Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
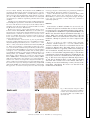
Am J Physiol Endocrinol Metab 297: E1414–E1419, 2009. First published October 13, 2009; doi:10.1152/ajpendo.00458.2009. Lactate distribution in culture medium of human myometrial biopsies incubated under different conditions Helena Åkerud,1 Gunnar Ronquist,2 and Eva Wiberg-Itzel3 Departments of 1Women’s and Children’s Health and 2Clinical Chemistry, Uppsala University, Uppsala; and 3Department of Obstetrics and Gynecology, Södersjukhuset, Karolinska Institutet, Stockholm, Sweden Submitted 24 July 2009; accepted in final form 12 October 2009 monocarboxylate transporter 1; monocarboxylate transporter 4; myometrial cells; uterus is an inhomogeneous muscle that consists of smooth muscle cells where the myocytes are arranged in bundles embedded in connective tissue (1, 7). This arrangement gives uterus elastic properties and facilitates the transmission of contractile forces generated by individual muscle cells (26). The way in which human uterine smooth muscle cells metabolize and meet the energy demands during labor is still obscure. Energy is produced by glycolysis, ending with the formation of ATP and pyruvate. Pyruvate is usually, under aerobic conditions, converted to acetyl-CoA and, in the presence of oxaloacetate, subjected to mitochondrial oxidation to carbon dioxide and water. Under anaerobic conditions, pyruvate is reduced to lactate by lactate dehydrogenase in the presence of NADH. Steingrimsdottir et al. (20) demonstrated THE MYOMETRIUM IN HUMAN UTERUS Address for reprint requests and other correspondence: H. Åkerud, Dept. of Women’s and Children’s Health, Uppsala Univ., 751 85 Uppsala, Sweden (e-mail: [email protected]). E1414 that the human uterus utilizes glucose as its main energy substrate at term pregnancy. It seems that smooth muscle cells in uterus are capable of producing lactate at a higher rate under aerobic conditions (20) compared with striated muscle cells. Lactate has been thought to cause muscle fatigue, but during the last decade evidence has been presented to support the view that lactate can be even used as a substrate in liver cells under aerobic conditions (11). Earlier, lactate was considered to be transported across the membrane only via passive diffusion (5), but subsequent publications revealed a carrier-mediated transport of lactate across membranes (18 –19). Expression of the lactate carrier monocarboxylate transporter 1 (MCT-1) (4) was shown in astrocytes and monocarboxylate transporter 4 (MCT-4) was likewise found in skeletal muscle cells (14). It has been proposed that the force of smooth muscle contractions during labor is reduced by hypoxia in uterus (21), and Quenby et al. (16) pointed out that myometrial lactic acidosis is associated with dysfunctional labors. Amniotic fluid contains high levels of lactate (8, 25), and in previous publications by us (24) a high lactate concentration in amniotic fluid during labor was reported to correlate strongly with labor dystocia. However, the cellular origin of the lactate contribution to amniotic fluid has not been elucidated. The specific aim of this study was to examine whether myometrial cells are involved in the production of lactate in amniotic fluid and whether there are quantitative differences in production and distribution of lactate in myometrial cells incubated under aerobic and anaerobic conditions. We also wanted to elucidate the involvement of specific membranebound lactate carriers in the distribution of lactate. MATERIALS AND METHODS The study was approved by the regional Ethics Committee of the Medical Faculty of Uppsala University, and informed consent was obtained from each patient included in the study. Sample collection. Myometrial biopsies from human uterus were obtained during elective caesarean section (CS) under spinal anesthesia. All patients were term pregnant, none was in labor, and none had been delivered by CS before. Immediately after delivery of the infant, a biopsy of size 1 cm3 was cut from the border of the CS incision. The samples were rinsed in cold physiological saline solution (9 mg/ml NaCl). A piece of the biopsy was fixed in 4% paraformaldehyde for ⬃24 h and stored in 70% ethanol for a few days until embedded in paraffin wax according to routine procedures. The remaining part of the biopsy (that not fixed in paraformaldehyde) was put in a covered petri dish. Dulbecco’s modified Eagle’s medium (DMEM) (21885; GIBCO, Invitrogen) was added, and the volume was corrected according to the weight of the biopsy. It should be pointed out that culture medium and incubation medium in this study were equivalent notions. Culture of biopsies. The biopsies were mainly cultured under aerobic conditions at 37°C (Forma scientific incubator) with 5% CO2 0193-1849/09 $8.00 Copyright © 2009 the American Physiological Society http://www.ajpendo.org Downloaded from http://ajpendo.physiology.org/ by 10.220.33.5 on August 3, 2017 Åkerud H, Ronquist G, Wiberg-Itzel E. Lactate distribution in culture medium of human myometrial biopsies incubated under different conditions. Am J Physiol Endocrinol Metab 297: E1414 –E1419, 2009. First published October 13, 2009; doi:10.1152/ajpendo.00458.2009.—It is generally believed that a relationship exists between muscle fatigue and intracellular accumulation of lactate. This reasoning is relevant to obstetrical issues. Myocytes in uterus work together during labor, and the contractions need to be strong and synchronized for a child to be delivered. At labor dystocia, the progress of labor becomes slow or arrested after a normal beginning. It has been described that, during labor dystocia, when the force of the contractions is low, the uterus is under hypoxia, and anaerobic conditions with high levels of lactate in amniotic fluid dominate. The purpose of this study was to examine whether myometrial cells are involved in the production of lactate in amniotic fluid and whether there are differences in production and distribution of lactate in cells incubated under aerobic and anaerobic conditions. We also wanted to elucidate the involvement of specific membrane-bound lactate carriers. Women undergoing elective caesarean section were included. Myometrial biopsies from uteri were collected and subjected to either immunohistochemistry to identify lactate carriers or in vitro experiments to analyze production of lactate. The presence of lactate carriers named monocarboxylate transporters 1 and 4 was verified. Myometrial cells produced lactate extracellularly, and the lactate carriers operated differently under anaerobic and aerobic conditions; while being mainly unidirectional under anaerobic conditions, they became bidirectional under aerobic conditions. Human myometrial cells produced and delivered lactate to the extracellular medium under both anaerobic and aerobic conditions. The delivery was mediated by lactate carriers. E1415 LACTATE DISTRIBUTION IN HUMAN MYOMETRIUM microscopy. Negative control staining was performed by omitting the primary antibody. Nonspecific staining was not detected. Statistics. All statistical analyses were performed by SPSS 15.0 for Windows software package. For comparisons of median values, a Mann-Whitney U-test was used for independent samples. All significance tests were two-tailed. P values ⱕ0.05 were considered as statistically significant. RESULTS Demonstration of MCT-1 and MCT-4 in myometrial cells. To address the question of whether there are lactate carriers in myometrial cells or not, immunohistochemistry using antibodies against MCT-1 and MCT-4 was performed. Both MCT-1 and MCT-4 were detected in biopsies from human uterus (Fig. 1, A and B). MCT-1 was present and localized mainly at the cell membrane. MCT-4 was found both in stroma and at the cell membrane, although its presence was lower compared with MCT-1. Recovery of lactate in culture medium under anaerobic conditions. Uterine biopsies (n ⫽ 8) were cultured under anaerobic conditions, and levels of extracellular lactate were analyzed at different times. The initial release of lactate in incubation medium was apparently unidirectional during the 24 h of incubation, meaning that lactate was transported from an intra- to an extracellular compartment (Fig. 2). The median concentration of lactate in the medium was 2.0 mmol/l after 4 h followed by an increase to 10.6 mmol/l after 24 h (Table 1 and Fig. 2). The release curve displayed a different shape between 24 and 48 h of incubation where a peak value was obtained after 48 h with a lactate level of 14.1 mmol/l. After that, a levelling off was observed indicating a reentry of lactate to the intracellular compartment. No lactate was detected in the