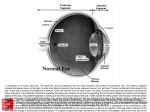
Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Research Signpost 37/661 (2), Fort P.O. Trivandrum-695 023 Kerala, India Advances in Ocular Drug Delivery, 2012: 1-31 ISBN: 978-81-308-0490-3 Editor: Ashim K. Mitra 1. Transport of drugs across the inner and outer blood-retinal barriers: Relevance of transporters in the retinal blood vessel endothelium and the retinal pigment epithelium Masanori Tachikawa1, Ken-ichi Hosoya1, Sylvia B. Smith2, Pamela M. Martin3 and Vadivel Ganapathy3 1 Department of Pharmaceutics, Graduate School of Medicine and Pharmaceutical Sciences University of Toyama, Toyama, Japan; 2Department of Cellular Biology and Anatomy, Medical College of Georgia, Augusta, Georgia, U. S. A.; 3Department of Biochemistry and Molecular Biology Medical College of Georgia, Augusta, Georgia, U. S. A. Introduction There is an urgent need for the development of optimal and efficient drug delivery systems for the treatment of retinal diseases because of the potential loss of vision if these diseases go untreated. Delivery of therapeutic drugs to the retina poses various hurdles. Topical application of drugs (eye drops) is ineffective in achieving therapeutically relevant concentrations of the drugs in the retina due to the long diffusional distance and counter-directional intraocular convection from the ciliary body to Schlemm’s canal [1]. Intravitreal delivery with implants and direct injections carries a high risk Correspondence/Reprint request: Dr. Masanori Tachikaw, Department of Pharmaceutics, Graduate School of Medicine and Pharmaceutical Sciences, University of Toyama, Toyama, Japan 2 Masanori Tachikawa et al. of deleterious side effects such as post-operative endophthalmitis, hemorrhage, and retinal detachment. It is widely believed that systemic administration is not an effective means to deliver drugs into the retina because of the blood-retinal barriers. The retina is considered as a privileged tissue protected by the inner and outer blood-retinal barriers from potentially harmful chemicals and agents that may be present in the systemic circulation. While this particular role of blood-retinal barriers is certainly beneficial to the retina, it also poses a significant problem to deliver therapeutic drugs to the retina via systemic administration. However, the blood-retinal barriers are not impermeable structures; essential nutrients to support the growth and viability of various cell types within the retina are transferred efficiently from systemic circulation via these barriers. Recent progress in blood-retinal barrier research has revealed that the specific cell types comprising these barriers express a wide variety of transporters essential for the blood-toretinal influx of these nutrients. There are some transporters however that contribute to the protective function of the blood-retinal barriers by mediating the retina-to-blood efflux of toxins and drugs, thus playing an active role in the removal of potentially harmful chemicals from the retina. It has become increasingly clear in recent years that even though the physiologic function of most of the transporters in mammalian cells is to facilitate transmembrane movement of biologically relevant compounds such as amino acids, monosaccharides, nucleosides, monocarboxylates, and vitamins, these transporters do recognize several xenobiotics and therapeutic agents that might bear structural resemblance to their physiological substrates. This feature can be exploited to our advantage to develop efficient strategies for optimal delivery of clinically relevant therapeutic drugs into the retina [2,3]. The blood-retinal barriers consist of retinal capillary endothelial cells (inner blood-retinal barrier) and retinal pigment epithelial (RPE) cells (outer bloodretinal barrier) (Fig. 1). Both these cell types form tight monolayers with complex tight junctions between adjacent cells within the monolayer. This prevents or decreases non-specific diffusion of chemicals across the monolayer. The inner blood-retinal barrier is responsible for nourishment of the inner two-thirds of the retina while the outer blood-retinal barrier is responsible for nourishment of the remaining one-third of the retina [4]. Thus, essential nutrients for photoreceptor cells are supplied through transfer across RPE from choroidal circulation whereas those for other neuronal cells (e.g., ganglion cells, bipolar cells, horizontal cells, amacrine cells) and Muller cells come mostly through transfer across the retinal capillary endothelial cells. Since these two monolayers have to perform vectorial transfer of nutrients in the blood-to-retina direction and also eliminate metabolic waste products in the retina-to-blood direction, the capillary endothelial cells of the Drug delivery to retina 3 Figure 1. Schematic representation of the retina and the inner and outer blood-retinal barriers. The blood-retinal barriers form complex tight junctions of retinal capillary endothelial cells (inner blood-retinal barrier) and retinal pigment epithelial cells (outer blood-retinal barrier). GC, ganglion cell; AC, amacrine cell; BC, bipolar cell; HC, horizontal cell; MC, Muller cell; RC, rod photoreceptor cell; CC, cone photoreceptor cell; RPE, retinal pigment epithelial cell. inner blood-retinal barrier and the RPE cells of the outer blood-retinal barrier are polarized. The plasma membrane of the endothelial cells consists of a luminal membrane that is in contact with blood and an abluminal membrane that faces the retina. Similarly, the plasma membrane of the RPE cells consists of a basolateral membrane that is in contact with the Bruch’s membrane apposed to choroidal blood and an apical membrane that faces the retina. The feasibility and success of drug delivery to the retina using transporters depend on several factors: (i) identity of the transporters that are expressed specifically in the inner and outer blood-retinal barriers, (ii) differential expression of the transporters in the two poles of the plasma membrane of the polarized cells that constitute these barriers, and (iii) substrate selectivity of the individual transporters, particularly differences in substrate specificity between the influx transporters and the efflux transporters. 4 Masanori Tachikawa et al. Influx transporters at the inner blood-retinal barrier The retinal capillary endothelial cells that constitute the inner bloodretinal barrier supply essential nutrients such as glucose, amino acids, vitamins, and nucleosides to the retina. The role of transporters in this process has been assessed by comparing the blood-to-retina permeability rates of these essential nutrients with that of mannitol, a marker of passive non-carrier-mediated diffusion. The nutrients have several-fold higher permeability rates than mannitol (Table 1), implying that specific transporters are involved in the blood-to-retina transfer of these compounds. The molecular identity of the transporters responsible for the transfer of individual nutrients has been established using a conditionally immortalized rat retinal capillary endothelial cell line (TR-iBRB cells) [5]. The influx transporters identified thus far in this cell line are listed in Fig. 2. Some of these transporters transport not only their physiologic substrates but also several xenobiotics and therapeutic drugs that structurally mimic the endogenous substrates, suggesting that these transporters may have potential as drug delivery systems for the treatment of