellularization - mitochondria - fibrils - polar granules no transcription no translation germline stabilization anterior posterior Localization of germ cell-less (gcl) gene products Human males with mutant homolog are often sterile Germ plasm at the vegetal pole of frog embryos Marker for frog homolog of fly/worm translation blocker, Nanos • The frog cells that take up these granules will become PGCs and migrate to the gonads as the kidney forms – Again, no transcription or translation – Therefore, no differentiation Conditional Specification of mammalian PGCs • Posterior of epiblast at the junction of the primitive streak and extraembryonic ectoderm – Cells are no different from other epiblast – Specified in gastrulation before 3 layers form – Wnts from endoderm make them competent – BMPs from extraembryo ectoderm finish it Picture the blastocyst full of yolk..... Poor old Henson discovered this node as well but didn’t get the naming rights Conditional Specification of mammalian PGCs • Same deal as the others: – Repress differentiation by repressing gene expression • Specified outside embryo forming cells – Once expression is shut down they can go back into embryo and not respond to signals Germ Cell Migration • • • • • Drosophila Zebrafish Frogs Mice Birds and Reptiles Germ Cell Migration: Drosophila As the endoderm invaginates, the ectoderm and mesoderm extend and converge to wrap around the dorsal side to form the “germ band” Germ Cell Migration: Drosophila - mitochondria - fibrils - polar granules no transcription no translation germline stabilization anterior posterior Germ Cell Migration: Drosophila Germ cells passively ride endoderm Germ Cell Migration: Drosophila Endoderm expresses repellent molecules Germ band is retracting PGCs and gonad progenitors in 2 migration streams Germ Cell Migration: Drosophila Combination of chemoattraction and repulsion drive them to gonad E-cadherin MET forms epithelium around PGCs Germ Cell Migration: Drosophila • Both mesoderm and PGCs divide through the larval stage, differentiate at metamorphosis • At larval-pupal transition anterior PGCs in gonad become germ line stem cells • In ovaries, the cells attach top stromal cap In testes, the cells attach to hub cells Remember: Zebrafish development occurs very rapidly 24-hours from 1 cell to vertebrate embryo! Germ Cell Migration: Zebrafish specification: germ plasm determination: PGCs by 32-cells four clusters join into two migration of bilateral clusters into developing gonad follows signal Sdf-1 using receptor CXCR4 Remember: Germ plasm at vegetal pole in frogs Marker for frog homolog of fly/worm translation blocker, Nanos Germ Cell Migration: Frogs During cleavage the germ plasm rises up until it ends up in the endoderm at top and back near lip Germ Cell Migration: Frogs The endoderm below mesdoderm are PGCs Germ Cell Migration: Frogs Migration anterior to gonads at endoderm-mesoderm boundary ~30 PGCs reach gonads by fibronectin and Sdf-1 • Remember Sdf-1 • Soluble signal whose receptor is CXCR4 • Common signal for vertebrate germ cells • Also used by humans to call HSC to bone marrow, guide lymphocytes, MSC? Germ Cell Migration: Mice PGCs formed in extraembryonic epiblast 10-100 cells @ Day 6.5 in mice Germ Cell Migration: Mice Once formed, they migrate directly into the hindgut endoderm and migrate anteriorly through Day 9 dividing the entire time They leave the gut by the dorsal mesentary and enter the genital ridges by Day 12 as 2500-5000 PGCs. Germ Cell Migration: Mice • The travelling stem cell niche – Support cells travel with PGCs to maintain the undifferentiated stem cell phenotype – They secrete stem cell factor (SCF) – The cells follow fibronectin trail – Sdf-1 also required Germ Cell Migration: Birds and Reptiles Germ line cells determined in the area pellucida, migrate to hypoblast Migrate to gonads via blood stream when extraembryonic vessels form Germ Cell Migration: Birds and Reptiles Sdf-1 from intermediate mesoderm draws them out of vessels and through the mesodermal tissues to the gonad Bio 127 - Section III Late Development Post-Embryonic Development Gilbert 9e – Chapter 15 Section 4 Encompasses : • • • • Development of the Tetrapod Limb Sex Determination The Saga of the Germ Line Post-Embryonic Development Student Learning Objectives 1. You should understand that development never stops during the life of the organism and that three major processes occur in the postembryonic animal: metamorphosis, regeneration and aging. 2. You should understand the Direct Development involves young organisms with the same body plan as the adult; whereas Indirect Development involves major changes to form the adult body plan. 3. Indirect Development, or metamorphosis, is hormonal reactivation of 4. You should understand that regeneration is the reactivation of developmental process to restore missing tissues. 5. You should understand that aging and physiological senescence are an interplay of genetic and environmental influences. Metamorphosis • Development of a larval stage and an adult stage specialized for different functions – Larvae often specialized for growth, dispersal, etc. – Adults usually specialized for reproduction – Example Cecropia moths: • Larvae are wingless eating machines • Adults have one day to mate – don’t even have mouth parts! • Two major types of larvae – Primary: little to no similarity to adult (sea urchins) – Secondary: add and subtract parts from similar form • (insects, amphibians) Metamorphosis: Sea Urchins Pluteus Larvae Metamorphosis: Sea Urchins Primary Larvae: No trace of adult morphology Metamorphosis: Amphibians • Hormone(s): T3 and T4 • Four Major Morphological Processes – Growth of new structures – Cell death in existing structures – Remodeling of existing structures – Biochemical respecification • Shift in the genes expressed and the physiological