medium before incubation. Fig. 1. Monocarboxylate transporter-1 (MCT 1) and monocarboxylate transporter 4 (MCT-4) in human uterus. Immunohistochemical stainings identifying MCT-1 (A) and MCT-4 (B) in myometrial cells. AJP-Endocrinol Metab • VOL 297 • DECEMBER 2009 • www.ajpendo.org Downloaded from http://ajpendo.physiology.org/ by 10.220.33.5 on August 3, 2017 and 95% relative humidity. The incubation media (DMEM) were repeatedly checked for levels of lactate (see Measurement of lactate). -Hydroxybutyrate (-OH butyrate, 1 mmol/l) (B5887; Sigma Aldrich), iodoacetate (0.5 mmol/l) (I9148; Sigma Aldrich), or pyruvate (10, 50, or 500 mol/l) (P5280; Sigma Aldrich) was added (final concentrations) to a number of experiments performed under aerobic conditions. Some biopsies were cultured under anaerobic conditions. The biopsies were initially put in separate boxes, and the boxes were flushed with 95% N2 and 5% CO2. Each box was then placed in the incubator with an atmosphere of 95% N2 and 5% CO2 at 37°C. The medium was checked for pH and levels of lactate at different times. Measurement of lactate. A device that determines lactate concentration in amniotic fluid (LMU061; ObsteCare) was used. The device measures lactate concentration in amniotic fluid between 0.5 and 25.0 mmol/l and has a coefficient of variation of 3% at a lactate concentration of 11 mmol/l. The lactate recognition system was based on lactate oxidase with amperometric detection of the enzymatically produced hydrogen peroxide. Immunohistochemistry. Serial sections (5 m) of paraformaldehyde-fixed, paraffin-embedded tissue were deparaffinized in xylene, rehydrated through graded ethanol (99.5% 1 ⫻ 3 min, 95% 2 ⫻ 3 min, 70% 2 ⫻ 3 min), and washed in deionized water followed by TBS, pH 7.4. Slides were incubated in a citrate buffer and heated in a microwave oven for 10 min at 650 W. Nonspecific binding was blocked by incubation with 5% BSA in TBS for 1 h. Sections were then incubated overnight at 4°C in a humidified chamber with an affinity-purified goat anti-MCT-1 antibody (sc-14916; Santa Cruz Biotechnolgy) or an affinity-purified goat anti-MCT-4 antibody (sc-14930; Santa Cruz Biotechnolgy). After incubation, the sections were washed in 0.01% Tween 20/TBS buffer. A biotinylated secondary antibody (VEBA9500; Vector Laboratories) was added for 1 h at room temperature. Sections were washed, and streptavidin-conjugated horseradish peroxidase complex (Vector Laboratories) was added. After being rinsed, the sections were exposed to chromogen solution using a liquid DAB substrate kit (DAKO) and counterstained with Mayer’s hematoxylin. The appearance of a brown reaction product was observed by light E1416 LACTATE DISTRIBUTION IN HUMAN MYOMETRIUM Recovery of lactate in culture medium under aerobic conditions. Biopsies (n ⫽ 12) from human uteri were cultured under aerobic conditions, and levels of lactate in media were analyzed. Aerobic incubation conditions produced a different type of release pattern of lactate compared with anaerobic conditions. The median concentration of lactate was 2.2 mmol/l after 4 h in the incubator (Table 1), and, as shown in Fig. 2, the unidirectional efflux of lactate was noted only for the first 4 h of incubation, after which a bidirectional pattern was evident. The lactate concentration increased to 6.4 mmol/l after 24 h, and the peak value was achieved after only ⬃36 h, followed by a decline back to almost zero after 72 h. No lactate was detected in the medium before incubation. As shown in Table 1, the median levels of lactate were significantly higher after 24 h (P ⬍ 0.01) and 48 h (P ⫽ 0.001) of incubation under anaerobic conditions than under aerobic conditions. The difference between anaerobic and aerobic conditions after 4 h was not significant. Effect of -OH butyrate and iodoacetate on recovery of lactate in culture medium under aerobic conditions. Incubation of the biopsies under aerobic conditions in the presence of 1 mmol/l of -OH butyrate, which has structural similarities with lactate, influenced the release pattern of lactate; an enhancement of extracellular lactate was noted compared with controls (Fig. 3A). A delayed peak value was again followed by a sharp decline, indicating a reentry of lactate to the intracellular compartment. The biopsies were, in another set of experiments, cultured in the presence of 0.5 