retinal diseases. Table 1. Comparison of the permeability rates of the blood-to-retina influx transport. The influx permeability rate was determined by integration plot analysis after intravenous injection of radiolabeled compound. †Influx permeability rate of D-glucose (544 μL min-1 g retina-1) is calculated from the influx rate of Dglucose (6.8 μmol min-1 g retina-1)/normal D-glucose concentration in rat plasma (12.5 mmol/L [49]. * μL min-1 g eye-1. Drug delivery to retina 5 Figure 2. Efflux and influx transporters expressed in retinal capillary endothelial cells that constitute the inner blood-retinal barrier. DHA, dihydroascorbic acid; MTF, N5methyltetrahydrofolate. Transporter-mediated drug delivery across the inner bloodretinal barrier Amino acid mimetic drugs L-DOPA [levodopa, (-)-3-(3,4-dihydroxyphenyl)-L-alanine], the amino acid precursor of dopamine, is a widely used drug for treatment of Parkinson’s disease. Many patients with this disease have blurred vision or other visual disturbances, which are reflected in the reduced retinal dopamine concentration and delayed visual evoked potentials [6]. L-DOPA corrects these deficiencies [7], indicating that this amino acid is transported into the retina to serve as the precursor for dopamine synthesis to correct the deficiency of dopaminergic neurotransmission associated with Parkinson’s disease. L-DOPA is a neutral aromatic amino acid that has been shown to be a substrate for the Na+-independent amino acid transporter LAT1 [L (Leucine-Preferring) Amino acid Transporter 1; SLC7A5] [8]. This drug crosses the blood-brain barrier efficiently and enhances dopaminergic neurotransmission in the central nervous system in patients with Parkinson’s disease. It is known that LAT1, which is expressed in the brain capillary endothelial cells, is responsible for this transport process [9]. Since this transporter is also expressed in retinal capillary endothelial cells [10], it provides a potentially important route for the delivery of L-DOPA into the retina. Melphalan (phenylalanine mustard), an alkylaing agent used in the treatment of certain cancers, and gabapentin (an analog of -aminobutyrate), a drug used in the treatment of variety of neuropathic pain, are also transported via LAT1 [11]. Tomi et al [10] and Hosoya et al [12] have investigated the potential participation of LAT1 in the retinal delivery of various amino acid mustards as alkylating agents using TR-iBRB cell line as a model for the inner blood-retinal barrier. Since LAT1 is an obligatory 6 Masanori Tachikawa et al. amino acid exchanger, it is possible to determine directly whether or not a compound serves as a transportable substrate for LAT1 by monitoring the ability of the compound to induce the efflux of preloaded radiolabeled substrate [3H]phenylalanine in these cells. Surprisingly, melphalan failed to induce the efflux of [3H]phenylalanine in TR-iBRB cells, suggesting that this drug is not an effective transportable substrate for LAT1. With the same criterion, tyrosine-mustard, alanine-mustard, ornithine-mustard, and lysine-mustard were also shown to be poorly transportable substrates. In contrast, phenylglycinemustard was very effective in inducing the efflux of [3H]phenylalanine, suggesting that this particular amino acid mustard is an effective transportable substrate for LAT1. Even though L-DOPA and certain amino acid-mustards are recognized as transportable substrates for LAT1, other factors should be considered to evaluate the efficacy of this transporter to deliver these drugs into the retina in vivo. LAT1 possesses high affinity for its substrates; the Michaelis constant for leucine in TR-iBRB cells is ~15 M [10]. The normal plasma concentration of leucine is several-fold higher than this value (80-160 M). Furthermore, LAT1 accepts not only leucine but also other branched chain (isoleucine and valine) and aromatic (phenylalanine, tyrosine, and tryptophan) amino acids as substrates, which are present in the plasma in considerable quantities. Therefore, endogenous amino acid substrates are likely to decrease the efficacy of LAT1-mediated delivery of drugs into the retina. This effect may even be enhanced further under certain conditions such as high protein diet that increases the plasma levels of branched chain and aromatic amino acids. Nucleoside analogs Several nucleoside analogs are currently used in the treatment of viral infection and cancer. Examples of these analogs include 3’-azido-3’deoxythymidine (AZT), 2’, 3’-dideoxycytidine (ddC), 2’,3’-dideoxyinosine (ddI), cladribine, cytarabine, fludarabine, gemcitabine, and capecitabine. These drugs are transported into mammalian cells via nucleoside transporters, which handle purine and pyrimidine nucleosides as their physiologic substrates. The blood-to-retina transport of [3H]adenosine, a purine nucleoside, is carrier-mediated and is inhibited competitively by unlabeled adenosine and thymidine but not by cytidine [13]. Similar features are evident for adenosine transport in TR-iBRB cell line [13]. Molecular analysis of nucleoside transporters expressed in this cell line revealed that the Na+independent equilibrative nucleoside transporter ENT2 (SLC29A2) is expressed in these cells [13]. Adenosine plays an important role in retinal neurotransmission, blood flow, vascular development, and cellular response to ischemia. The delivery of adenosine into the retina across the inner blood- Drug delivery to retina 7 retinal barrier is therefore an important physiological process. The Michaelis constant for adenosine for transport via ENT2 is ~30 M, which is much higher than adenosine concentration in plasma (~0.1 M), indicating that ENT2 is not saturated with adenosine under physiologic conditions. Since the nucleoside analogs mentioned above are substrates for ENT2 [14,15], the transporter potentially plays a role in the delivery of such drugs into the retina. Antioxidants Retina is subjected to oxidative insult due to light exposure. Therefore, this tissue has an obligate need for antioxidants for protection against lightinduced damage to the cells. In addition, many retinal diseases have oxidative damage as an underlying pathological component. This includes diabetic retinopathy and age-related macular degeneration. Vitamin C, vitamin E, and glutathione are important antioxidants that may have potential in the treatment of retinal diseases caused by oxidative stress. Understanding the transport characteristics of these antioxidants at the inner blood-retinal barrier may assist in the design and development of suitable therapy with appropriate antioxidants for treatment of various retinal diseases. Vitamin C: Vitamin C, also known as ascorbic acid, exists in the plasma mostly in the oxidized form dehydroascorbic acid (DHA). The influx permeability rate of DHA is ~40fold greater than that of the reduced form ascorbic acid [16]. After entering the cells, DHA is reduced to generate ascorbic acid. The facilitative glucose transporter GLUT1 (SLC2A1) is responsible for cellular uptake of DHA, and this transporter is expressed on both the luminal and abluminal membranes of the endothelial cells that constitute the inner blood-retinal barrier [16,17]. The primary function of GLUT1 is to transport glucose from blood into retina. The Michaelis constant for GLUT1 for the transport of glucose is 5-8 mM, which is similar to the physiological plasma concentration of glucose (~5 mM). Therefore, GLUT1 is not completely saturated with its physiologic substrate in