functions they control Metamorphosis: Amphibians Tadpole eyes are on the sides of the head, frog eyes are on the front and top Binocular Vision New neurons differentiate and form new ipsilateral tracts Metamorphosis: Amphibians • Cell death in existing structures – Apoptosis • T3 induces apoptosis in tail and gills • Apoptosis occurs in gut epithelium due to ECM loss – Phagocytosis • Macrophages finish off the cells of the tail • Also destroy larval RBCs to make fresh ones with the adult hemoglobin protein Metamorphosis: Amphibians Remodeling: - Eyes - Skull - Skeleton - Gut - Sensory Metamorphosis: Amphibians Biochemical Respecification NH3 = ammonia, amino group NH4+ = ammonium ion T3 causes a shift in transcription factor expression that upregulates these genes. Metamorphosis: Amphibians 2NH3 + CO2 + H2O (urea) Actions of thyroxine (T4) and tri-iodothyronine (T3) Thyroid receptors are found in the nucleus where they are bound to inactive promoters. When thyroid hormone enters such a cell, it will bind to receptor and the combination is an active transcription factor for that specific gene. Cells that make high levels of deiodinase II are more responsive to thyroid stimulation. Cells that make high levels of deiodinase III are less responsive Actions of thyroxine (T4) and tri-iodothyronine (T3) • What genes you have your thyroid receptors on is very important to your function – Limb muscle cells grow in response to thyroid hormones – Tail muscle apoptoses in response to thyroid hormones • How much deiodinase II you make is very important – Limb buds make a lot and respond to early low levels of T4 – Tails make very little and wait for later very high levels of T4 – This is good! • How a tissue is organized before T4 is very important – Thyroid hormones make skin apoptose – Head and body have basal stem cells, tail does not • Your tail degenerates during week 4 of gestation in much the same fashion as the frogs! Metamorphosis: Amphibians • Some amphibian species have evolved alternatives to metamorphosis: Heterochrony – Neoteny: Normal gonadal maturity, retention of juvenile form – Progenesis: Accelerated gonadal maturity, retention of juvenile form – Direct Development: No larval form Metamorphosis: Amphibians Neoteny in the Mexican axolotl (salamander) Normal adult with juvenile form. Adult form not seen in nature, resulting from T4 in the pond. Metamorphosis: Amphibians Direct development in a common Puerto Rican frog Two views of the developing limb buds within the egg Frogs, not tadpoles, hatch from the eggs. Very large, nutrient-rich eggs allow skipping larval food gathering stage. Metamorphosis: Insects Adult = imago Metamorphosis: Insects • Larva are eating machines to provide energy for non-feeding pupal development • They have both doomed larval cells and rudiments of imaginal cells for adult – The larval cells will apoptose in the pupa – Imaginal disc cells will form wings, legs, antennae, eye, head, thorax and genitalia – Histioblasts will form adult abdomen – Imaginal cell clusters in each organ will form adult organ as the larval organ degenerates Metamorphosis: Insects Locations and developmental fates of imaginal discs and imaginal tissues in the third instar larva of Drosophila Metamorphosis: Insects Leg Imaginal Disc before pupa during pupa Epidermis cells form a hollow tube that coils Telescopes out in pupa Metamorphosis: Insects Metamorphosis: Insects A view into the minds of fly-guys.... The cells at center of the disc secrete Wingless (Wnt) and Decapentaplegic (TGF-B) This causes different expression levels of transcription factors Dachsund (green) and Distal-less (red). Metamorphosis: Insects • Like amphibians, control is hormonal in insects – Presence of juvenile-hormone makes a larval molt – Shift to steroid 20-hydroxyecdysone gives pupal molt – Differential timing in the development of pupal structures is due to 20E receptor expression timing Regeneration • Restoration of missing tissues • Post-embryonic reactivation of development • Occurs in some form in all species – Stem cell-mediated regeneration – Epimorphosis: adult cell de- and re-differentiation – Morphallaxis: adult cell repatterning – Compensatory regeneration: adult cell division Regeneration: Epimorphosis flatworms, salamanders The surviving cells lose their specification and form an undifferentiated tissue bulge Regeneration : Epimorphosis Vertebrate limb development from apical ectodermal cap (ridge) Regeneration: Morphallaxis The hydra is constantly regenerating cells that are sloughed off. Only the head and foot aren’t moving Regeneration: Morphallaxis • Any cells along the length can become head, body or foot • If you cut a hydra up all pieces will form all structures • Each piece has a dual gradient already established and both ends are specified Regeneration: Morphallaxis The bud always forms roughly the same distance anterior-posterior due to the combined gradient Regeneration: Compensatory Regeneration • No dedifferentiation occurs in liver regeneration • All five cell types produce more of themselves • • • • • hepatocytes duct cells fat-storing (Ito) cells endothelial cells Kupffer macrophages • Progenitor cell back-up plan: Oval cells Aging • Time-related deterioration of physiological functions necessary for survival and fertilization – Life span vs. senescence • Combination of: – Mutations – Environmental factors – Random epigenetic changes Aging We’ve learned not to die young as often more than we’ve extended life Aging Hutchinson-Gilford progeria Mutation repair deficiency Aging Low caloric intake is associated with long-life in all species. Figure 15.36 Differential DNA methylation patterns in aging twins (Part 1) Epigenetic alterations Figure 15.37 Methylation of the estrogen receptor gene occurs as a function of normal aging Can effect reproductive capacity directly