mmol/l Fig. 3. Effects of -hydroxybutyrate (-OH butyrate) and iodoacetate. Median values of lactate in culture medium with -OH butyrate (A) or iodoacetate (B) added. iodoacetate, which is an inhibitor of glycolysis. A decisively inhibitory effect on lactate appearance was then observed under aerobic conditions (Fig. 3B). Effect of pyruvate on recovery of lactate in culture medium under aerobic conditions. Pyruvate, added at the very low concentration of 10 mol/l in the culture medium, resulted in a modest increase of lactate under aerobic conditions (Fig. 4). At moderately elevated concentrations of pyruvate (50 and 500 mol/l), a sharp increase of lactate was observed in the incubation medium (Fig. 4). Moreover, a postponed addition of pyruvate at the same concentration, 36 h after the start of Table 1. Levels of lactate in culture medium Time, h Lactate, mmol/l Aerobic Lactate, mmol/l Anaerobic 4 10 24* 48* 72 2.2 0.3–3.5 2.0 1.5–2.7 5.3 2.5–6.8 6.4 1.1–11.9 10.6 8.8–12.6 3.0 0.1–13.6 14.1 9.3–17.2 0.8 0.1–13.4 13.4 4.4–17.2 Values are presented as median levels ⫾ variation of the lactate concentration. Aerobic, biopsies cultured under aerobic conditions; anaerobic, biopsies cultured under anaerobic conditions. *P ⬍ 0.05. AJP-Endocrinol Metab • VOL 297 • DECEMBER 2009 • www.ajpendo.org Downloaded from http://ajpendo.physiology.org/ by 10.220.33.5 on August 3, 2017 Fig. 2. Levels of lactate. Median values of lactate at different time points during culture. F, culture under aerobic conditions; ■, culture under anaerobic conditions. *Release curve is unidirectionally outward. **Bidirectional fluxes (outward and inward). LACTATE DISTRIBUTION IN HUMAN MYOMETRIUM incubation, brought about an even clearer elevation of the lactate in the culture medium (Fig. 5). DISCUSSION This study supports the idea that myometrial cells are capable of producing lactate at an elevated level under both anaerobic and aerobic conditions. The lactate recovered in the extracellular medium of incubated uterine biopsies was most probably glycolytically derived, since iodoacetate at a low concentration was an efficient inhibitor of glycolysis, resulting in very low levels of lactate throughout the experiment. Lactate transport across plasma membranes is facilitated by MCTs, and the MCT family of transporters consists of several isoforms. These transporters or carriers allow lactate to move (in symport with a proton) from a high to low concentration (3, 4, 5, 14), in our case either from uterine smooth muscle to amniotic fluid (which means passage across two layers of placental membranes, the chorion and amnion), or from circulating blood plasma to various active or inactive smooth muscle cells or other tissue cells where it may be metabolized. In this context, it should be kept in mind that MCT-1 has been demonstrated not only in the plasma membrane but also in mitochondria, where it is thought to facilitate lactate uptake in mitochondria for oxidation (13). The MCT-1 and MCT-4 are found abundantly in skeletal striated muscle cells (2, 12) and, as we show here for the first time to the best of our knowledge, in uterine smooth muscle cells as well. Although bidirectional in function, MCT-4 is well positioned to assist the efflux of lactate produced in cells with high rates of glycolysis (13); uterine smooth muscle cells (similarly to other smooth muscle cells) and epithelial cells both have a high rate of glycolysis (17). Our present study on uterine smooth muscle cells confirmed a high rate of glycolysis together with a high production rate of lactate, regardless of oxygen presence. High concentrations of lactate in culture medium of uterine biopsies incubated for prolonged time intervals (days) were detected; this is in line with a previous investigation suggesting that human myometrial cells can survive for an extended period of time (several days) under in vitro conditions, regardless of oxygen availability for energy metabolism (15). The release pattern of lactate AJP-Endocrinol Metab • VOL under strictly anaerobic conditions in the present study revealed a mainly unidirectional flux of this metabolite from the myometrial cells to the extracellular medium during the first 24 h. This first phase of lactate delivery was followed by a second phase, culminating in the attainment of a peak value after 48 h of incubation. This second