vivo. Transfer of DHA from blood into retina may occur via GLUT1 to a great extent even in the presence of normal physiologic levels of glucose in blood. The fact that GLUT1 is responsible for the delivery of glucose and DHA to the retina across the inner blood-retinal barrier is very relevant to diabetic retinopathy. Since plasma levels of glucose rise markedly in untreated diabetes, the transfer of DHA from blood to retina via GLUT1 may be impaired significantly in diabetes. The resultant deficiency of antioxidant machinery may contribute to the pathologic sequelae associated with diabetic retinopathy [18]. Vitamin E: Vitamin E has preventive and therapeutic effects in human retinopathies. Among the members of the vitamin E family, -tocopherol has the highest biologic activity, and is 8 Masanori Tachikawa et al. exclusively associated with high-density lipoprotein (HDL) in blood [19]. Uptake of HDL-associated -tocopherol into TR-iBRB cells is most likely mediated by scavenger receptor class B type I (SR-BI) [20]. Immunostaining of SR-BI is observed in rat retinal capillaries [20]. SR-BI at the inner bloodretinal barrier provides an efficient pathway for the supply of -tocopherol to retina from blood. Cystine: Glutathione is a major antioxidant in the retina. It is a tripeptide ( -Glu-Cys-Gly). This peptide is synthesized in mammalian cells using the component amino acids. Intracellular cysteine is low compared to the other two amino acids, and consequently cysteine represents the rate-limiting amino acid for glutathione synthesis. Cysteine is present in plasma predominantly in the oxidized form cystine. Uptake of cystine in mammalian cells occurs mostly via an amino acid exchanger known as cystine-glutamate exchanger. It is a Na+-independent process in which influx of cystine into cells is coupled to efflux of glutamate from the cells. The transport protein identified as xCT (SLC7A11) is responsible for the process. xCT is expressed in TR-iBRB cells [21]. The expression and activity of xCT in these cells is regulated in response to intracellular levels of glutathione. When the cellular levels of glutathione are depleted by treatment with diethylmaleate, the expression of xCT is upregulated to facilitate glutathione synthesis [21,22]. xCT at the inner blood-retinal barrier may thus be an important determinant of glutathione homeostasis in the retina. Miscellaneous protective compounds Compounds such as creatine, carnitine, acetylcarnitine, and taurine are biologically relevant to retina due to their role in energy homeostasis, fatty acid oxidation, and calcium signaling. Creatine: Creatine plays a vital role in the storage and transmission of phosphate-bound energy in retina. The Na+- and Cl--dependent creatine transporter (CRT, SLC6A8) plays a role in the influx of creatine into retina at the inner blood-retinal barrier. CRT is localized on both the luminal and abluminal membranes of rat retinal capillary endothelial cells [23]. CRT expressed on the luminal membrane would mediate creatine supply to retina; but the function of CRT on the abluminal membrane is not yet known. Creatine supplementation into retina is a potentially promising treatment for gyrate atrophy of the choroid and retina with hyperornithinemia. However, CRT at the inner blood-retinal barrier is almost saturated by plasma creatine (140-600 M in mice and rats), since the Michaelis constant for creatine uptake in TR-iBRB cells (~15 M) is much lower than these plasma concentrations [23]. The development of drugs which increase the density of CRT on the luminal membrane and/or CRT transport activity at the inner blood-retinal barrier is needed for rational creatine therapy of the gyrate Drug delivery to retina 9 atrophy. Carnitine and acetylcarnitine: Carnitine has multiple roles in mammalian cells, even though its primary and most recognized function is to promote fatty acid oxidation for energy production. Acetylcarnitine is effective in improving visual function in patients with early age-related macular degeneration. Uptake of carnitine and acetylcarnitine in TR-iBRB cells occurs via the Na+-dependent organic cation/carnitine transporter 2 (OCTN2, SLC22A5). OCTN2 is expressed in isolated rat retinal vascular endothelial cells [24]. The Michaelis constant for the transport of carnitine and acetylcarnitine via OCTN2 in TR-iBRB cells is ~30 M, a value similar to the physiological levels of these compounds in plasma (carnitine, ~50 M; acetylcarnitine, ~20 M) [24]. Exogenous administration of carnitine and acetylcarnitine would therefore be able to increase the retinal levels of these protective compounds through OCTN2mediated transfer across the inner blood-retinal barrier. Taurine: Taurine, the most abundant free amino acid in retina, functions as an osmolyte to regulate cellular volume under altered osmotic conditions. It also has antioxidant properties and ability to modulate calcium signaling. The Na+- and Cl--dependent taurine transporter (TAUT, SLC6A6) at the inner blood-retinal barrier mediates taurine transport from blood to the retina [25]. Since the Michaelis constant for taurine uptake by TR-iBRB cells (~20 M) is several-fold smaller than the plasma taurine concentration (100-300 M) in rats, the blood-to-retina taurine transport appears to be more than 80% saturated by the endogenous taurine under in vivo conditions [25]. We recently found that TAUT transports -aminobutyric acid (an inhibitory neurotransmitter) with a lower affinity than taurine; but the physiologic role of TAUT in the transport of this neurotransmitter in retina is not readily apparent [26]. Other transport systems Arginine Arginine is the precursor for generation of nitric oxide via nitric oxide synthases. Nitric oxide not only regulates vascular tone and blood flow but also is a critical component in a variety of cell signaling pathways. It is also an important determinant in the progression of retinal pathology in diseases such as diabetic retinopathy and glaucoma. Arginine uptake by TR-iBRB cells is most likely mediated by the Na+-independent cationic amino acid transporter 1 CAT1 (SLC7A1) [27]. CAT1 is expressed in retinal capillary endothelial cells [27]. Since nitric oxide synthesis depends on extracellular arginine, the function of CAT1 in the delivery of arginine into retina across the inner blood-retinal barrier may represent a rate-limiting step in nitric oxide production in retina. Since CAT1 also transports a variety of arginine- 10 Masanori Tachikawa et al. and lysine-based inhibitors of nitric oxide synthases, this transporter at the inner blood-retinal barrier can be exploited for delivery of such compounds into retina in the treatment of specific retinal diseases associated with overproduction of nitric oxide (e.g., inflammation). Lactic acid: Lactic acid is an important energy source for retinal neurons. Uptake of lactic acid in TRiBRB cells occurs via the H+-coupled monocarboxylate transporter 1 MCT1 (SLC16A1) [28]. MCT1 is localized in both the luminal and abluminal membranes of retinal capillary endothelial cells [29]. A number of monocarboxylates and monocarboxylic drugs inhibit lactic acid uptake in TR-iBRB cells. For example, salicylate and valproate competitively inhibit this process [28]. Therefore, MCT1 has potential for the delivery of monocarboxylic drugs into retina. Folates: Tetrahydrofolate plays an essential role as a cofactor for de novo synthesis of purines and pyrimidines, and also as a critical component in the metabolism of the sulfur-containing