Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
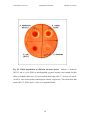
Laviad Sivan et al. 2015 supplementary material Chitinase Aeromonas Supporting information for accompanying manuscript entitled: Aeromonas chitinase degrades chironomid egg masses Sivan Laviad1, Amnon Golan2,3, Tamar Shaked1, Dalit Vaizel-Ohayon4, Malka Halpern1,2* and Elah Pick1,2* 1 Dept. of Evolutionary and Environmental Biology, Faculty of Natural Sciences, University of Haifa, Haifa, Israel; 2Dept. of Biology and Environment, Faculty of Natural Sciences, University of Haifa, Oranim, Kiryat Tivon, Israel; 3Protein purification center, Ramat Yohanan, Israel; 4Nesin Central Laboratory, Mekorot National Water Co. Ltd., Nazareth Illit, Israel. * Corresponding author: Elah Pick, Malka Halpern This file contains the following sections: 1) Appendix S1. Experimental procedures 2) A supplementary Table 3) Supplementary Figures 1 Laviad Sivan et al. 2015 supplementary material Chitinase Aeromonas Appendix S1. Experimental procedures Egg mass sampling and Aeromonas isolation Egg masses were sampled every other week between April and September 2009 at the Tivon waste stabilization pond (WSP), Israel. Styrofoam boards (25X25 cm) were used as artificial oviposition sites for adult females. Egg masses along the side of the board were collected 24h after placement of the board in the water habitat. The egg masses were brought to the laboratory immediately after collection. Three egg masses were chosen randomly for the bacterial isolation. To remove the bacteria that were not attached to the egg masses, egg masses were rinsed individually in sterile saline water (0.85% NaCl) and then vortexed for 1 min. This procedure was repeated five times. Each egg mass was then crushed in 1 ml sterile saline water, by means of a sterile glass homogenizer. The homogenate was diluted, and 0.1 ml was spread on m-Aeromonas agar plates (Biolife, Italy) for isolation of Aeromonas strains. Yellow colonies from the mAeromonas agar medium that were suspected of being Aeromonas were subcultured on LB agar five times to obtain clonal isolates. The identities of the isolates were further verified as described below. Bacterial isolates were kept in LB with 30% glycerol at 80°C. Identification of Aeromonas isolates Yellow colonies from m-Aeromonas selective agar medium (Biolife, Italy), which were suspected as Aeromonas, were identified by PCR assay in accordance with Kong et al. (1999). According to this assay, a specific intergenic region between the 16S rRNA and the 23S rRNA genes was replicated. Isolates that were found positive belonged to the Aeromonas genus. For species identification, the rpoD gene (RNA polymerase that encodes subunit D) was amplified and sequenced according to Soler et al. (2004). Newly determined sequences were compared with those available in the GenBank (http://www.ncbi.nlm.nih.gov) databases, by means of the standard nucleotide–nucleotide BLAST program (BLASTN; http://www.ncbi.nlm.nih.gov), to ascertain their closest relatives. These sequence data have been submitted to the GenBank database under accession numbers JN663897 – JN664025 and KM884830 – KM884833. 2 Laviad Sivan et al. 2015 supplementary material Chitinase Aeromonas EMD activity assay EMD activity was applied according to Halpern et al. (2003), with minor changes. In brief, 129 Aeromonas isolates obtained and identified from the egg masses were tested for their chironomid EMD skills. Bacterial isolates were grown in 10 ml LB medium in a shaker incubator at 30°C and 37°C for 24h and 48h. Culture supernatants were obtained by centrifugation (13,000 rpm. for 15 min) and were filtered through a 0.2 µm pore-size filter (Corning, NY), which resulted in a clear supernatant. EMD activity was determined by using 70% ethanol disinfected chironomid egg masses. Degradation assay was performed by incubation of one egg mass in 1 ml sterile supernatant at 35°C for up to 48h. Each test was carried out at least three times. A reaction was considered positive