phase probably comprised a reentry phenomenon of lactate from the extracellular medium into the myometrial cells, indicatory of a switch from unidirectional to bidirectional fluxes of lactate. Although revealing an identical efflux pattern of lactate with anaerobic incubation conditions during the first 4 h of incubation, the aerobic incubation conditions displayed subsequently a different release pattern of lactate. This was most probably because of the bidirectional character of the fluxes when reentry of lactate was favored, therewith depressing the extracellular levels of lactate. Also, the time of the peak value occurred considerably earlier, as did the decline of extracellular lactate to a value of almost zero, after 72 h of incubation. It was apparently possible to manipulate the working carriers of myometrial cells in different directions. By adding -OH butyrate to the extracellular medium at a modest concentration (compared with the lactate concentration), the extracellular lactate concentrations were substantially increased concomitantly with a delayed time for attainment of the peak value. This retarding effect on the reentry phenomenon of lactate by -OH butyrate was most probably a competitively inhibitory effect on the lactate carrier due to structural similarity between lactate and -OH butyrate. This view is corroborated by a previous finding on another structural analog. Moving the hydroxyl group of butyrate from a - to a ␥-position (i.e., from the third to the fourth carbon atom in the carbon chain) means formation of the neurotransmitter or neuromodulator ␥-hydroxybutyrate (GHB). GHB is present in mammalian brain as a metabolite of ␥-aminobutyric acid (10). Recent studies have demonstrated that MCT-1 constitutes an important carrier for GHB transport across the proximal tubule cells (22). Accordingly, -OH butyrate might well have been working as a competitive inhibitor in this context. It should be kept in mind that we only used one concentration of -OH butyrate. By increasing the concentration of this molecule in the extracel- Fig. 5. Effect of a second addition of pyruvate. Median values of lactate in culture medium after a postponed addition of pyruvate. Arrow indicates postponed addition of pyruvate. 297 • DECEMBER 2009 • www.ajpendo.org Downloaded from http://ajpendo.physiology.org/ by 10.220.33.5 on August 3, 2017 Fig. 4. Effect of pyruvate. Median values of lactate in culture medium with different concentrations of pyruvate added. E1417 E1418 LACTATE DISTRIBUTION IN HUMAN MYOMETRIUM AJP-Endocrinol Metab • VOL come from the myometrial cells since it is believed only to be transported through passive diffusion depending on a pH gradient (5). It is also known that the concentration of lactate is four to six times higher in amniotic fluid compared with fetal and maternal blood, and passive diffusion under these conditions would not be possible. Our results support the theory that lactate produced in human myometrial cells can be transported by carriers out into the surrounding compartments. This indicates, furthermore, that the lactate found in amniotic fluid at dystosic labor is certainly derived from the myometrial cells in the uterus. One limitation with our study is that it is an in vitro study; however, the fact that we used tissue from the human uterus means that the results nevertheless might be applicable to human physiology. The specific relevance of the lactate carriers in pregnancy and labor would be interesting to study further. Animal studies with pregnant MCT-1 or MCT-4 knockout mice would help us to better understand the mechanisms behind transportation of lactate during pregnancy and labor. Furthermore, additional clinically oriented studies are planned, where the relevance of MCT-1 and MCT-4 at different obstetrical issues will be elucidated. In conclusion, we show that human myometrial cells produce lactate under both anaerobic and aerobic conditions. Lactate carriers are operative in human uterus, and the presence of pyruvate in culture medium leads to increased levels of lactate supporting a carrier-mediated transport of lactate. ACKNOWLEDGMENTS We thank associate professor Matts Olovsson for valuable discussions. GRANTS This project was financed by grants from the Swedish Society of Medicine and from the Uppsala-Örebro Regional Research Council. DISCLOSURES E. Wiberg-Itzel has received shares in a company aiming to use other obstetrical research results for commercial means. No product is yet on the market. The rest of the authors report no conflicts of interest. The authors alone are responsible for the content and writing of the paper. REFERENCES 1. Andersson KE, Forman A, Ulmsten U. Pharmacology of labor. Clin Obstet Gynecol 26: 56 –77, 1983. 2. Benton CR, Campbell SE, Tonouchi M, Hatta H, Bonen A. Monocarboxylate transporters in subsarcolemmal and intermyofibrillar mitochondria. Biochem Biophys Res Commun 323: 249 –253, 2004. 3. Bergman BC, Wolfel EE, Butterfield GE, Lopaschuk GD, Casazza GA, Horning MA, Brooks GA. Active muscle and whole body lactate kinetics after endurance training in men. J Appl Physiol 87: 1684 –1696, 1999. 4. Broer S, Rahman B, Pellegri G, Pellerin L, Martin JL, Verleysdonk S, Hamprecht B, Magistretti PJ. Comparison of lactate transport in astroglial cells and monocarboxylate transporter 1 (MCT 1) expressing Xenopus laevis oocytes. Expression of two different monocarboxylate transporters in astroglial cells and neurons. J Biol Chem 272: 30096 –30102, 1997. 5. Brooks GA. Lactate shuttles in nature. Biochem Soc Trans 30: 258 –264, 2002. 6. Christensen HN. A transport system serving for mono- and diamino acids. Proc Natl Acad Sci USA 51: 337–344, 1964. 7. Conrad JT, Ueland K. Physical characteristics of the cervix. Clin Obstet Gynecol 26: 27–36, 1983. 297 • DECEMBER 2009 • www.ajpendo.org Downloaded from http://ajpendo.physiology.org/ by 10.220.33.5 on August 3, 2017 lular medium, we might well have been able to increase the extracellular concentration of lactate under these aerobic incubation conditions to a level comparable to those achieved under strictly anaerobic incubation conditions; it might also have been possible to eliminate the escape of extracellular lactate into myometrial cells. The presence of micromolar concentrations of pyruvate in the extracellular medium under aerobic conditions resulted in an increase of the extracellular lactate content (mM concentrations) over and above the level that was obtained under strictly anaerobic conditions. The hydroxyl group of lactate has been replaced by oxygen in a carbonyl group in pyruvate. Hence, a structural dissimilarity now exists. This obvious effect by small amounts of pyruvate on the extracellular lactate levels is not easily understood. Because phosphofructokinase is an allosterically regulated enzyme in glycolysis, one could argue that pyruvate might be influential on the enzyme in stimulating glycolysis and therewith lactate production. This is, however, not very probable. Known positive effectors of this allosterically regulated enzyme are free ADP, free AMP, and free orthophosphate concentrations but not pyruvate, nor could pyruvate be an extra substrate in the redox equlibrium reaction between pyruvate and lactate, since pyruvate was added in micromolar concentrations and the outcome of lactate augmentation was in the millimolar concentration range. Theoretically, extracellular pyruvate might act as an inducer of the upregulation of the synthesis of the membrane-bound carriers responsible for outward flux of lactate, but this argument is weakened by the rapid switch from reentry of lactate to unidirectional efflux of lactate brought about by the postponed addition of pyruvate (Fig. 5). Hence, a direct impact of pyruvate on the carrier seems more probable. We hypothesize that the enhanced effect on lactate exodus by pyruvate might have been exerted via a so-called “trans-stimulatory” effect. This classical term was introduced by Christensen (6) and implies a participation in the exchange of certain amino acids transported by systems demonstrating substrate selectivity and asymmetry of operation (23); a recent study revealed that even molecules not directly involved in exchange reactions were able to exert a trans-stimulatory effect on exodus of amino acids (9). We suggest that the concept of trans-stimulation, originally valid only for amino acids, could now be extended to encompass short-chain monocarboxylic acids. It is generally believed that a relationship exists between muscle fatigue (and thereby performance) and lactate intracellular accumulation. More specifically, muscle tissues that have a low, rather than a high, lactate concentration at the same muscular intensity can withstand dysfunction for a longer period. This reasoning is relevant to obstetrical issues. The human uterus consists of myocytes arranged in bundles (1, 7). These myocytes work together during labor, and the contractions need to be strong and synchronized for a child to be delivered. At labor dystocia, the progress of labor becomes slow or arrested after a normal beginning. It has been described that, during labor dystocia, when the force of the contractions is low, the uterus is under hypoxia and anaerobic conditions dominate (16). We have shown that a high lactate concentration in amniotic fluid during labor strongly correlates to labor dystocia (24). It has been discussed in the literature whether lactate in amniotic fluid could be derived from the uterus. In some publications, it has been postulated that lactate cannot LACTATE DISTRIBUTION IN HUMAN MYOMETRIUM AJP-Endocrinol Metab • VOL 18. Roth DA, Brooks GA. Lactate and pyruvate transport is dominated by a pH gradient-sensitive carrier in rat skeletal muscle sarcolemmal vesicles. Arch Biochem Biophys 279: 386 –394, 1990. 19. Roth DA, Brooks GA. Lactate transport is mediated by a membranebound carrier in rat skeletal muscle sarcolemmal vesicles. Arch Biochem Biophys 279: 377–385, 1990. 20. Steingrimsdottir T, Ronquist G, Ulmsten U. Energy economy in the pregnant human uterus at term: studies on arteriovenous differences in metabolites of carbohydrate, fat and nucleotides. Eur J Obstet Gynecol Reprod Biol 51: 209 –215, 1993. 21. Taggart MJ, Wray S. Hypoxia and smooth muscle function: key regulatory events during metabolic stress. J Physiol 509: 315–325, 1998. 22. Wang Q, Lu Y, Morris ME. Monocarboxylate transporter (MCT) mediates the transport of gamma-hydroxybutyrate in human kidney HK-2 cells. Pharm Res 24: 1067–1078, 2007. 23. White MF, Gazzola GC, Christensen HN. Cationic amino acid transport into cultured animal cells. I. Influx into cultured human fibroblasts. J Biol Chem 257: 4443– 4449, 1982. 24. Wiberg-Itzel E, Pettersson H, Cnattingius S, Nordstrom L. Association between lactate concentration in amniotic fluid and dysfunctional labor. Acta Obstet Gynecol Scand 87: 924 –928, 2008. 25. Wiberg-Itzel E, Pettersson H, Cnattingius S, Nordstrom L. Association between lactate in vaginal fluid and time to spontaneous onset of labour for women with suspected prelabour rupture of the membranes. Bjog 113: 1426 –1430, 2006. 26. Young RC. Myocytes, myometrium, and uterine contractions. Ann NY Acad Sci 1101: 72– 84, 2007. 297 • DECEMBER 2009 • www.ajpendo.org Downloaded from http://ajpendo.physiology.org/ by 10.220.33.5 on August 3, 2017 8. Fadel HE, Northrop G, Misenhimer HR, Harp RJ. Acid-base determinations in amniotic fluid and blood of normal late pregnancy. Obstet Gynecol 53: 99 –104, 1979. 9. Falck B, Bendsoe N, Ronquist G. Mediated exodus of L-dopa from human epidermal Langerhans cells. Amino Acids 26: 133–138, 2004. 10. Fishbein WN, Bessman SP. Gamma-hydroxybutyrate in mammalian brain. Reversible oxidation by lactic dehydrogenase. J Biol Chem 239: 357–361, 1964. 11. Gladden LB. Lactate metabolism: a new paradigm for the third millennium. J Physiol 558: 5–30, 2004. 12. Hashimoto T, Brooks GA. Mitochondrial lactate oxidation complex and an adaptive role for lactate production. Med Sci Sports Exerc 40: 486 – 494, 2008. 13. Hashimoto T, Masuda S, Taguchi S, Brooks GA. Immunohistochemical analysis of MCT1, MCT2 and MCT4 expression in rat plantaris muscle. J Physiol 567: 121–129, 2005. 14. Manning Fox JE, Meredith D, Halestrap AP. Characterisation of human monocarboxylate transporter 4 substantiates its role in lactic acid efflux from skeletal muscle. J Physiol 529: 285–293, 2000. 15. Olovsson M, Nordling M, Ulmsten U, Lindblom B, Waldenstrom A, Ronquist G. Human uterine myocytes retain their energy charge with no gross alterations in morphology for at least 8 days when cultured under anaerobic conditions. Gynecol Obstet Invest 49: 165–169, 2000. 16. Quenby S, Pierce SJ, Brigham S, Wray S. Dysfunctional labor and myometrial lactic acidosis. Obstet Gynecol 103: 718 –723, 2004. 17. Ronquist G, Andersson A, Bendsoe N, Falck B. Human epidermal energy metabolism is functionally anaerobic. Exp Dermatol 12: 572–579, 2003. E1419









![fermentation[1].](http://s1.studyres.com/store/data/008290469_1-3a25eae6a4ca657233c4e21cf2e1a1bb-150x150.png)