amino acids methionine and homocysteine. Deficiency of folate increases cellular levels of homocysteine, which has a wide variety of detrimental effects. Folate deficiency also causes visual dysfunction. Folate in the plasma of most mammals exists predominantly as the methyl derivative of the reduced folate, namely N5-methyltetrahydrofolate (MTF). Reduced folate carrier 1 (RFC1, SLC19A1) mediates MTF uptake by TR-iBRB cells [30]. This process is inhibited by methotrexate and formyltetrahydrofolate. RFC1 mRNA is expressed abundantly in freshly isolated rat retinal endothelial cells. Methotrexate is widely used as a chemotherapeutic agent in the treatment of cancer and also as an immunosuppressant in the treatment of rheumatoid arthritis. When present in circulation, this drug is most likely transported into retina via RFC1, consequently interfering with the entry of the physiologic substrate MTF into retina. Choline: Choline is an important cell membrane constituent in the form of phosphatidylcholine and sphingomyelin. It is also a precursor of the neurotransmitter acetylcholine. Choline uptake by TR-iBRB cells is Na+-independent and potentialdependent, indicating that a specific carrier exists at the inner blood-retinal barrier for the transfer of this important nutrient into retina [31]. The features of this uptake process are distinct from those of choline uptake mediated by other known organic cation transporters. Even though it is clear that a specific organic cation transporter is responsible for the transfer of choline into retina across the inner blood-retinal barrier, the molecular identity of the transporter remains to be established. In general, organic cation transporters exhibit broad substrate selectivity. Therefore, the transporter responsible for choline transfer across the inner blood-retinal barrier also has potential for delivery of specific organic cationic drugs into retina. Glycine: Glycine plays a pivotal role in neurotransmission and in the biosynthesis of creatine and Drug delivery to retina 11 glutathione in retina. It is also a co-agonist for the N-methyl-D-aspartate (NMDA) receptor. In vivo and in vitro studies have demonstrated that the Na+- and Cl--dependent glycine transporter 1 GlyT1 (SLC6A9) most likely mediates the blood-to-retina transport of glycine across the inner bloodretinal barrier [32]. Sarcosine, a methyl-substituted glycine, is also a substrate for GlyT1, and thus inhibits GlyT1-mediated cellular entry of glycine. Sarcosine also functions as a co-agonist for the NMDA receptor. Therefore, sarcosine has potential to modulate the activity of NMDA receptor by elevating extracellular levels of the co-agonist glycine through inhibition of glycine uptake via GlyT1 and also by acting directly as a co-agonist. This glycine analog is currently under investigation for treatment of schizophrenia as a means to activate the NMDA receptor. Since the NMDA receptor mediates the signaling of the neurotransmitter glutamate in retina as it does in central nervous system, the potential of GlyT1 as a delivery system for glycine and sarcosine into retina may be of clinical significance. Efflux transporters at the inner blood-retinal barrier Because the retina produces various metabolites and neurotoxic compounds, there must be mechanisms at the blood-retinal barrier to eliminate such compounds as a means to protect the retina from unwanted harmful effects. Effective efflux transport systems operate in other cell types, which are located strategically in different tissues to eliminate drugs and other xenobiotics from the body (e.g., intestine, liver, kidney, blood-brain barrier, placenta). While such efflux transport systems undoubtedly provide protection against potentially toxic xenobiotics, the transport systems also pose a significant problem for effective delivery of therapeutically active drugs if these drugs are recognized as substrates by the transport systems. Since the efflux transport systems actively remove their substrates from the cells, the presence of such transport systems at the blood-retinal barriers would effectively interfere with the delivery of therapeutic drugs into retina. Therefore, it is important to identify the efflux transport systems at these barriers and elucidate their substrate selectivity in terms of various drugs that are of potential use for the treatment of retinal diseases. Understanding the retina-to-blood efflux transport of drugs across the inner blood-retinal barrier will provide important information about the efficacy of drug delivery to the retina. The retina-to-blood efflux transport of anionic drugs from vitreous humor/retina across the inner blood-retinal barrier Many clinically important drugs including antibiotics, anti-tumor drugs, anti-HIV therapeutics, and anti-inflammatory agents are organic anions. The 12 Masanori Tachikawa et al. limited distribution of -lactam antibiotics in the vitreous humor/retina after systemic administration is problematic, resulting in reduced efficacy in the treatment of bacterial endophthalmitis [33]. 6-Mercaptopurine (6-MP) is frequently used as an anti-cancer drug in patients with childhood acute lymphoblastic leukemia. Relapse of childhood acute lymphoblastic leukemia involving eye is a rare but challenging problem. This is probably due to the restricted distribution of 6-MP in the eye [34]. One possible factor in the restricted drug distribution in the retina/eye is the retina-to-blood efflux transport of such anionic drugs across the blood-retinal barriers. In support of this notion, the transport of fluorescein, an organic anion, in the vitreous-toblood direction is more than 100-fold greater than that in the blood-tovitreous direction in humans [35]. Betz and Goldstein demonstrated that the uptake of p-aminohippuric acid (PAH), a well-known organic anion, by isolated retinal capillaries was greater than that of extracellular marker, sucrose, and that the uptake was inhibited by fluorescein [36]. We used microdialysis to carry out in vivo evaluation of vitreous/retina-toblood efflux transport in rats and to determine the efflux transport of organic anions across the blood-retinal barriers. 6-MP, PAH, and the -lactam antibiotic benzylpenicillin (PCG) were injected with D-mannitol, a bulk flow marker, into the vitreous humor of the rat eye, and a microdialysis probe was placed in the vitreous humor [37]. PCG, 6-MP, PAH and D-mannitol are biexponentially eliminated from the vitrous humor after vitreous bolus injection. The elimination rate constant of PCG, 6-MP, and PAH during the terminal phase was about 2-fold greater than that of D-mannitol. This efflux transport was reduced in the retina in the presence of probenecid, PAH, and PCG, relatively specific substrates of organic anion transporter (Oat) 3 (SLC22A8) [38]. Oat3 is localized on the abluminal membrane of retinal capillary endothelial cells [37]. Thus, Oat3 is involved in the uptake of PCG and 6-MP across the abluminal membrane of retinal capillary endothelial cells and contributes to the efflux transport of PCG and 6-MP from vitreous humor/retina into blood across the inner blood-retinal barrier. This process would be a critical factor in the restricted distribution of anionic drugs in retina. Some -lactam antibiotics are substrates for organic anion transporter polypeptide (oatp) 1a4 (Slco1a4; oatp2), which is present at the inner bloodretinal barrier in the rat [39,40]. We have recently reported that estradiol 17 glucuronide undergoes efflux