Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
West Nile fever wikipedia , lookup
Hepatitis C wikipedia , lookup
Marburg virus disease wikipedia , lookup
Human cytomegalovirus wikipedia , lookup
Henipavirus wikipedia , lookup
Herpes simplex virus wikipedia , lookup
Antiviral drug wikipedia , lookup
Hepatitis B wikipedia , lookup
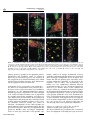
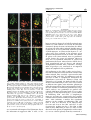
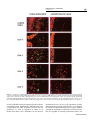
Journal of NeuroVirology (2000) 6, 359 ± 372 ã 2000 Journal of NeuroVirology, Inc. www.jneurovirol.com Abortive rabies virus central nervous infection is controlled by T lymphocyte local recruitment and induction of apoptosis Anne Galelli*,1, LeõÈla Baloul1 and Monique Lafon1 1 Unit of Neurovirology and Regeneration of Nervous System, Department of Virology, Pasteur Institute, 25 ± 28 rue du Dr Roux, 75724 Paris Cedex 15, France Nonfatal paralysis, induced by the attenuated Pasteur strain of rabies virus, is characterised by local and irreversible ¯accid paralysis of the inoculated limbs. We characterised the spread and localisation of virus in the CNS of infected mice, determined the nature of cell injury and examined the role of the immune response. Data indicate that infection of BALB/c mice induced paralysis in 60% of infected mice, the others recovering without sequelae. In both groups of mice, virus was detected in restricted sub-populations of neurons from the brain and spinal cord, and intensity of the neuropathology correlated with levels of rabies RNA and apoptotic infected neurons. However, apoptosis of neurons and paralysis were not due to a direct deleterious effect of the virus, but induced by a T-dependent immune response, as evidenced by their absence in nude mice. Paralysed and asymptomatic mice developed a similar rabies virus-speci®c IgG2a antibody response, thus excluding the role of any modi®cation of the humoral immune response. In contrast, three events were critically associated with the development of neurological symptoms: the amount of virus in the CNS, the level of apoptosis in both infected neurons and uninfected surrounding cells and the progressive parenchymal in®ltration of CD4+ and CD8+ T cells at the site of infection. These data suggest that during nonfatal rabies infection, the levels of viral replication and primary degeneration of infected neurons by apoptosis could be responsible for the in®ltration of T lymphocytes capable of inducing secondary degeneration of neural cells. Journal of NeuroVirology (2000) 6, 359 ± 372. Keywords: infectious viral immunity; neuroimmunology; T lymphocytes; apoptosis Introduction Rabies virus is a highly neurotropic virus (Murphy et al, 1973; Wunner, 1987) which induces in both humans and animals, two forms of disease (Gamaleia, 1887; Wunner, 1987). The most frequent manifestation of disease is an acute encephalitis which may be associated with spastic hind limb paralysis, thought to be a direct result of the damaging effect of the virus on neurons. Depending on viral pathogenicity, natural resistance of certain mouse strains, and site of inoculation, rabies infection can generate another form of disease, which is also characterised by paralysis of the limbs but is not due to a direct neuropathogenic effect of the virus (Baer et al, 1977; Iwasaki et al, *Correspondence: A Galelli Received 29 October 1999; revised 24 January 2000; accepted 27 March 2000 1977). This paralysis can be either ascending and fatal or remain local and nonfatal. Nonfatal paralysis is induced by virus strains of low pathogenicity and is clinically characterised by local and irreversible ¯accid paralysis of the inoculated limbs. Initial observations correlated the ef®cacy of the immune response with the induction of this form of paralysis (Iwasaki et al, 1977; Smith, 1981). Further studies using athymic or immunosuppressed mice established the importance of a virus-induced immunopathological process involving both CD4+ and CD8+ T cells in the development of paralysis (Smith et al, 1982; Sugamata et al, 1992; Weiland et al, 1992). Therefore, the immune response to rabies virus infection is thought to play a dual role, both promoting survival of the infected animals and exacerbating disease. However, the nature of the pathological Immunopathology of nonfatal rabies A Galelli et al 360 immune response remains poorly understood. Furthermore, probably due to differences in the observed clinical phenomena (extensor spasticity in ascending fatal paralysis or ¯accid nonfatal paralysis), controversial results were obtained concerning the nature and localisation of cell injury as well as the role of T cells (Chopra et al, 1980; Iwasaki et al, 1977; Sugamata et al, 1992; Weiland et al, 1992). To resolve these contradictions and discriminate between the deleterious effects of the immune response and the direct neuropathogenic effect of rabies virus, we established a model of nonfatal paralysis. BALB/c mice were intramuscularly inoculated with the Pasteur strain of the rabies virus (PV) which induces a non-lethal infection characterised by the development of paralysis in about 60% of the infected mice, the others recovering without sequelae (asymptomatic). We ®rst characterised the spread and localisation of rabies virus in the central nervous system (CNS), determined the nature of cell injury, then examined the role of suppressor cells on the peripheral immune response. Finally, we isolated in¯ammatory cells in®ltrating the CNS, evaluated their CD4+ and CD8+ T cell content and analysed their ability to express cytokine genes. Our results provide strong evidence that infection of BALB/c mice with the PV strain of rabies virus induced a localised infection resulting in apoptosis of infected neurons and that development of paralysis was associated with increased virus replication and apoptosis of both infected and non-infected cells in the CNS. However, paralysis was not due to the direct neuropathogenic effect of rabies virus, but correlated with the in®ltration and persistence at the site of infection of a population of activated mononuclear cells which contained a majority of CD8+ T cells and expressed high levels of mRNA for speci®c cytokines with the capacity to exert deleterious effects on neural cells. Results Clinical disease of BALB/c mice infected with the PV strain of rabies virus Normal BALB/c mice infected i.m. with 107 infectious particles of the PV strain of rabies virus appeared healthy for the ®rst 3 days of infection, then all developed non-speci®c signs of illness characterised by apathy and rough fur. From day 7 of infection, mice either recovered without sequelae (asymptomatic) and remained healthy or developed neurological signs of disease characterised by tremor, ataxia, and a reduction or total loss of hind leg motility. Before day 7, mice subsequently paralysed were indistinguishable from mice subsequently asymptomatic. As shown in Figure 1A, about 60% of infected normal BALB/c mice developed ¯accid paralysis between days 7 and 11 of infection, but no mice died of infection (Figure Journal of NeuroVirology Figure 1 Pathology induced by infection of BALB/c mice with the PV strain of rabies virus. Groups of eight mice were inoculated i.m. with 107 infectious particles in both hind legs. Paralysis (A) was de®ned as the loss of hind leg mobility. Lethality (B) was expressed as the cumulative percentage of mortality. +/+ BALB/c mice (* ± *), CD8+ T-depleted BALB/c mice (* ± *), and nu/nu BALB/c mice (~ ± ~). 