when the egg mass structure disintegrated and a large excess of free eggs was found separated from the egg mass, as compared with the LB medium control. EMD activity of egg masses hatching prevention was also tested in dilutions of fraction 9 by use of freshly collected egg masses instead of 70% ethanol disinfected egg masses. Egg masses were incubated at 35°C for up to 48h. EMD activity had been used to detect and purify the enzyme, and was tested after each step of purification (Fig. S1). At the stage of checking the partially purified enzyme, we defined one unit of the EMD activity as the amount of EMD factor that resulted in the egg mass structure disintegrating after 24h incubation at 35°C. Aeromonas dhakensis strain 3K1C15 (accession number EF634216) was isolated by Senderovich et al. (2008), and approached for the purification of the EMD factor. We chose this particular strain for the EMD purification experiment because it demonstrated a higher EMD activity than the other strains collected in the current study. Concentration of the EMD factor The optimal temperature for strain 3K1C15 growth on LB medium was 30°C for 24h with shaking (100 rpm). To obtain a sterile supernatant, strain 3K1C15 was grown for 24h. Then, it was centrifuged for 15 min at 13,000 rpm. The supernatant was filtered through a 0.2 µm pore-size filter (Corning, NY). The filtered supernatant was subjected to a two steps of (NH4)2SO4 precipitation at 4°C (Biewener, 1992). First, the filtrate was brought to 30% saturation, and the precipitated proteins were collected by centrifuging 3 Laviad Sivan et al. 2015 supplementary material Chitinase Aeromonas for 25 min at 12,000 rpm, 4°C. Next, the (NH4)2SO4 saturation was increased to 70%. The precipitate was collected by centrifuging for 25 min at 12,000 rpm at 4°C, and dissolved in 2 ml of 50mM Tris Buffer pH 8.5. The precipitate was then dialyzed with 25 mM Tris Buffer pH 8.5 with 50mM NaCl and 10% glycerol overnight at 4°C. The precipitate was concentrated with a 100 kD centricon, and then with a 30 kD centricon (Sartorius stedim biothech, Germany), by centrifuging at 7,000 rpm for 30 min at 4°C. The precipitate on the centricon was dissolved in 50mM Tris buffer pH 8.5 and fluidized to the original volume. EMD activity was measured at every step of this whole process as described. Screening for proteolytic enzymes by casein and gelatin zymograms After ultrafiltration with a 30 kD-cutoff membrane, the filtrate was loaded on 0.1% casein and 0.1% gelatin zymogram gels. Zymograms were carried out as described previously (Oldak and Trafny, 2005). Briefly, 10% SDS- polyacrylamide gels (Laemmli, 1970) were co-polymerized with 0.1% casein or gelatin (Becton Dickinson, USA). 10µl of protein samples were mixed with 15µl double distilled water (DDW) and 25µl of sample buffer (62.4 mmol l-1 Tris HCl pH 6.8, 2.5% SDS and 0.001% bromophenol blue), and 25µl were loaded on the gel. Electrophoresis was performed at 4°C and 150V, until the remnant of the loading stain reached down to the bottom of the gel. To remove the SDS, gels were washed with 2.5% Triton-X-100 (Sigma Aldrich, Germany) solution, for 45 min with shaking at room temperature. Then, the gels were transferred to the zymogram reaction buffer (50 mmol l-1 Tris–HCl pH 8.0 with 5 mmol l-1 CaCl2) for 30 min, with shaking at room temperature. Thereafter the gel was transferred to a new zymogram reaction buffer for 1 hour at 35°C, followed by staining with Coomassie Brilliant Blue R-250 (10% acetic acid, 50% methanol, 40% DDW and 0.1% Commassie R-250) (American Bioanalytical, USA), for 30 min, with shaking at room temperature. Coomassie staining was washed with a distaining buffer (25% methanol, 10% acetic acid and 65% DDW) until the appearance of transparent fields in the lanes. Transparent fields represent the absence of either gelatin or casein, which were digested by the partially purified proteolytic enzymes, fractionated in the gel. Enzymatic activity was quantified by the Image J program (http://imagej.nih.gov/ij/). Zymograms were used in the search 4 Laviad Sivan et al. 2015 supplementary material Chitinase Aeromonas for a correlation between protease and EMD activities, therefore was tested in the various mono-Q fractions (Fig. S1). Purification and identification of the Aeromonas EMD factor Purification of EMD factor was achieved in a few steps, as summarized in Fig. 2. Concentrated samples (see above) were loaded on a Heparin column attached to FPLC (ÄKTA, Amersham Biosciences) and activity was detected only in the “flow through” (FT) fraction. To yield a highly purified protein, a strong anion exchange Mono-Q column (5/50 GL, Amersham Biosciences) was applied for further separation. Heparin unbound fraction was diluted to a final volume of 50 ml with buffer A (20mM Tris pH 7.2, 50mM KCl, 10% Glycerol, 1mM β- Me) and loaded 3 times on the mono-Q column at a flow rate of 0.5 ml/min. Fractionation was achieved by a linear gradient of buffer B (20mM Tris pH 7.2, 1.5M KCl, 10% Glycerol, 1mM β- Me) at a flow rate of 1 ml/min. Apparently, the EMD factor was highly enriched in one specific fraction (Mono-Q fraction 9: Fig. 2), which contained 17% buffer B (comparable to 160-180 mM KCl). EMD and protease activities were obtained for all fractions. The active fraction and controls (non-active fractions) were sent for MS MS analysis at the Smoler protein research center, Technion, Israel (http://proteomics.net.technion.ac.il). Chitin presence in the egg masses Congo red dye (BDH, UK), which stains oligosaccharides, was taken as an indicator for chitin (Hageage and Harrington, 1984). Egg masses were stained with 1 mg/ml Congo red for 3h with gentle rotation, followed by 2 washes of 30 min with 50mM potassium phosphate buffer pH 6.0 for half an hour. Egg masses stained with Congo red were observed by light microscopy and pictures were taken. Calcofluor white (Pringle, 1991), a fluorochrome dye (cat. No. 18909; Fluka SigmaAldrich, Switzerland), was also used to confirm chitin presence in the egg masses. Calcofluor white binds with β-1,3 and β-1,4 polysaccharides. The dye was observed under UV light and pictures were taken. 5 Laviad Sivan et al. 2015 supplementary material Chitinase Aeromonas EMD activity using commercial chitinase Commercial chitinase (200 units/gr) extracted from Streptomyces griseus (cat. no. C6137; Sigma, Germany) was diluted with distilled water to a concentration of 1 mg/ml. EMD bioassay was applied for serial dilutions of the enzyme. One unit of chitinase activity is defined as the liberation of 1.0 mg of N-acetyl-D-glucosamine from chitin per hour at pH 6.0 at 25°C in a 2h assay. Chitin Degradation Assay Aeromonas isolates were cultured on 1% acid swollen chitin agar plates that were prepared out of practical grade chitin (Sigma, Germany), as described by Senderovich et al. (2010). Plates were incubated at 35°C for 24h and stained with 1.0 mg/ml Congo red (BDH, UK) for their ability to degrade chitin. Color was distained with 50mM buffer potassium phosphate pH 6.0 until a clear zone around the bacterial colonies showed the presence of chitinase activity. Induction of Aeromonas EMD factor with chitin Experiment I. 14 Aeromonas isolates were grown in 10 ml LB broth or LB broth supplemented with 0.5% chitin in a shaker incubator at 30°C for 24h. Culture supernatants were obtained by centrifugation (13,000 rpm. for 15 min) and filtration through a 0.22µm pore-sized filter (Corning, NY). EMD activity was determined as described above. Experiment II. A. dhakensis 3K1C15 and A. caviae EG4A were grown in five different media combinations: LB broth, LB broth supplemented with 0.5% chitin, LB broth supplemented with 0.5% glucose, Minimal Davis broth without Dextrose (BD, Difco, France) supplemented with 0.5% chitin and Davis broth without Dextrose (BD, Difco, France) supplemented with 0.5% glucose. The strains were grown in a shaker incubator at 30°C for 24h. Culture supernatants and EMD activities were treated and determined as described above. Colony forming units (cfu) were determined for all the cultures. Statistical analysis Repeated-measures ANOVA was used to determine whether Aeromonas spp. compositions/diversity vary between different samplings. Repeated-measures ANOVA 6 Laviad Sivan et al. 2015 supplementary material Chitinase Aeromonas was also applied to compare Aeromonas spp. abundances (i.e. number of isolates per species) in the different samplings. Mauchly's Test indicated that the assumption of sphericity was violated in both cases (X2=103.19, df=44, p<0.05), therefore the HyunFeldt correction was applied for calculating the F-statistic. References Biewener, A.A. (1992) In vivo measurement of bone strain and tendon force. In Biomechanics – Structures and Systems: A Practical Approach. New York: Oxford University Press: 123–147. Hageage, G., and Harrington, B. (1984) Use of calcofluor white in clinical mycology. Lab Med 15: 109-112. Halpern, M., Gancz, H., Broza, M., and Kashi, Y. (2003) Vibrio cholerae hemagglutinin/protease degrades chironomid egg masses. Appl Environ Microbiol 69: 4200-4204. Kong, R., Pelling, A., So, C. and Wu, R. (1999) Identification of oligonucleotide primers targeted at the 16S–23S rDNA intergenic spacers for genus-and species-specific detection of Aeromonads. Mar Pollut Bull 38: 802-808. Laemmli, U.K. (1970) Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 227: 680-685. Oldak, E., and Trafny, E.A. (2005) Secretion of proteases by Pseudomonas aeruginosa biofilms exposed to ciprofloxacin. Antimicrob Agent Chemother 49: 3281-3288. Pringle, J.R. (1991) Staining of bud scars and other cell wall chitin with calcofluor. Methods Enzymol 194: 732-735. Senderovich, Y., Gershtein, Y., Halewa, E., and Halpern, M. (2008) Vibrio cholerae and Aeromonas: Do they share a mutual host? ISME J 2: 276-283. Senderovich, Y., Izhaki, I., and Halpern, M. (2010) Fish as reservoirs and vectors of Vibrio cholerae. PLoS One 5: e8607. Soler, L., Yanez, M.A., Chacon, M.R., Aguilera-Arreola, M.G., Catalan, V., Figueras, M.J., and Martinez-Murcia, A.J. (2004) Phylogenetic analysis of the genus Aeromonas based on two housekeeping genes. Int J Syst Evol Microbiol 54: 15111519. 7 Laviad Sivan et al. 2015 supplementary material Chitinase Aeromonas A supplementary table Table S1. EMD and protease activities in various mono-Q fractions. Mono Q fractions were subjected for casein and gelatin zymograms, as well as for EMD activity. The results demonstrate that EMD activity is mostly present in fraction 9, while the peaks of casein and gelatin degradation activities are observed in fractions 6 and 8 respectively. Color is coded according to the relative activity. EMD Casein Gelatin Fraction 5 ++ ++ Fraction 6 +++ +++ Fraction 8 + ++ +++ Fraction 9 +++ + ++ Fraction 10 + + . 8 Laviad Sivan et al. 2015 supplementary material Chitinase Aeromonas Supplementary Figures Fig. S1. Extracellular proteases profiles of A. dhakensis 3K1C15, as seen by using 6% polyacrylamide gel, co-polymerized with casein or gelatin (Zymogram). Six different proteins that digested casein and eight different proteins that digested gelatin can be observed in the supernatant. Activity found in the gel was analyzed with Image J software. 9 Laviad Sivan et al. 2015 supplementary material Fig. S2. Chitin degradation by different bacterial species. Chitinase Aeromonas Isolates A. dhakensis 3K1C15 and A. caviae EG4A (a non-degrading egg mass isolate) were scanned for their ability to degrade chitin on a 1% acid swollen chitin agar plate. V. cholerae O9 and E. coli BL21 were used as positive and negative controls, respectively. The results show that strains 3K1C15, EG4A and V. cholerae O9 degraded chitin. 10 Laviad Sivan et al. 2015 supplementary material Chitinase Aeromonas Fig. S3. The ability of Aeromonas isolates to degrade chitin on a chitin agar plate. 47 non-degrading egg mass Aeromonas isolates were randomly chosen and cultured on agar plates supplemented with 1% acid swollen chitin. 85% of these isolates showed chitinase activity characterized by the ability to produce a clear zone around their colonies. 11