transport from retina via oatp1a4 at the bloodretinal barriers. Taken together, oatp1a4 could also be involved in the clearance of anionic -lactam antibiotics at the inner blood-retinal barrier [41]. The retina-to-blood efflux transport of drugs consists of two steps, i.e., influx across the abluminal membrane from retina into retinal capillary Drug delivery to retina 13 endothelial cells and subsequent efflux across the luminal membrane from endothelial cells into blood. An ATP-dependent efflux pump for such anionic drugs is likely to exist on the luminal membrane to carry out the efflux process. ATP-binding cassette (ABC) transporter C 4 (multidrug resistance-associated protein, MRP4) is a promising candidate for the transport of these anionic drugs. Indeed, MRP4 transports several anionic drugs such as PAH, and also 6MP and -lactam antibiotics such as PCG [42]. We have demonstrated that MRP4 (ABCC4) mRNA is expressed abundantly in isolated mouse and rat retinal vascular endothelial cells [3,43]. It has been demonstrated that MRP4 is localized on the luminal membrane of human brain capillary endothelial cells [44], although the protein localization of MRP4 at inner blood-retinal barrier is yet to be demonstrated. These findings imply that MRP4 acts as an active efflux transporter for anionic drugs at the inner blood-retinal barrier and decreases the blood-to-retina transfer of -lactam antibiotics and 6-MP. In addition to MRP4, ABCC3 (MRP3) and ABCC6 (MRP6) mRNAs are also expressed abundantly in isolated mouse retinal vascular endothelial cells [43]. Other transporters involved in the retina-to-blood efflux transport at the inner blood-retinal barrier We have found that Oatp1c1 (Slco1c1/Oatp14) mRNA is highly expressed in isolated rat retinal endothelial cells [45]. Oatp1c1 transports estradiol 17 -glucuronide as is the case with Oatp1a4. On the other hand, Oatp1c1 does not have high affinity for digoxin [46], a specific substrate of Oatp1a4. This suggests that Oatp1c1 and Oatp1a4 play distinct roles in the retina-to-blood efflux transport in terms of the specificity of the drugs and xenobiotics that the two transporters handle. Further studies are needed to clarify the individual contribution of Oatp1c1 and Oatp1a4 to the efflux of specific anionic drugs across the inner blood-retinal barrier. Other members belonging to the ATP-dependent family of efflux transporters such as ABCA, ABCB, and ABCG could play a role in restricting the distribution of endobiotics and xenobiotics, including therapeutic agents, in retina. ABCB1 (P-glycoprotein, P-gp) is localized on the luminal membrane of retinal capillary endothelial cells [4]. The active efflux transport function of P-gp at the inner blood-retinal barrier could lower the blood-to-retina permeability of its substrates. For example, cyclosporin A, a substrate of P-gp, was not detected in the intraocular tissues of cyclosporine A-treated rabbits, although the blood level of cyclosporine A was within the therapeutic window [47]. TR-iBRB cells express P-gp, and rhodamine 123 accumulation in TR-iBRB cells is enhanced in the presence of inhibitors of P-gp [5]. ABCG2 (breast cancer resistance protein BCRP/MXR/ABCP) is also expressed on the 14 Masanori Tachikawa et al. luminal membrane of the inner blood-retinal barrier [48]. ABCG2 recognizes as its substrates not only drugs such as mitoxantrone and doxorubicin, but also photosensitive toxins such as pheophorbide a, a chlorophyll-derived dietary phototoxin related to porphyrin. In vitro studies have demonstrated that ABCG2 is involved in the excretion of pheophorbide a from TR-iBRB cells [48]. Because retina is subject to high levels of cumulative irradiation, ABCG2 may protect this tissue from the light-induced damage caused by a variety of phototoxic compounds including porphyrins. ABCA transporters play an essential role in the efflux of endogenous lipids such as sterols, phospholipids and retinoids from cells. We recently demonstrated that ABCA3 and ABCA9 mRNAs are highly expressed in isolated mouse retinal vascular endothelial cells [43]. Thus, the ABC transporters collectively exhibit a very broad range of substrate selectivity, and work cooperatively at the inner blood-retinal barrier to provide an effective protective mechanism for the retina by restricting the entry of potentially harmful chemicals into retina. But at the same time, this mechanism also poses a significant problem in the delivery of specific drugs into retina for therapeutic purposes. A clear understanding of the various influx and efflux transporters that are expressed at the inner blood-retinal barrier and their detailed substrate selectivity is necessary to determine whether or not a given drug would enter retina from blood in therapeutically relevant concentrations. Influx transporters at the outer blood-retinal barrier RPE cells constitute the outer blood-retinal barrier. These cells form a tight monolayer, separating the neural retina from the choroidal blood circulation. The endothelial cells of the choroidal capillaries are highly fenestrated and thus do not function as a barrier for the transfer of nutrients or xenobiotics. Therefore, it is the monolayer of the RPE cells that provides the barrier function. Since one of the important biologic roles of RPE is to provide essential nutrients to photoreceptors, this cell expresses a multitude of transporters, both on the choroid-facing basolateral membrane and the photoreceptor-facing apical membrane. Even though RPE is a polarized cell similar to the epithelial cells of the intestine and kidney, the distribution of various transporters in the apical versus the basolateral membrane in RPE is strikingly different from that in the other two cell types. For example, the Na+/K+ pump is localized in the blood-facing basolateral membrane in intestinal and renal epithelial cells to mediate the absorption of Na+ from the intestinal or kidney tubular lumen into blood. In contrast, the pump is located not in the blood-facing basolateral membrane but in the apical membrane on the opposite side in RPE. This differential localization is necessary to Drug delivery to retina 15 mediate the transfer of Na+ from blood into neural retina. This is true for several other transporters as well. The “reverse polarity” in RPE is not all that surprising, considering the fact that the RPE cells transport nutrients from blood into neural retina whereas the intestinal and renal epithelial cells transport nutrients into blood. Several different approaches have been used to investigate transport processes in RPE and to identify the transporters that are responsible for these processes. Apical membrane vesicles from RPE can be isolated easily, provided the amount of available tissue is large enough (e.g., bovine eyes). These membrane vesicles are ideal to study the function of apical membrane transporters in a cell-free system. Isolated RPE /choroid preparations have also been used with Ussing chambers to investigate the directional movement of nutrients. Alternatively, RPE cells can be isolated and used directly for transport studies. However, the cells may not polarize well if cultured on impermeable plastic supports. But, when cultured on permeable membrane supports, the cells do polarize with the basolateral membrane in contact with the membrane support and apical membrane away from the membrane support. This culture system is suitable not only for immunocytochemical localization of the transporters present in the apical versus basolateral membranes but also for studies of vectorial transfer of nutrients or xenobiotics in the apical-to-basolateral or basolateral-to-apical direction. A considerable amount of work on the transport characteristics of RPE has also been carried out using the ARPE-19 cell line. This cell line also polarizes when cultured on permeable membrane supports [50]. Recently, primary cultures of RPE cells from human fetal eyes have also been shown to form polarized monolayers on permeable membrane supports under specific culture conditions [51]. With the use of these various approaches, a great deal of information is now available on the identity and characteristics of transporters in the apical and basolateral membranes of the RPE cells to understand the role of various transporters in the handling of xenobiotics and therapeutic drugs at the outer blood-retinal barrier. Transporter-mediated drug delivery across the outer bloodretinal barrier Amino acid mimetic drugs L-DOPA and amino acid mustards represent examples of amino acid mimetic drugs. These compounds are recognized as substrates by the Na+independent amino acid transport system known as System L. At the molecular level, System L is comprised of several isoforms. RPE cells express mRNAs for LAT1 (SLC7A5) and LAT2 (SLC7A8) [52]. Both LAT1 16 Masanori Tachikawa et al. and LAT2 are obligatory amino acid exchangers, meaning that influx of one amino acid substrate into cells is coupled obligatorily to efflux of some other amino acid substrate. LAT1 exhibits high affinity towards its substrates whereas LAT2 shows relatively much lower affinity. In addition, LAT1 prefers bulky neutral amino acids and also recognizes certain D-amino acids; in contrast, LAT2 has much broader substrate selectivity, though its substrates also must be neutral amino acids. Furthermore, LAT2 does not transport D-amino acids. The high affinity transport of D-serine by LAT1 may be of clinical signficance because this D-amino acid functions as a potent co-agonist of the NMDA receptor. Functional studies have confirmed the expression of LAT2 in RPE [53-55]. Interestingly, the transport of leucine, a bulky neutral amino acid, occurs in the apical-to-basolateral direction in RPE [54,55]. This does not mean however that LAT2 functions exclusively in efflux of amino acids from retina into blood across the outer blood-retinal barrier. Since LAT2 is an amino acid exchanger, the efflux of leucine across the cell has to be coupled to influx of certain other amino acids in the blood-to-retina direction. LAT2 is expressed exclusively in the basolateral membrane of intestinal and kidney epithelial cells [56]. Recently, Yamamoto et al [57] found that LAT1 and LAT2 are expressed at the functional level in ARPE-19 cells. At the outer blood-retinal barrier, the amino acid flux is primarily from the circulating blood into retina. The location of LAT2 in RPE seems to be analogous to what is found in the intestinal and renal epithelial cells mediating the efflux of amino acids from the cells (i.e., on the apical membrane of RPE facing the neural retina, but on the basolateral membrane of intestinal and renal epithelial cells facing the blood) [56]. LAT1 may be expressed in RPE on the basolateral membrane, although this remains yet to be established. RPE also expresses mRNA for y+LAT1 (SLC7A7) [52], which transports cationic amino acids such as arginine and lysine in a Na+-independent manner and neutral amino acids in a Na+-dependent manner. Under physiologic conditions, the transport process mediated by y+LAT1 involves Na+-dependent entry of neutral amino acids into cells coupled with exit of cationic amino acids from the cells. Therefore, there may a functional coupling between LAT1/LAT2 and y+LAT1 in the vectorial transfer of amino acid mimetic drugs across RPE, but the polarity of the distribution of these transporters needs to be established to make a rational prediction of their potential in the delivery of such drugs into retina. Antioxidants and other protective compounds We have recently reviewed the transport of vitamin C in the retina and the transporters involved in the process [58]. Since the reduced form of the Drug delivery to retina 17 vitamin (known as ascorbic acid) is transported via the Na+-coupled transporters SVCT1 (SLC23A1) and SVCT2 (SLC23A2) whereas the oxidized form of the vitamin (known as dehydroascorbic acid, DHA) is transported via the facilitative glucose transporter GLUT1 (SLC2A1), the expression and localization of these transporters in RPE would determine the molecular species of the vitamin that is transported, the direction in which the transfer of the vitamin occurs, and the identity of the transporters that are responsible for the process, at the outer blood-retinal barrier. GLUT1 is expressed abundantly in RPE, and is present both at the apical membrane and basolateral membrane [17]. Of the two Na+-coupled vitamin C transporters, RPE expresses predominantly SVCT2 [58]. Cultured RPE cells as well as RPE cell lines take up ascorbic acid in a Na+-dependent manner with high affinity [59-61]. There is no information available on the polarized expression of SVCT2 in RPE, but the functional studies have shown that the Na+-dependent transport of ascorbic acid occurs predominantly at the apical membrane [61]. Based on the expression pattern of GLUT1 and the Na+coupled ascorbic acid transport in RPE, it seems that vitamin C enters RPE cells from choroidal blood primarily in the form of DHA via GLUT1 (Fig. 3A). Once inside the cell, DHA is reduced into ascorbic acid for subsequent use in the cell as an antioxidant. Vitamin C also enters RPE cells from subretinal space via SVCT2 at the apical membrane. The expression of GLUT1 also at the apical membrane suggests that DHA that comes into RPE from choroidal blood via GLUT1 at the basolateral membrane, and which is generated inside the cells from ascorbic acid during antioxidant reactions may be delivered into subretinal space via the apical membrane GLUT1 for use by photoreceptors and other retinal cells. Cystine: Cystine, which plays a critical role in the maintenance of cellular levels of glutathione, is transported into mammalian cells by the amino acid transporter xc-. This transporter consists of the “transporter proper” xCT and the chaperone 4F2hc. Functional and immunocytochemical studies have shown that RPE expresses this transporter [62,63], and that the transporter may play a role in RPE cell proliferation [63]. The expression of the transporter is upregulated in RPE under conditions of increased oxidative stress, indicating a protective role of this transporter as an antioxidant mechanism through glutathione [63,64]. ARPE-19 cells also express xc- robustly; the Michaelis constant for cystine for transport via xc- is in the range of 80-100 M [65]. Expression of the transporter in these cells is markedly upregulated by the transactivator protein Tat encoded by HIV-1 genome [65]. Even though these studies have demonstrated unequivocally the expression of the transporter in RPE, there is no information available on the location of the transporter in the apical versus the basolateral membrane. Creatine: Recently we were comparing the gene 18 Masanori Tachikawa et al. Figure 3. Models for the transport of vitamin C (A), lactate (B), and folate (C) in RPE that constitutes the outer blood-retinal barrier. GLUT1, facilitative glucose transporter 1; SVCT2, sodium-coupled vitamin C transporter 2; DHA, dehydroascorbic acid; AA, ascorbic acid; MCT1, monocarboxylate transporter 1; MCT3, monocarboxylate transporter 3; SMCT1, sodium-coupled monocarboxylate transporter 1; FR , folate receptor ; PCFT, proton-coupled folate transporter; RFC, reduced folate carrier; F, folate and its analogs such as N5-methyltetrahydrofolate and methotrexate. expression pattern between control ARPE-19 cells and ARPE-19 cells transfected with HIV-1 Tat cDNA (unpublished results) using microarray. These studies revealed that ARPE-19 cells express the creatine transporter (SLC6A8) and that the expression is upregulated by HIV-1 Tat. The uptake of creatine in control ARPE-19 cells is Na+- and Cl- -dependent, and saturable. The Michaelis constant for the transport process is 36 6 M. The Na+- and Cl- -activation kinetics of creatine uptake indicated that the Na+:Cl-:creatine stoichiometry is 2:1:1. The expression of HIV-1 Tat in these cells enhanced creatine uptake, and the process was associated with an increase in the steady-state levels of the transporter mRNA as evident from RT-PCR and Northern blot, transporter protein as evident from western blot, and transport activity as evident from the increase in maximal velocity. There was no change in substrate affinity nor in Na+:Cl-:creatine Drug delivery to retina 19 stoichiometry. We have not yet examined the expression of the transporter in primary RPE cells and we also do not know whether or not the transporter is differentially expressed in the apical versus the basolateral membrane. Carnitine and acetylcarnitine: Carnitine is transported at least by two different transport systems, namely the Na+-dependent carnitine transporter OCTN2 (SLC22A5) [66,67] and the Na+- and Cl- -dependent amino acid transporter ATB0,+ (SLC6A14) [68]. Uptake studies with ARPE-19 cells have provided evidence for transport of carnitine via both transporters (unpublished results). Saturation kinetics of Na+-dependent carnitine uptake showed the presence of two distinct transport systems responsible for the observed uptake, one with high affinity (Michaelis constant, 3.2 0.4 M) and the other with low affinity (Michaelis constant, 655 87 M). Based on the known affinities of OCTN2 and ATB0,+, the high affinity uptake is likely to be mediated by OCTN2 and the low affinity uptake by ATB0,+. At micromolar concentrations, carnitine uptake occurs predominantly via OCTN2. This high affinity uptake is inhibited by acetylcarnitine, suggesting that OCTN2 accepts this carnitine derivative as a substrate. In addition to carnitine and its acylderivatives, several other cationic and zwitterionic drugs are transportable substrates for OCTN2. This includes lactam antibiotics [69], tetraethylammonium, pyrilamine, quinidine, verapamil, and valproate [70]. Similarly, SLC6A14 is capable of transporting a wide variety of drugs and prodrugs [71], including nitric oxide synthase inhibitors [72] and amino acid derivatives of antiviral agents such as valacyclovir [73] and valgancicolovir [74]. These data show that RPE expresses OCTN2 and ATB0,+ with the ability to transport carnitine, acylcarnitines, and a multitude of drugs, but the polarity of their expression in the apical versus basolateral membrane remains to be determined. Taurine: Several studies have documented the expression of the Na+/Cl- -coupled taurine transporter (SLC6A6) in RPE [75-79]. There is unequivocal evidence for the presence of the transporter in the apical membrane of this cell. Isolated apical membrane vesicles from bovine RPE demonstrate robust Na+/Cl- -coupled taurine uptake [75,76]. There is also evidence for potential contribution of a -aminobutyrate transporter to the transport of taurine at this membrane [76]. RPE cell lines are highly active in taurine uptake [77-79], and the expression and activity of the transporter are subject to regulation by nitric oxide [78] and by changes in extracellular osmolality [79]. Monocarboxylic drugs Endogenous monocarboxylates such as lactate, pyruvate, and ketone bodies ( -hydroxybutyrate and acetoacetate) are transported in mammalian 20 Masanori Tachikawa et al. cells via two distinct classes of transporters, namely the H+-coupled electroneutral monocarboxylate transporters belonging to the SLC16 gene family (MCTs) and the Na+-coupled electrogenic monocarboxylate transporters belonging to the SLC5 gene family (SMCTs). RPE expresses MCT1 (SLC16A1), MCT3 (SLC16A8), and SMCT1 (SLC5A8). MCT1 is expressed exclusively in the apical membrane of RPE whereas MCT3 is expressed exclusively in the basolateral membrane (Fig. 3B), both in the form of their heterodimeric complexes with the cell surface glycoprotein CD147 [80-82]. Since there is no significant H+ gradient across the apical membrane or the basolateral membrane in RPE in vivo, MCT1 and MCT3 are able to transport their endogenous substrates either into the cell or out of the cell, depending on the concentration gradients for the substrates. Several monocarboxylic drugs have been shown to be substrates for MCT1; this includes foscarnet, salicylate, benzoate, and a prodrug of gabapentin [83,84]. -Hydroxybutyrate, an endogenous monocarboxylate as well as a therapeutic drug, is also a substrate for MCT1 [84]. SMCT1 is expressed only in the basolateral membrane of RPE [85]. Since the transport process mediated by SMCT1 is Na+-coupled and electrogenic [86], the substrates of SMCT1 are actively transported into RPE from choroidal blood (Fig. 3B). The endogenous monocarboxylates and the monocarboxylic drugs that are recognized by SMCT1 as substrates include lactate, pyruvate, short-chain fatty acids such as propionate and butyrate [86], the B-complex vitamin nicotinate [87], ketone bodies [88], -hydroxybutyrate [89], benzoate, salicylate, 5-aminosalicylate [90], and 3-bromopyruvate [91]. Recent studies have shown that the cysteine prodrug L-2-oxothiazolidine carboxylate is a high affinity substrate for SMCT1 [91a], indicating that SMCT1 in the RPE basolateral membrane can be exploited to deliver this drug into cells to increase intracellular glutathione levels as a means to reduce oxidative stress. Interestingly, nonsteroidal anti-inflammatory drugs (e.g., ibuprofen, ketoprofen), which are also monocarboxylates, are not recognized by SMCT1 as transportable substrates; but these drugs function as blockers of the transporter [92]. Folate and folate analogs There are at least three transport proteins that play a role in the uptake of folate and its analogs in mammalian cells. These are the reduced folate carrier (RFC, SLC19A1), folate receptor , and the proton-coupled folate transporter (PCFT, SLC49A1) [93]. All three proteins are expressed in RPE [94-97]. RPE is capable of transcellular transfer of folate, indicating that the folate transport Drug delivery to retina 21 proteins must be expressed in this cell in a polarized manner [98]. Folate receptor is expressed exclusively in the choroid-facing basolateral membrane whereas the reduced folate carrier is expressed exclusively in the apical membrane (Fig. 3C) [96]. Since RPE constitutes the outer blood-retinal barrier, the essential vitamin folate must pass through the cell in the blood-to-retina direction. Folate exists predominantly in the form of N5-methyltetrahydrofolate in plasma, and therefore this form of folate is the principal substrate for the folate transport proteins in RPE. Since the transfer across the basolateral membrane is the first step in the movement of folates across RPE from blood into retina, folate receptor must participate in this first step. However, the folate receptor -mediated entry into cells results in the delivery of folates into endosomes. Therefore, an additional step is needed to deliver folates from the endosomes into cytoplasm. We postulated several years ago that a hitherto unidentified transport system is necessary to mediate this process [99]. The recently cloned proton-coupled folate transporter PCFT has all the features necessary for the delivery of folates from the endosomes into cytoplasm [100]. Since the endosomal compartment is acidic, there exists a H+ gradient across the endosomal membrane in the endosome-to-cytoplasm direction. Such a gradient would be ideal to drive PCFT. PCFT is indeed expressed in RPE [97], but its localization in the basolateral membrane has not been demonstrated. We strongly believe that PCFT is expressed in RPE in a polarized manner with exclusive localization in the basolateral membrane, colocalizing with folate receptor . The transfer of folates across this membrane would involve a functional coupling between the folate receptor and PCFT (Fig. 3C). Folate will bind to the receptor first to initiate endocytosis. Because of the colocalization, endocytosis will result in the transfer of the receptor-folate complex as well as PCFT into endosomes. When the endosomes are acidified subsequently, folate will dissociate from the receptor and then get transferred into the cytoplasm via PCFT. This process will be energized by the transmembrane H+ gradient that exists across the endosomal membrane. The reduced folate carrier in the apical membrane will then facilitate the delivery of folate from the cytoplasm into the subretinal space, thus completing the transcellular transfer. Folate analogs that serve as antifolates (e.g., methotrexate, pemetrexed) can be delivered into neural retina across the outer blood-retinal barrier via the concerted actions of the three folate transport proteins. Recently, Zhao et al have provided evidence for a role for PCFT in folate receptor-mediated endocytosis [101]. With electron microscopic analysis, we have shown that folate receptor and PCFT colocalize in Muller cells on the plasma membrane as well as on the endosomal membrane [102]. Muller cells take up folates using these two transport 22 Masanori Tachikawa et al. proteins. Such studies have not yet been done with RPE, but we hypothesize that PCFT colocalizes with folate receptor in the basolateral membrane and that the two proteins work together in the delivery of folates from choroidal circulation into cells with the participation of endosomes as an intermediate. Biotin, pantothenate, and lipoic acid Biotin and pantothenate are also water-soluble vitamins that exist predominantly in anionic form. As any other mammalian cell, RPE and other retinal cells require these vitamins for their biological function. These vitamins are transported via SMVT, a Na+-coupled mutltivitamin transporter. SMVT also accepts lipoic acid as a substrate. Lipoic acid is a potent antioxidant, and it protects RPE cells from oxidative stress [103,104]. The expression of SMVT in RPE in intact retina has not been studied, but the transporter is expressed in the human RPE cell line ARPE-19 [105]. In addition to the normal physiologic role in the cellular uptake of biotin, pantothenate, and lipoic acid, SMVT also has potential as a delivery system for a variety of prodrugs [105,106]. Organic cationic drugs Endogenous and exogenous organic cations are transported into mammalian cells via a variety of organic cation transporters [107]. One of these transporters is OCT3 (SLC22A3) that transports its substrates in an electrogenic manner. This transporter is expressed in RPE cells [108]. The exact location of the transporter in RPE (apical versus basolateral) is not known, but its substrates of pharmacological and therapeutic significance include prazocin ( -adrenoceptor antagonist), clonidine ( -adrenoceptor agonist), cimetidine (histamine H1 receptor antagonist), verapamil (calcium channel blocker), imipramine and desipramine (antidepressants), quinine (antimalarial drug), and nicotine and methylenedioxymethamphetamine (addictive drugs) [107]. There is also evidence of a novel organic cation transporter in RPE cells that has not been characterized at the molecular level. This transporter transports verapamil, diphenhydramine, pyrilamine, quinidine, and quinacrine [109]. The same transporter may also transport the 2-adrenergic agonist brimonidine [110]. Vitamin E and carotinoids The scavenger receptor class B type 1 (SR-BI) and its splice variant SRBII are expressed in RPE [111,112], and the expression is found both in the apical and basolateral membranes [112]. In various cell types including the Drug delivery to retina 23 retinal blood vessel endothelial cell line TR-iBRB, SR-BI is responsible for the uptake of vitamin E, an antioxidant, either in the free form or when present as a component of HDL [20,113]. Therefore, it is likely that SR-BI mediates the uptake of this vitamin from choroidal blood into RPE. Since most of vitamin E in blood is associated with HDL, SR-BI in RPE does not transport free vitamin E in vivo but rather the HDL-associated vitamin E. The xanthophylls carotenoids such as lutein, zeaxanthin, and lycopene play a significant role in the maintenance of normal vision. These carotenoids are taken up into differentiated ARPE-19 cells via SR-BI [114], suggesting that a similar mechanism might operate in vivo in the uptake of these pigments from blood. Opioid peptides and peptidomimetic drugs RPE cells express two novel oligopeptide transport systems that handle a wide variety of endogenous and synthetic opioid peptides [115,116]. Nonpeptide opioids or non-peptide opioid antagonists do not interact with these transport systems. The two transport systems are called sodium-coupled oligopeptide transporters SOPT1 and SOPT2. These transport systems are distinct from the H+-coupled peptide transporters PEPT1 and PEPT2. In addition to opioid peptides, SOPT1 and SOPT2 also transport other oligopeptides such as the peptide fragments of HIV-1 Tat (e.g., Tat47-57) and HIV-1 Rev (e. g., Rev34-50). These transport systems are upregulated in ARPE-19 cells several-fold by HIV-1 Tat. There is a marked overlap between SOPT1 and SOPT2 in substrate specificity, but the two transport systems are distinguishable based on the opposing effects of small peptides on transport activity. Dipeptides and tripeptides stimulate the activity of SOPT1 but inhibit the activity of SOPT2. The dipeptide opioid kyotorphin is not a substrate for these oligopeptide transport systems, but it stimulates the activity of SOPT1 and inhibits the activity of SOPT2 [116,117]. Peptides consisting of up to 25 amino acids have been shown to interact with these transport systems. SOPT1 and SOPT2 hold great potential for the transport of peptide and peptidomimetic drugs into RPE. The exact location of these transporters in RPE has not yet been established. Organic anion transporting polypeptides This group of transporters mediates Na+-independent transport of a wide variety of organic compounds such as bile salts, dyes, steroid sulfates and glucuronides, anionic oligopeptides, digoxin, thyroid hormones, and prostaglandins. The expression of two of these transporters has been studied in rat retina [118, 119]. Oatp2, encoded by Slco1a4, is expressed prominently 24 Masanori Tachikawa et al. in the apical membrane of RPE [118]. Oatp-E, encoded by Slco4a1, is expressed in RPE, but its exact location is not known [119]. