Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
Fatty acid metabolism wikipedia , lookup
Genetic code wikipedia , lookup
Plant nutrition wikipedia , lookup
Amino acid synthesis wikipedia , lookup
Biosynthesis wikipedia , lookup
Citric acid cycle wikipedia , lookup
Nucleic acid analogue wikipedia , lookup
Fatty acid synthesis wikipedia , lookup
Biochemistry wikipedia , lookup
15-Hydroxyeicosatetraenoic acid wikipedia , lookup
Specialized pro-resolving mediators wikipedia , lookup
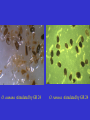
European Weed Research Society 2nd International Conference, Santorini, Greece September 7-10, 2009 Allelopathic relations in the rhizosphere between broomrapes and ordinary weeds Ana Höniges1, Franz Hadacek2, Aurel Ardelean1 & Klaus Wegmann1 1Vasile-Goldiş-University, Arad, Romania 2University of Vienna, Austria Allelopathy among plants The German botanist Hans Molisch coined the term allelopathy, derived from the Greek „allelon“ (the other) and „pathein“ (suffer), in order to describe negative effects on germination, growth and development of a different plant species. This definition is unbalanced, because also positive effects have become known. Rüdiger Knapp in 1950 created his research field „Experimental Sociology of Higher Plants“. Knapp never analyzed and identified chemical compounds. However, allelopathic effects are brought about by chemical compounds, which we call allelochemicals. Allelopathic effects by neighbour plants Allelopathic effects caused by plants in the neighborhood are well known. Germination stimulation of parasitic plants by strigolactones in root exudates of host or non-host plants belong to them. Germination inhibition by metabolites, exuded by roots or washout from leaves are observed. Among other, rarer metabolites, the cinnamic acid family (cinnamic, p-coumaric, ferulic acids) are well known as germination inhibitors. Phelipanche/Orobanche suppression by weeds? The observation that a weedy part of a heavily infested tobacco field in Germany was free of Phelipanche ramosa led us to the suspicion that allelopathic effects might be involved. Therefore we carried out analyses of root exudates. Methods Plants were cultivated in Vermiculite, then washed out and transferred into hydroculture in half-concentrated Murashige/Skoog medium. The water from the rhizosphere was collected during several days, concentrated in a rotation evaporator (50 °C), then preseparated into a hydrophilic phase (containing mainly sugars, polyols and organic acids) and a hydrophobic phase by an Amberlite XAD1180 column. This hydrophobic phase was analyzed by HPLC separation on a Dionex HPLC instrument equipped with a UVD340U photodiode array detector and data analysis. The absorption spectrum for each peak between 220-590 nm was recorded and compared with the data bank. The data bank proposes compounds for spectra with 95% or more identity with known compounds. Rising plants in Vermiculite Teucrium chamaedrys Galium verum Centaurea scabiosa Cirsium vulgare Hedera helix root exudate Benzoic acid Berberis vulgaris root exudate Benzoic acid Artemisia campestris root exudate Benzoic acid Pelargonium zonale root exudate Benzoic acid Root exudate of associated flora Benzoic acid Results of the analyses A metabolite is identified, if the elution time in the HPLC separation and the absorption spectrum are matching with that compound. Members of the cinnamic acid family have not been identified. Surprisingly, benzoic acid was a major component in many root exudates (see list). Benzoic acid in root exudates Artemisia campestris Berberis vulgaris Carduus personata Centaurea scabiosa Cirsium vulgare Galium verum Hedera helix Pelargonium zonale Teucrium chamaedrys Thymus pannonicus Examples with no or little benzoic acid in root exudates were Lycopersicon esculentum, Medicago sativa, Salvia aethiopis and Urtica dioica. Germination experiments Germination experiments have been carried out under standard conditions with seeds of Phelipanche ramosa and Orobanche cumana, two important weedy broomrape forms. Germination stimulant was the synthetic strigolactone analogue GR 24 (10-6 M). The following potential germination inhibitors were COOH COOH added (10-3 M). COOH O syringic acid O COOH H3CO O OCH 3 OH O O GR 24 CH3 t-cinnamic acid OCH 3 OH ferulic acid benoic acid O. cumana stimulated by GR 24 O. ramosa stimulated by GR 24 Results of the germination experiments Stimulant Dist. water (blank) GR 24 GR 24 + cinnamic acid GR 24 + ferulic acid GR 24 + syringic acid GR 24 + benzoic Phelipanche Orobanche ramosa cumana 0 0 96 85 5 23 3 22 5 7 20 40 Conclusions In previous studies the role of ordinary weeds for seed bank setup and spreading of Phelipanche ramosa has been studied, in particular their role as associated hosts in colza (Gibot-Leclerc et al. 2003). Suicide germination by germination stimulants in weed root exudates would lead to parasitic seed bank reduction, if the weeds are not parasitized. We report here about a potential for reducing parasitic weeds by germination inhibition by benzoic acid exuded by common weeds (or crop plants).