1B). This paralysis was restricted to the inoculated limbs and was irreversible during an observation period of several months. When BALB/c mice were depleted of CD8+ T cells, they all recovered from infection without sequelae. In contrast nu/nu BALB/c developed cachexia, died between days 12 and 14 of infection, but never developed neurological symptoms. These data showed that PV infection of BALB/c mice induced a nonfatal paralysis and con®rmed that CD4+ T cells are required for resistance to PV rabies virus while CD8+ T cells are involved in the development of paralysis. Development of paralysis is correlated with increased but controlled viral replication in the CNS We ®rst characterised the spread and replication of rabies virus in the CNS of infected BALB/c mice. To exclude possible interference by rabies-speci®c antibodies in the detection of infectious virus, we evaluated the presence of rabies-speci®c genomic RNA by semi-quantitative RT ± PCR. The same amount of RNA was ampli®ed for 30 cycles both for viral RNA and the housekeeping gene GAPDH, and expression of viral RNA determined relative to that of GAPDH mRNA. As shown in Figure 2A, rabies-speci®c RNA expression was detected in the spinal cord of all infected normal BALB/c mice by day 4 and a signi®cant difference was observed by day 7, between asymptomatic and paralysed mice. While rabies RNA expression progressively decreased in asymptomatic mice and disappeared by day 12, development of neurological signs was correlated with increased expression of rabies RNA, which remained elevated on day 11 and then decreased signi®cantly. Rabies RNA was still detected in the spinal cord from paralysed mice at the end of the observation period. Furthermore, from days 7 to 11 after infection, a correlation was observed between the expression of viral RNA in Immunopathology of nonfatal rabies A Galelli et al 361 Figure 2 PCR ampli®cation of PV rabies nucleoprotein genomic RNA extracted from the CNS of infected mice. Relative rabies genomic RNA expression was determined as a ratio of the viral expression to that of the housekeeping gene GAPDH and evaluated in spinal cord (A) and brain (B) of either asympto± ) or mice developing neurological signs of matic mice ( disease ( ± ). Each point represents the arithmetic mean+s.d. of results from 3 ± 6 mice. the spinal cord and the intensity of neurological symptoms. For example on day 9, relative rabies RNA expression mean values were 0.22+0.05 for asymptomatic mice, 0.41+0.07 for mice with partial hind limb paralysis and 1.31+0.02 for mice with complete hind limb paralysis. Rabies RNA expression was also observed in the brain of all infected mice (Figure 2B), and development of neurological signs of disease was correlated with earlier and increased expression of rabies RNA, which was detected on day 7 and disappeared on day 13. These results excluded the possibility that, in asymptomatic mice, the virus was eliminated before invading the CNS, and showed that paralysis was correlated with increased viral replication. However, this replication was limited and the virus was progressively cleared from the CNS of paralysed mice. On day 12, rabies RNA was undetected in spinal cord and brain from CD8 depleted BALB/c mice. In contrast, on day 12, nude mice expressing high levels of rabies RNA (relative rabies RNA expression: 1.49 and 2.29 for the spinal cord and the brain, respectively) died, but did not develop paralysis. Therefore, difference in viral expression may contribute to, but is not the only crucial factor involved in the development of paralysis. Localisation of rabies virus in the CNS We next compared the localisation of rabies virus antigen in the CNS of asymptomatic and paralysed BALB/c mice. In both groups of mice, rabies virus NC was detected in the large neurons of the spinal cord (Figure 3A). In the brain, rabies virus NC was restricted to scattered neurons of the brain stem and to some Purkinje cells of the cerebellum, which were heavily infected (Figure 3B, C). Paralysed and asymptomatic mice differed in the number of infected neurons. On day 7, a mean value of 76+17 infected Purkinje cells was detected in 12 cerebellar sections from paralysed mice (n=2), whereas no infected Purkinje cells were detected in the same number of cerebellar sections from asymptomatic mice (n=2). The percentages of infected neurons detected on the same number of spinal cord sections were 3.2% for asymptomatic mice versus 15.5% for paralysed mice. Restricted infection in the brain explained the relatively low viral RNA expression detected using total RNA extracted from whole brain (Figure 2B). In contrast, as shown in Figure 3E ± H, we observed widespread infection of rabies virus in the brain of nu/nu BALB/c mice. Cell death during nonfatal rabies infection Apoptosis of infected neurons has been implicated in the pathogenesis of acute rabies encephalitis (Jackson and Rossiter, 1997; Theerasurakarn and Ubol, 1998). We therefore investigated the possible involvement of apoptosis in the development of nonfatal paralysis. At different times after infection, CNS tissue sections from both paralysed and asymptomatic mice were tested simultaneously for apoptosis by the TUNEL method and for the presence of rabies antigen. TUNEL positive cells were undetected in the CNS from control mice (data not shown). In contrast, as shown in Figure 4, TUNEL staining was observed in CNS sections from paralysed mice and was restricted to areas positive for rabies antigen staining (Figure 4A ± F). TUNEL staining was observed in Purkinje cells of the cerebellum (Figure 4B ± F), in scattered neurons of the brain stem and in motor neurons of the spinal cord (data not shown). In infected Purkinje cells of the cerebellum, apoptosis led ®rst to the disappearance of cell bodies (Figure 4D), followed by the progressive elimination of infected dendrites (Figure 4E, F). However, only a limited number of cells were positive both for TUNEL and rabies antigen (Figure 4A ± C). These infected TUNEL-positive neurons were surrounded by a signi®cant number of cells strongly positive for TUNEL only. Cells positive for TUNEL only were ®rst observed in areas of the granular layer of the cerebellum close to the cell bodies of infected Purkinje cells (Figure 4A, C), then around infected dendrites (Figure 4D, E). In the CNS from paralysed mice, apoptosis was observed between days 7 and 9 in the spinal cord and from days 8 to 10 in the brain. In asymptomatic mice, moderate numbers of cells positive for TUNEL and rabies antigen were also observed on days 8 and 9 in the spinal cord (data not shown) and in the cerebellum (Figure 4G). In contrast, despite widespread infection of their CNS, nude mice exhibited very few TUNEL-positive cells (Figure 4H, I). Recent reports suggested that cells containing DNA fragmentation are not necessarily apoptotic and that TUNEL staining alone should not be considered as satisfactory evidence for apoptosis. TUNEL-positive sections were therefore tested for the presence of activated caspase-3, the central Journal of NeuroVirology Immunopathology of nonfatal rabies A Galelli et al 362 Figure 3 Localization of rabies virus in the CNS 7 days after infection. Parasagittal brain sections and longitudinal spinal cord sections were double immunostained for rabies virus NC (green) and NeuN (red) speci®c for most neurons, except Purkinje cells. In +/+ BALB/c mice, anti-rabies virus NC staining was seen in motor neurons of the spinal cord (A), and Purkinje cells of the cerebellum (B, C), but not in the hippocampus (D). In nu/nu BALB/c mice, rabies virus NC staining was detected in all regions of the CNS, including neurons of the spinal cord (E), Purkinje and granular layer cells of the cerebellum (F), neurons of the cortex (G), and hippocampal neurons (H). Photographed with a 406 objective. effector protease speci®c to the apoptotic process (Thornberry and Lazebnik, 1998). As shown in Figure 4J ± L), activated caspase-3 was detected in the cytoplasm of all infected Purkinje cells which retained their cell body integrity, thus demonstrating the apoptotic nature of cell death. Comparable levels of anti-rabies virus antibodies are induced in asymptomatic and paralysed mice Previous studies suggest that rabies virus antibodies participate in the immunopathological process leading to paralysis (Sugamata et al, 1992; Weiland et al, 1992). Clearance of rabies virus is dependent on speci®c neutralising antibodies and their isotype. Differential viral clearance observed between asymptomatic and paralysed mice could be related to their particular antibody response to the virus. We therefore analysed, during the course of infection, development of the rabies-speci®c humoral immune response in individual mice. Results presented in Figure 5 show that asymptomatic and paralysed BALB/c mice developed comparable levels of total IgG anti-rabies virus-speci®c antiJournal of NeuroVirology bodies. Analysis of isotype distribution of these antibodies demonstrated that both types of infected mice developed comparable levels of IgG2a antibodies but did not produce IgG1 anti-rabies virusspeci®c antibodies. Similarly, at the peak of the response, titre of rabies virus neutralising antibodies was not signi®cantly different between asymptomatic mice (7.1+0.4 IU/ml) and paralysed mice (7.2+1.2 IU/ml). These results exclude the role of a defect or a switch in the humoral response in the immunopathological process involved in the development of paralysis. Levels of IgG anti-rabies virus-speci®c antibodies were also determined in CD8+ T cell depleted mice and in nude mice on day 12 after infection. CD8-depleted mice developed a level of IgG2a comparable to that of normal mice (4.2+0.1 versus 4.3+0.1), whereas nude mice did not develop rabies virus-speci®c IgG antibodies. Absence of suppressor cells in the peripheral lymphoid organs from paralysed mice We then examined the possibility that, in infected mice, development of neurological signs of disease Immunopathology of nonfatal rabies A Galelli et al 363 Figure 5 Asymptomatic and paralysed mice develop comparable levels of rabies virus-speci®c antibodies. Rabies-speci®c total IgG and IgG2a from mouse sera were measured by ELISA. Individual titres were expressed as the log10 of the highest dilution giving a value twice as high as that observed with normal mouse serum. Each point represents the arithmetic mean+s.d. of results from 3 ± 6 mice. been previously observed in lethally infected mice (Hirai et al, 1992; Wiktor et al, 1977a). We therefore examined, during the course of infection, the ability of spleen cells from rabies infected BALB/c mice to respond to an in vitro stimulation with non-speci®c T and B mitogens. As illustrated in Figure 6, 26105 spleen cells were cultured in the presence of an optimal concentration of Con A or LPS. The ability of spleen cells from infected mice to proliferate varied during the course of infection. However, there was no signi®cant difference in spleen cell proliferation between infected mice which developed neurological signs of disease and infected asymptomatic BALB/c mice. Therefore, development of paralysis was unrelated to an impaired ability of lymphoid cells to proliferate. Figure 4 Cell death in the CNS of rabies virus infected BALB/c mice. Double immunostaining for rabies virus NC (green) and TUNEL positive nuclei (red) in the cerebellum of paralysed (A ± F) or asymptomatic (G) +/+ BALB/c mice and in the spinal cord (H) and cerebellum (I) of nu/nu BALB/c mice, after infection by the PV stain of rabies virus. Detection of rabies virus NC (A), TUNEL-positive nuclei (B), and co-localisation of NC and TUNEL (yellow) in the Purkinje cell bodies from paralysed mice (C). Disappearance of infected Purkinje cell bodies (D, arrow), followed by the appearance of TUNEL-positive cells around infected dendrites (E), then by the elimination of infected dendrites and TUNEL-positive cells in the cerebellum of paralysed mice (F). Double immunostaining for rabies virus NC (green) and activated caspase-3 (red) in the cerebellum of paralysed +/+ BALB/c mice (J ± L). Detection of rabies NC (J), activated caspase-3 (K) and co-localisation of rabies virus NC and activated caspase-3 (L) in the Purkinje cell bodies from paralysed mice, at day 8 of infection. Photographed with a 406 (A ± I) or a 606 objective (J ± L). was associated with impaired T cell function, due to the induction of suppressor CD8+ T cells, as has Cell in®ltration in the CNS of rabies infected mice Previous histological studies on brain sections from rabies infected mice revealed a perivascular in®ltration of CD4+ and CD8+ T lymphocytes and the presence of CD8+ T lymphocytes throughout the parenchyma (reviewed in; Wunner (1987) and Sugamata et al (1992)). To further de®ne the role of these CNS in®ltrating T cells, cells were isolated at different time points after infection, phenotypically characterised by FACS and quanti®ed. Due to the low numbers of in®ltrating mononuclear cells, FACS analysis was performed on pooled cells isolated from brain and spinal cord of 2 to 3 mice per group. As illustrated in Figure 7, a low number of in®ltrating CD4+ and CD8+ T lymphocytes (600+87 and 290+65, respectively) was steadily isolated from the CNS of uninfected control BALB/c mice. After infection a modest but signi®cant increase in the number of in®ltrating CD4+ and CD8+ T cells was observed in all infected mice (P50.02 and P50.01 for CD4+ and CD8+ T cells respectively), peaking on day 5. The CD4 to CD8 ratio of these in®ltrating T cells was close to that observed in the spleen or lymph nodes of these mice (1.8 versus 2 for the spleen). From day 7 onwards, a Journal of NeuroVirology Immunopathology of nonfatal rabies A Galelli et al 364 Figure 6 Spleen cells from asymptomatic and paralysed rabiesinfected BALB/c mice proliferative in response to non-speci®c mitogens. 26105 spleen cells/well were stimulated with an optimal concentration of either ConA (1 mg/ml) of LPS (10 mg/ml). The proliferative response was assessed on day 3. Results are expressed as the mean of c.p.m.+s.d. of triplicates from 3 ± 6 mice. Mean c.p.m. in unstimulated cultures: 812+393 for asymptomatic mice and 1018+609 for paralysed mice. by day 14 the number of CD4+ T cells had declined whereas the number of CD8+ T cells remained constant. Analysis on day 12, of the distribution of T cells in brain and spinal cord indicated that T cells present in the brain (236103 CD4+ and 30.86103 CD8+) outnumbered T cells present in the spinal cord (12.86103 CD4+ and 18.66103 CD8+). In a previous report, in¯ammatory cell in®ltration in the CNS had been correlated with protection against infection (Weiland et al, 1992). We therefore compared T cell in®ltration in the CNS of rabies virus infected BALB/c mice with that induced in CBA/J mice, a strain resistant to rabiesinduced infection and paralysis (Lafon and Galelli, 1996). As shown in Figure 7, the pattern of T cell in®ltration in these mice was similar to that of asymptomatic BALB/c mice: only a low and transient in®ltration of both CD4+ and CD8+ T cells was observed in all infected CBA/J mice. Together these data demonstrate that all CBA/J and BALB/c mice infected with the PV strain of rabies virus developed a CNS T cell in®ltration but that the pattern level of in®ltration was different for paralysed mice. Development of paralysis correlated with increased and sustained in®ltration in the CNS by both CD4+ and CD8+ T lymphocytes, with the latter subset becoming predominant. Figure 7 T cell in®ltration in the CNS of rabies virus-infected mice. Mononuclear cells in®ltrating the CNS were puri®ed on a Percoll gradient and phenotypically characterised by two-colour staining with either anti-CD4 or anti-CD8 and anti-Mac-1 at the indicated times after infection. Each point represents the arithmetic mean+s.d. of staining results from 2 ± 4 experiments performed on pooled in®ltrating cells isolated from the brain and spinal cord from 2 ± 3 mice per group. T lymphocytes accumulate at the site of infection To determine the localisation of in®ltrating T cells in the CNS, sequential frozen sections from brain and spinal cord were analysed for viral antigen and for CD4+ or CD8+ cells by immuno¯uorescence. During the ®rst phase of cell in®ltration observed in all infected mice, CD4+ and CD8+ were detected perivascularly (data not shown). In contrast, in mice developing neurological signs of disease, T lymphocytes accumulated at the site of infection in brain parenchyma (Figure 8B, C) and in the spinal cord (Figure 8D, E). CD8+ T cells appeared more abundant than CD4+ T cells, consistent with FACS analysis of mononuclear cells puri®ed from the CNS. Low numbers of CD4+ and CD8+ T cells were observed in the brain (Figure 8G, H) and spinal cord (Figure 8I, J) of asymptomatic mice. These observations con®rmed our above mentioned results showing that paralysis is correlated with an increased accumulation of T cells in the CNS. Furthermore their localisation at the site of infection explained the relatively low level of cells obtained after dissociation of the CNS, due to the restricted spreading of rabies virus in the CNS of BALB/c mice. clear difference was observed between asymptomatic and paralysed BALB/c mice (P50.005). CNS T cell numbers declined in mice becoming asymptomatic but remained higher than in uninfected control mice during the observation period (P50.02 and P50.01 for CD4+ and CD8+ T cells respectively). In contrast, in mice developing neurological signs of disease, the number of T cells continued to increase as paralysis developed and correlated with the severity of the disease. Furthermore, this increase was characterised by a progressive diminution of the CD4 to CD8 ratio (1.7 on day 8 versus 0.53 on day 14), with CD8+ T cells outnumbering CD4+ T cells. Therefore a different pattern of in®ltration was observed for CD4+ and CD8+ T cells. Furthermore, Journal of NeuroVirology Messenger RNA expression of mediators involved in antiviral immunity by CNS in®ltrating mononuclear cells from paralysed mice With the aim of de®ning the role of the CNS in®ltrating T cells, we ®rst investigated a possible interaction between CD8+ T cells and infected neurons. Experiments measuring anti-rabies-CTL Immunopathology of nonfatal rabies A Galelli et al 365 Figure 8 Localisation of in®ltrating T cells in the CNS of paralysed and asymptomatic rabies virus infected mice. Cryostat parasagittal sections of brain (A ± C and F ± H) and longitudinal sections of spinal cord (D, E, I, J) were double immunostained for NeuN (red) and either rabies virus NC or CD4 or CD8 (green). Detection by immuno¯uorescent staining for rabies virus NC (green) (A and F) and in®ltrating CD4+ (B, D, G, I) and CD8+ (C, E, H, I) T cells in the cerebellum (A ± C) and the spinal cord (D, E) of paralysed mice or in the cerebellum (G, H) and the spinal cord (I, J) of asymptomatic mice, 12 days after infection. Photographed with a 406 objective. activity in BALB/c mice are impeded by the lack of a susceptible target. Furthermore, in¯ammatory cells isolated from the CNS of paralysed mice did not proliferate in vitro in response to ConA or to inactivated rabies virus. Therefore, in the absence of functional assays to assess the speci®city and the direct cytotoxic activity of these cells, we evaluated the ability of CNS in®ltrating cells to express mRNA speci®c for mediators essential for antiviral activity but also capable of exerting detrimental effects on Journal of NeuroVirology Immunopathology of nonfatal rabies A Galelli et al 366 neuronal function. Messenger RNA expression was evaluated by semi-quantitative RT ± PCR, relative to the expression of GAPDH on the same sample. As previously indicated for FACS analysis, mRNA cytokine expression was evaluated on pooled cells isolated from the brain and spinal cord of 2 to 3 mice per group. As shown in Figure 9, the expression of two pro-in¯ammatory cytokines with antiviral activity, IFN-g and TNF-a, were up-regulated at different time points of maximum in®ltration but declined at day 14. The expression of IL-2, IL-4, and IL-10 mRNA was also evaluated, but no mRNA was detected (data not shown). Viral antigen recognition by CTLs leads to the release of chemokines such as MIP-1a, MIP-1b, RANTES and IP-10 at the site of infection and the production of these mediators has been directly related to cytolytic function (Biddison et al, 1997; Price et al, 1999). CNS-in®ltrating cells from paralysed mice were therefore tested for their ability to express these chemokine mRNAs. As illustrated in Figure 9, high levels of mRNA of these mediators, essential for ef®cient ampli®cation of the immune response and able to exert deleterious effects on neural cells, were expressed by immune cells in®ltrating the CNS of paralysed mice. Discussion The present study describes a model of nonfatal paralysis induced in BALB/c mice by the Pasteur strain of rabies virus. In immunocompetent mice this infection induces a local and irreversible ¯accid paralysis of the inoculated limbs in about 60% of the infected mice, the rest recovering without sequelae. We analysed the spread of rabies virus in the CNS, the nature of injury and the role of T cells in this process. We demonstrated, using CD8+ T-depleted mice and nude mice, that development of paralysis was not due to a direct deleterious effect of virus on neurons, but linked to an immunopathologic process involving CD8+ T cells and apoptosis. Development of paralysis was dependent on three linked events: the levels of infection and apoptosis in the CNS and the progressive parenchymal in®ltration of CD4+ and CD8+ T cells at the site of infection. Furthermore, the effect of these three phenomena correlated with the intensity of neurological symptoms. In contrast, in asymptomatic BALB/c, the absence of neurological signs of disease was associated with low levels of infection and apoptosis and a modest and transient in®ltration of the CNS by both CD4+ and CD8+ T cells. In all infected +/+ BALB/c mice, rabies virus antigens were detected in motor neurons of the spinal cord and in restricted sub-populations of neurons in the brain, and the intensity of neurological symptoms correlated with levels of rabies RNA. As previously observed with virulent strains (Jackson and Rossiter, 1997; Theerasurakarn and Ubol, 1998), infection by the PV attenuated strain of rabies virus rendered neurons susceptible to Figure 9 RT ± PCR detection of mRNAs for soluble mediators of antiviral immunity in mononuclear cells in®ltrating the CNS from paralysed BALB/c mice. Relative expression of cytokine-speci®c mRNA was expressed as ratio of the mediator transcript to that of the housekeeping gene GAPDH. Each point represents the value obtained with pooled cells from three mice per group. RNA extracted uninfected BALB/c mice spleen cells was used for day 0. Journal of NeuroVirology Immunopathology of nonfatal rabies A Galelli et al 367 apoptosis and the intensity of neurological symptoms CNS also correlated with the number of apoptotic infected neurons in the CNS. However, in contrast to virulent rabies virus activity (Theerasurakarn and Ubol, 1998), apoptosis induced by the PV strain of rabies virus was not due to a direct deleterious effect of the virus but to a T-dependent immune response triggered by the virus, as shown by the absence of apoptosis in the heavily infected CNS of nude mice. These data con®rmed results from Smith et al (1982), who did not observe necrosis or neuronophagia in the CNS of immunosuppressed rabies-infected mice, despite massive infection of their CNS. The absence of paralysis in CD8+ T-depleted mice and the site speci®c recruitment of a disproportionate number of CD8+ T cells at the site of infection in the CNS of paralysed mice also suggested that these cells are of crucial importance in CNS injury. Several pathways may trigger a deleterious CD8+ T cell-mediated immune response. As shown in our experiments using CD8+ T-depleted mice and also observed by Dietzschold et al (1992), rabies virus is cleared by speci®c antibodies without the help of the virus-speci®c cell-mediated immune response. However virusspeci®c CD8+ T cells can be induced, contribute to, and regulate the response to infection and its consequences (Wiktor et al, 1977b). Because paralysed mice developed a speci®c antibody response identical to that of asymptomatic mice, we can exclude the possibility that CD8+ suppressor T cells impair the speci®c humoral antiviral response in paralysed mice. As previously reported (Murphy et al, 1973; Wunner, 1987) and con®rmed in this study, rabies virus exclusively infects neurons in the nervous system. To explain the role of CD8+ T cells, one hypothesis could be that, in rabies infected mice which become paralysed, T cells accumulate in the brain because they recognize rabies peptides expressed at the surface of infected neurons in association with MHC class I molecules and that CNS injury might result from the destruction of infected neurons by CTLs. Neurons of uninfected mice do not express MHC class I molecules, and there are con¯icting reports concerning their ability to up-regulate MHC class I expression after infection or exposure to in¯ammatory cytokines (Duguid and Trzepacz, 1993; Joly et al, 1991; Massa et al, 1993; Momburg et al, 1986; Neumann et al, 1997; Sethna and Lampson, 1991; Thomas et al, 1997; Wong et al, 1984). Furthermore, even if they can be induced to express MHC class I molecules, neurons may be resistant to cell-mediated lysis (Oldstone et al, 1986). We failed to detect MHC class I expression on rabies-infected neurons, suggesting that direct lysis of infected neurons by speci®c CTLs can not account for the immunopathology observed within the CNS following infection and that other viral antigen presentation pathways may exist. The sequence and the nature of events leading to paralysis is unknown, but our results suggest that the number of infected neurons dying by apoptosis could be a crucial event. Surprisingly, apoptosis of infected neurons was observed both in paralysed mice and in mice recovering without sequelae. In asymptomatic mice, apoptosis was limited and led to the rapid elimination of infected neurons. These results support the view that apoptosis of infected neurons may have a physiological role in protecting the CNS from the progression of infection and allowing contact between virus and immune components. They also argue in favour of the results of Morimoto et al (1999), suggesting that neuronal apoptosis is a host defence mechanism to limit the infection. The events which trigger apoptosis of infected neurons have yet to be determined. The observation that a low number of mononuclear cells in®ltrate the CNS of all infected immunocompetent mice during the ®rst week of infection and the results from Hooper et al (1998) showing that T-cell mediated, IFN-g dependent CNS in¯ammation collaborates with antibodies to clear virus from the CNS, suggest that the in®ltrating cells may induce this clearing event. However, in paralysed mice, the number of TUNEL-positive infected neurons was elevated and these cells were surrounded by a huge number of non-infected TUNELpositive cells which may be either apoptotic cells or scavenger cells containing apoptotic material. While apoptosis is regarded as the most ef®cient mechanism to eliminate cell debris or unwanted cells without inducing deleterious effect on surrounding tissue, cells dying by apoptosis may trigger damaging immune responses (Ren and Savill, 1998). Apoptotic bodies are recognised and engulfed by scavenger phagocytes (Savill et al, 1993) and their processing may generate epitopes recognised by speci®c CTLs (Bellone et al, 1997; Ronchetti et al, 1999). Although this presentation normally induces tolerance to these antigens, it can also lead to the induction of virus-speci®c CTLs, depending on the relative number of cells undergoing apoptosis at a given time and on the type of scavenger phagocytes involved in their clearance (Albert et al, 1998). Rabies virus-infected Purkinje cells seemed particularly vulnerable to apoptosis and the huge amount of infected dendrites remaining in CNS tissue after the disappearance of apoptotic cell bodies as well as the observation that higher numbers of infected neurons and apoptotic cells are present in the CNS of paralysed mice, as compared to asymptomatic mice, may signify that the mechanism involved in the removal of these cells is insuf®cient and lead to the accumulation of apoptotic material. Our results showing that inJournal of NeuroVirology Immunopathology of nonfatal rabies A Galelli et al 368 fected neurons undergo apoptosis suggest that viral antigens contained in apoptotic cells are accessible to cross-priming, thus providing MHC class I restricted Ags (Bevan, 1976; Jondal et al, 1996). The subsequent lysis by CTLs of the APC presenting the viral Ags could amplify the process of apoptosis and explain the presence of non-infected TUNEL positive cells in the vicinity of dying infected neurons in the CNS of paralysed mice. CTL induction by cross-priming requires CD4+ T cell help (Bennett et al, 1997), and this may explain the presence of these cells in the parenchyma. Inef®cient phagocytosis of cells dying by apoptosis may also induce harmful in¯ammatory responses. The mechanism by which T cells mediate injury has yet to be determined, but our results show that part of their function could be to secrete proin¯ammatory cytokines and chemokines. Cytokines alone, or in concert with chemokines may amplify the recruitment of T cells and macrophages to the site of infection and tissue injury. Activation of cerebral endothelial cells and subsequent modi®cation of the permeability of the BBB (Ludowyk et al, 1992; Sethna and Lampson, 1991) by these mediators could facilitate the recruitment of auto-reactive T cells which recognise self antigens produced by degeneration of the infected neurons (Wekerle, 1993). Cytokines may also act through activation of resident neural cells. Neural cells express receptors for chemokines, and these molecules could induce detrimental effects on neuronal function (Glabinski and Ransohoff, 1999). IFN-g is a potent activator of microglia, inducing upregulation of MHC class II, phagocytosis and production of cytokines (Merrill and Benveniste, 1996). Cytokines may also contribute to development of paralysis by directly damaging neural cells or indirectly via activation of macrophages and microglial cells which secrete factors toxic for oligodendrocytes such as nitric oxide (NO) and TNF-a (Merrill et al, 1993; Selmaj et al, 1991; Stoll et al, 1993; Wallach, 1997; Zajicek et al, 1992). Thus, IFN-g can directly injure oligodendrocytes (Vartanian et al, 1995). Therefore, Cytokines and chemokines produced by T cells could initiate the synthesis, by both in®ltrating and resident cells of a cascade of molecules, capable of exerting detrimental effects on neuronal activity, and resulting in secondary degeneration of the uninfected surrounding cells. Studies are in progress to de®ne the speci®city and the role of T cells in®ltrating the CNS of rabiesinfected mice and to determine which of these mechanisms are involved in the induction of apoptosis and development of paralysis. In conclusion, our results suggest that immune-mediated apoptosis of infected neurons and induction of rabies-speci®c CTLs by cross-priming of viral Ags could initiate a cascade of deleterious events in the neighbouring cells that escape the primary injury. Journal of NeuroVirology Epidemiological studies suggest a role of viral infections in the aetiology of various neurological diseases. Therefore, this model of nonfatal paralysis induced by peripheral infection with a neurotropic virus may be very helpful in understanding the mechanisms of damaging immune responses in the CNS. Materials and methods Rabies virus infection of BALB/c mice, assessment of clinical disease and preparation of cell populations Six-week-old +/+ BALB/c, nu/nu BALB/c and +/+ CBA/J female mice were purchased from Janvier (St. Berthevin, France). BALB/c mice were depleted of CD8+ T cells by 10 daily i.v. injections (beginning 5 days before infection) of 100 mg puri®ed anti-CD8 antibody (53-6.72 rat hybridoma). Depletion was checked on spleen cells, by ¯ow cytometry analysis (FACS), in a pilot experiment and on all infected mice at the end of the experiment. Groups of eight mice were inoculated intramuscularly (i.m.) with 16107 infectious particles of rabies virus Pasteur Virus strain (PV4) in both hind legs. Neurological symptoms were assessed by evaluating the animal's ability to support its body weight with its hind legs. Paralysis was de®ned as the total loss of hind leg mobility. At different times after infection, mice were anaesthetised with Hypnorm (Janssen, Oxford, UK) and perfused by intracardiac injection of 50 ml PBS. Spleen, popliteal lymph nodes, and CNS were removed and either used immediately or stored at 7808C. Reagents ConA was purchased from Sigma Chemical Co. (St. Louis, MO, USA) and LPS from Difco Laboratories (Detroit, MI, USA). The following antibodies were used for immuno¯uorescence staining in FACS analysis: PE-conjugated anti-CD4 and anti-CD8 mAbs purchased from Caltag (South San Francisco, CA, USA); FITC-conjugated anti-CD4 and antiCD11b (anti-Mac-1) mAbs. The following primary antibodies were used for immuno¯uorescence staining of sections: unlabelled rat anti-CD4 and anti-CD8 mAbs (Caltag); biotinylated mouse antiMHC class I (H-2Kd) mAb (SF1-1.1; Pharmingen); FITC-conjugated rabbit anti-rabies nucleocapsid (NC) Ab (Sano® Diagnostics, Marnes la Coquette, France); mouse mAb anti-calbindin-D (Sigma) to visualise Purkinje cells, mouse mAb anti-neuronal nuclei (NeuN; Chemicon, Temecula, CA, USA) to visualise other neurons; goat polyclonal IgG speci®c for the p20 subunit of CPP32 activated by cleavage but not procaspase-3 (Santa Cruz Biotechnology, Santa Cruz, CA, USA) to detect activated caspase-3 in apoptotic cells. Staining with primary or secondary biotinylated antibody was followed by FITC-streptavidin (Caltag) or Cy3-conjugated strep- Immunopathology of nonfatal rabies A Galelli et al 369 tavidin (Jackson Immunoresearch, Westgrove, PA, USA). The following af®nity pure secondary antibodies were used: FITC-conjugated goat-anti rat FITC (Caltag); rhodamine (TRITC)-conjugated goat anti-mouse IgG1 (H+L). RT ± PCR Total cellular RNA was extracted from popliteal lymph nodes or CNS mononuclear cells, using RNA BTM (Bioprobe, Montreuil, France), according to the manufacturer's instructions. Integrity of RNA was tested by electrophoresis. Two mg of total RNA was subjected to ®rst-strand cDNA synthesis in 20 ml reaction volume containing 10 mM Tris, 50 mM KCl, 1.5 mM MgCL2, 1 mM of each dNTPs, 20 mg oligo(dT16), 20 U AMV reverse transcriptase (Boehringer Mannheim, Mannheim, Germany) and 50 U RNase inhibitor, for 40 min at 428C. For detection of genomic RNA rabies virus, reverse transcription was performed using 1 mM rabies virus N proteinspeci®c sense primer as described (Shankar et al, 1991), instead of oligo(dT16). The resulting cDNA was diluted 1 : 5 with distilled water and used in PCR. PCR ampli®cations were performed in 50 ml reaction volume containing 10 mM Tris, 50 mM KCl, 5 mM MgCl2, 200 mM of each dNTP, 1 mM of each speci®c primer, and 2.5 U Taq polymerase (Perkin-Elmer, Cetus Corp., Norwalk, CT, USA). An initial denaturation step was performed at 948C for 3 min. The cycling program consisted of a denaturation step at 948C for 45 s, primer annealing for 1 min and elongation at 728C for 1 min. In the last cycle, the elongation step was prolonged to 10 min. PCR reactions were performed in a GeneAmp1 PCR System 9600 Thermal Cycler (Perkin-Elmer). Primers for RT ± PCR Oligonucleotide sequence of primers and probes were synthesised by Genset (Genset SA, Paris, France) as follows: IFN-g (GenBank accession number K00083) 5'-AGC TCT GAG ACA ATG AAC GC-3' (left), 5'-GGA CAA TCT CTT CCC CAC CC-3' (right), 5'-GTG GAA GGA CTC ATG ACC ACA GTC CATG CC-3' (probe); TNF-a (GenBank accession number M13049), 5'-GTA TGA GAT AGC AAA TCG GC-3' (left), 5'-CTG AAC TTC GGG GTG ATC GG-3' (right), 5'-AGG AGG GCG TTG GCG CGC TG3' (probe); MIP-1a (GenBank accession number X53372), 5'-AAG CAG CAG CGA GTA CCA GT-3' (left), 5'-CTC AAG CCC CTG CTC TAC AC-3' (right), 5'-AGG TGT CAT TTT CCT GAC TAA GAG AAA C3' (probe); MIP-1b (GenBank accession number X62502), 5'-GCT CTG TGC AAA CCT AAC CC-3' (left), 5'-AAG AAG AGG GGC AGG AAA TC-3' (right), 5'-AGA AGC TTT GTG ATG GAT TAC TAT GAG AC-3' (probe); IP-10 (GenBank accession number L07417), 5'-CCT ATC CTG CCC ACG TGT TGA G-3' (left), 5'-CGC ACC TCC ACA TAG CTT ACA G-3' (right), 5'-ATG GAC AGC AGA GAG CCT GT-3' (probe); RANTES (GenBank accession num- ber U02298), 5'-CCC TCA CCA TCA TCC CTC ACT3' (left), 5'-ATT TCT TGG GTT TGC TGT GC-3' (right), 5'-AAG AAT ACA TCA ACT ATT TGG AGA T-3' (probe); GAPDH (GenBank accession number M32599) 5'-ACC ACC ATG GAG AAG GCT GG-3' (left), 5'-CTC AGT GTA GCC CAG GAT GC-3' (right), 5'-GTG GAA GGA CTC ATG ACC ACA GTC CAT GCC-3' (probe). Forward and reverse primers were selected from different exons. Rabies N protein (GenBank accession number AF033905), 5'-GGA ATT CTC CGG AAG ACT GGA CCA GCT ATG G-3' (sense primer), 5'-AGA ATT CCC ACT CAA GCC TAG TGA ACG G-3' (antisense primer), 5'-GGC CGG AAC CTA TGA CAT GTT TTT CTC CCG-3' (probe) (Shankar et al, 1991). Analysis and semi quantitation of PCR products In pilot experiments, designed to control the speci®city of ampli®cation, PCR products were run on an agarose gel stained with ethidium bromide, gel-puri®ed and sequenced with the same primers, using the Dye Terminator sequencing kit and automatic sequencing (ABI Prims 377 DNA Sequencer; Perkin-Elmer). PCR products were quanti®ed by ELISA after hybridisation in solution using the PCR ELISA Boehringer kit (Boehringer Mannheim). Identical results were obtained when PCR products were quanti®ed by densitometry analysis of ethidium bromide-stained agarose gels, using a video system and ImageQuaNT software (Molecular Dynamics, Sunnyvale, CA, USA). Therefore, this method was used for semi-quantitative determination of gene expression. The amount of PCR products accumulated was measured during the exponential phase of target ampli®cation, when none of the reaction components is limiting. The exponential phase was determined for each set of primers, with samples containing a high starting copy number. Ampli®cation of an endogenous control, the housekeeping gene glyceraldehyde-3phosphate dehydrogenase (GAPDH) was performed to standardise the amount of cDNA added to a reaction. The relative quantitation of genomic rabies RNA or the relative expression of cytokinespeci®c mRNA was determined by dividing the value obtained for the target RNA by that obtained for GAPDH on the same sample. Targets and endogenous controls were ampli®ed in separate tubes. The PCR to detect rabies virus RNA and GAPDH transcripts was performed with 30 cycles of ampli®cation. Detection of mRNA speci®c for cytokines and chemokines was performed using 35 cycles of ampli®cation. The presence of mRNA speci®c for IL-2, IL-4, IL-10 was also tested after 40 cycles of ampli®cation. Immuno¯uorescence staining of tissue sections Dissected brains and spinal cords were embedded in tissue-Tek O.C.T. compound (Miles Elkhart, IN, USA), frozen on powdered dry ice and stored at Journal of NeuroVirology Immunopathology of nonfatal rabies A Galelli et al 370 7808C. Parasagittal brain and longitudinal spinal cord sections (8 mm) were cut on a cryostat, airdried, and stored at 7808C until preparation for ¯uorescence staining. Sections were ®xed in 4% paraformaldehyde for 10 min, washed three times in PBS, permeabilised in PBS-0.25% Triton X-100 for 10 min, and incubated for 30 min with blocking buffer (5% goat serum in PBS) to minimise background staining. Sections were then incubated with predetermined optimal concentrations of primary antibodies in blocking solution for 2 h at room temperature or overnight at 48C. After washing, sections were incubated for 30 min with either FITC-streptavidin, FITC-, or TRITC-labelled appropriate secondary antibodies and then mounted with glycerol containing 1,4-diazobicyclo-(2,2,2) octane (Sigma). Activated caspase-3 was detected in a three step method including staining with an unlabelled primary antibody, then with an appropriate biotinylated secondary antibody and lastly with ¯uorochrome-conjugated streptavidin. Controls omitting one or the other primary antibody revealed no staining and no cross reactivity between the stains. Reactivity was visualised with a light microscope equipped with ¯uorescence (Leica Optovar). Appropriate ®lters for each ¯uorochrome and a combined ®lter for ¯uorescein and rhodamine were used. Images were fused in one document using the Macintosh Adobe Photoshop 3 program. Quantitation of rabies virus infected neurons Six similar parasagittal brain sections and longitudinal spinal cord sections were selected from asymptomatic (n=2) and paralysed mice (n=2) to estimate the number of infected neurons. Sections were double stained for either rabies virus NC and calbindin (cerebellum) or rabies virus NC and NeuN (spinal cord). The total number of infected Purkinje cells per section was determined. In spinal cord sections, neurons were counted in ®ve randomly selected ®elds per section and a mean percentage of infected cells was calculated. Detection of apoptosis Apoptosis was detected either by staining with an antibody speci®c for the activated form of caspase-3, the central effector of apoptosis, or by enzymatic labelling of DNA strand breaks with the TUNEL technique (terminal deoxynucleotidyl transferase-mediated dUTP nick end labelling (Gavrieli et al, 1992). Tissue sections were ®xed and incubated with blocking solution containing Triton X-100, as described above. Sections were then incubated for 1 h at 378C with labelling mix containing 0.3 U/ml terminal deoxynucleotidyl transferase (TdT) and 20 mM biot-16-dUTP in supplied buffer (Boehringer Mannheim). Labelled nuclei were detected by Cy3-streptavidin. Negative control was performed by omitting TdT. Journal of NeuroVirology Isolation of mononuclear cells from the CNS Brain and spinal cord tissues were dissociated between frosted glass slides. After complete dissociation by vigorous pipetting, the brain suspension was centrifuged at 2006g for 10 min to remove cell debris and lipids. Mononuclear cells were isolated using a discontinuous Percoll gradient as previously described (Clatch et al, 1990). Brie¯y, the cell pellet was re-suspended in 5 ml of 70% Percoll (Pharmacia, Uppsala, Sweden) in RPMI 1640. This suspension was then overlaid with equal volumes of 37% and 30% Percoll and the gradient was centrifuged at 5006g for 15 min at 208C. The cell layer between the 70% : 37% Percoll interface was recovered and brain mononuclear cells were washed twice in medium. Fluorescence staining for FACS analysis Cells (106 in 50 ml) were stained with predetermined optimal concentrations of ¯uorescent reagents in FACS buffer (PBS supplemented with 2% BSA and 0.02% sodium azide) for 15 min at 48C. Brain T cells were characterised by double staining with either anti-CD4 or anti-CD8 and anti-Mac-1. Phenotypic analysis was performed with an Excalibur ¯ow cytometer equipped with the CellQuest software (Becton Dickinson, Mountain View, CA, USA). Dead cells and debris were gated out, prior to data acquisition, using side-scatter/forward-scatter scatterplots. Ten thousand events were acquired in each run. Detection of anti-rabies virus antibodies Mouse sera were tested for the presence of rabiesspeci®c antibodies by ELISA as previously described (Lafon et al, 1990). Plates were coated with 0.1 mg/well of UV-inactivated ERA strain rabies virus. Serum samples were tested in twofold serial dilutions. The negative control consisted of pooled normal mouse sera. Individual titres were expressed as the log10 of the highest dilution giving a value twice as high as that observed with negative control. Rabies virus neutralising antibodies were quanti®ed by the rapid ¯uorescent focus inhibition test (RFFIT), using CVS rabies virus and BSR cells (Montano-Hirose et al, 1993). Neutralisation titres were expressed as IU/ml using a standard rabies neutralising human IgG. Proliferation assays Triplicate spleen cell samples (106/ml) were cultured for 3 days in 200 ml of RPMI 1640 medium (Gibco Laboratories, Grand Island, NY, USA) supplemented with 5% FCS, 2 mM L-glutamine, 561075 M 2-ME, 1 mM sodium pyruvate, 10 mM HEPES and antibiotics. Cultures were stimulated with the lowest concentration of mitogen giving an optimal response: Con A (1 mg/ml) or LPS (10 mg/ ml). Cells were pulsed with 0.25 mCi/well of Immunopathology of nonfatal rabies A Galelli et al [3H]TdR (DuPont de Nemours, NEN Products, Les Ulis, France) and harvested 4 h later. Radioactivity was determined in a liquid scintillation beta counter (Micro-b-6 detector Plus, Wallac, Ltd, Turku, Finland). 371 Acknowledgements This work was supported by an institutional grant from the Pasteur Institute. The authors gratefully acknowledge Dr S Nicholson for her critical review of this manuscript. Statistics Standard deviation (s.d.) was used as an estimate of variance and means were compared by using Student's t-test. References Albert ML, Sauter B, Bhardwaj N (1998). Dendritic cells acquire antigen from apoptotic cells and induce class I restricted CTLs. Nature 392: 86 ± 89. Baer GM, Cleary WF, Diaz AM, Perl DF (1977). Characteristics of 11 rabies virus isolates in mice: titers and relative invasiveness of virus incubation period of infection, and survival of mice with sequelae. J Infect Dis 136: 336 ± 345. Bellone M, Lezzi G, Rovere P, Galati G, Ronchetti A, Protti MP, Davoust J, Rugarli C, Manfredi AA (1997). Processing of engulfed apoptotic bodies yields T cell epitopes. J Immunol 159: 5391 ± 5399. Bennett SRM, Carbone FR, Karamalis F, Miller JFAP, Heath WR (1997). Induction of a CD8+ cytotoxic T lymphocyte response by cross-priming requires cognate CD4+ T cell help. J Exp Med 186: 65 ± 70. Bevan MJ (1976). Cross-priming for a secondary cytotoxic response to minor H antigens with H-2 congenic cells which do not cross-react in the cytotoxic assay. J Exp Med 143: 1283 ± 1288. Biddison WE, Kubota R, Kawanishi T, Taub D, Cruikshank WW, Center DM, Connor EW, Utz U, Jacobson S (1997). Human T cell leukemia virus type I (HTLV1)-speci®c CD8+ CTL clones from patients with HTLV1-associated neurologic disease secrete proin¯ammatory cytokines, chemokines, and matrix metalloproteinase. J Immunol 159: 2018 ± 2025. Chopra JS, Banerjee AK, Murthy JM, Pal SR (1980). Paralytic rabies: a clinicopathological study. Brain 103: 789 ± 802. Clatch RJ, Miller SD, Metzner R, Dal Canto MC, Lipton HL (1990). Monocytes/macrophages isolated from the mouse central nervous system contain infectious Theiler's murine encephalomyelitis virus (TMEV). Virology 176: 244 ± 254. Dietzschold B, Kao M, Zheng YM, Chen ZY, Maul G, Fu ZF, Rupprecht CE, Koprowski H (1992). Delineation of putative mechanisms involved in antibody-mediated clearance of rabies virus from the central nervous system. Proc Natl Acad Sci USA 89: 7252 ± 7256. Duguid J, Trzepacz C (1993). Major histocompatibility complex genes have an increased brain expression after scrapie infection. Proc Natl Acad Sci USA 90: 114 ± 117. Gamaleia N (1887). Etude sur la rage paralytique chez l'homme. Ann Inst Pasteur (Paris) 1: 63 ± 83. Gavrieli Y, Sherman Y, Ben-Sasson SA (1992). Identi®cation of programmed cell death in situ via speci®c labeling of nuclear DNA fragmentation. J Cell Biol 119: 493 ± 501. Glabinski AR, Ransohoff RM (1999). Chemokines and chemokine receptors in CNS pathology. J Neurovirol 5: 3 ± 12. Hirai K, Kawano H, Mifune K, Fujii H, Nishizono A, Shichijo A, Mannen K (1992). Suppression of CellMediated Immunity by Street Rabies Virus infection. Micriobiol Immunol 36: 1277 ± 1290. Hooper DC, Morimoto K, Bette M, Weihe E, Koprowski I, Dietzschold B (1998). Collaboration of antibody and in¯ammation in clearance of Rabies virus from the central nervous system. J Virol 72: 3711 ± 3719. Iwasaki Y, Gehrard W, Clark HF (1977). Role of host immune response in the development of either encephalitic or paralytic disease after experimental rabies infection in mice. Infect Immun 18: 220 ± 225. Jackson AC, Rossiter JP (1997). Apoptosis plays an important role in experimental rabies virus infection. J Virol 71: 5603 ± 5607. Joly E, Mucke L, Oldstone MBA (1991). Viral persistence in neurons explained by lack of major histocompatibility class I expression. Science 253: 1283 ± 1285. Jondal MR, Schirmbeck R, Reimann J (1996). MHC class I-restricted CTL responses to exogenous antigens. Immunity 5: 295 ± 302. Lafon M, Edelman L, Bouvet JP, Lafage M, Montchatre E (1990). Human monoclonal antibodies speci®c for the rabies virus glycoprotein and N protein. J Gen Virol 71: 1689 ± 1696. Lafon M, Galelli A (1996). Superantigen related to virus. Springer Semin Immunopathol 17: 307 ± 318. Ludowyk PA, Willenborg DO, Parish CR (1992). Selective localisatioan of neuro-speci®c T lymphocytes in the central nervous system. J Neuroimmunol 37: 237 ± 250. Massa PT, Ozato K, McFarlin DE (1993). Cell-type speci®c regulation of major histocompatibility complex (MHC) class I gene expression in astrocytes, oligodendrocytes, and neurons. Glia 8: 201 ± 207. Merrill JE, Benveniste EN (1996). Cytokines in in¯ammatory brain lesions: helpful and harmful. Trends Neurosci 19: 331 ± 338. Merrill JE, Ignarro LJ, Sherman MP, Melinek J, Lane TE (1993). Microglial cell cytotoxicity of oligodendrocytes is mediated through nitric oxyde. J Immunol 151: 2132 ± 2141. Momburg F, Koch N, Moller P, Moldenhauer G, Butcher GW, Hammerling GJ (1986). Differential expression of Ia and Ia-associated invariant chain in mouse tissues after in vivo treatment with IFN-gamma. J Immunol 136: 940 ± 948. Journal of NeuroVirology Immunopathology of nonfatal rabies A Galelli et al 372 Montano-Hirose JA, Lafage M, Weber P, Badrane H, Tordo N, Lafon M (1993). Protective activity of a murine monoclonal antibody against European bat lyssavirus 1 (EBL1) infection in mice. Vaccine 11: 1259 ± 1266. Morimoto K, Hooper DG, Spitsin S, Koprowski H, Dietzschold B (1999). Pathogenicity of different rabies virus variants inversely correlates with apoptosis and rabies virus glycoprotein expression in infected primary neuron cultures. J Virol 73: 510 ± 518. Murphy FA, Harrison AK, Winn WC, Bauer SP (1973). Comparative pathogenesis of rabies and rabies-like viruses: infection of the central nervous system and centrifugal spread of virus to peripheral tissues. Lab Invest 29: 1 ± 16. Neumann H, Schmidt H, Cavalie A, Jenne D, Wekerle H (1997). Major histocompatibility complex (MHC) class I gene expression in single neurons from the central nervous system: differential regulation by interferon (IFN)-g and tumor necrosis factor (TNF)-a. J Exp Med 185: 305 ± 316. Oldstone MBA, Blount P, Southern PJ, Lampert PW (1986). Cytoimmunotherapy for persistent virus infection reveals a unique clearance pattern from the central nervous system. Nature 321: 239 ± 243. Price DA, Klenerman P, Booth BL, Phillips RE, Sewell AK (1999). Cytotoxic T lymphocytes, chemokines, and antiviral immunity. Immunol Today 20: 212 ± 216. Ren Y, Savill J (1998). The importance of being eaten. Cell Death Differ 5: 563 ± 568. Ronchetti A, Rovere P, Iezzi G, Galati G, Heltai S, Protti MP, Garancini MP, Manfredi AA, Rugarli C, Bellone M (1999). Immunogenicity of apoptotic cells in vivo: Role of antigen load, antigen-presenting cells, and cytokines. J Immunol 163: 130 ± 136. Savill JS, Fadok V, Henson P, Haslett C (1993). Phagocyte recognition of cells undergoing apoptosis. Immunol Today 14: 131 ± 136. Selmaj K, Raine CS, Farooq M, Norton WT, Brosnan CF (1991). Cytokine cytotoxicity against oligodendrocytes. Apoptosis induced by lymphotoxin. J Immunol 147: 1522 ± 1529. Sethna MP, Lampson LA (1991). Immune modulation within the brain: recruitment of in¯ammatory cells and increased major histocompatibility antigen expression following intracerebral injection of interferon-g. J Neuroimmunol 34: 121 ± 132. Shankar V, Dietzschold B, Koprowski H (1991). Direct entry of Rabies virus into the central nervous system without prior local replication. J Virol 65: 2736 ± 2738. Smith JS (1981). Mouse model of abortive rabies infection of the central nervous system. Infect Immun 31: 397 ± 308. Journal of NeuroVirology Smith JS, McClelland CL, Reid FL, Baer GM (1982). Dual role of the immune response in street rabies virus infection of mice. Infect Immun 35: 213 ± 221. Stoll G, Jung S, Jander S, van der Meide P, Hartung H-P (1993). Tumor necrosis factor-a in immune-mediated demyelination and Wallerian degeneration of the rat peripheral nervous system. J Neuroimmunol 45: 175 ± 182. Sugamata M, Miyazawa M, Mori S, Spangrude GJ, Ewalt LC, Lodmell DL (1992). Paralysis of street rabies virusinfected mice is dependent on T lymphocytes. J Virol 66: 1252 ± 1260. Theerasurakarn S, Ubol S (1998). Apoptosis induction in brain during the ®xed strain of rabies virus infection correlates with onset and severity of illness. J Neurovirol 4: 407 ± 414. Thomas HE, Dutton R, Bartlett PF, Kay TWH (1997). Interferon regulatory factor 1 is induced by interferong equally in neurons and glial cells. J Neuroimmunol 78: 132 ± 137. Thornberry NA, Lazebnik Y (1998). Caspases: enemies within. Science 281: 1312 ± 1316. Vartanian T, Li Y, Zhao M, Stefansson K (1995). Interferon-induced oligodendrocyte cell death: Implications for the pathogenesis of multiple sclerosis. Mol Med 1: 732 ± 743. Wallach D (1997). Cell death induction by TNF: a matter of self control. Trends Biochem Sci 22: 107 ± 109. Weiland F, Cox JH, Meyer S, Dahme E, Reddehase MJ (1992). Rabies virus neuritic paralysis: Immunopathogenesis of nonfatal paralytic rabies. J Virol 66: 5096 ± 5099. Wekerle H (1993). T-cell autoimmunity in the central nervous system. Intervirology 35: 95 ± 100. Wiktor TJ, Doherty PC, Koprowski H (1977b). In vitro evidence of cell-mediated immunity after exposure of mice to both live and inactivated rabies virus. Proc Natl Acad Sci USA 74: 334 ± 338. Wiktor TJ, Doherty PC, Koprowsky H (1977a). Suppression of cell-mediated immunity by street rabies virus. J Exp Med 145: 1617 ± 1622. Wong GH, Bartlett PF, Clark-Lewis I, Battye F, Schrader JW (1984). Inducible expression of H-2 and Ia antigens on brain cells. Nature 310: 688 ± 691. Wunner WH (1987). Rabies Viruses-pathogenesis and immunity. In: The Rhabdoviruses. Wagner RR (ed). Plenum Press: New York, pp 361 ± 426. Zajicek JP, Wing M, Scolding NJ, Compston DA (1992). Interaction between oligodendrocytes and microglia: A major role for complement and tumor necrosis factor in oligodendrocyte adherence and killing. Brain 115: 1611 ± 1631.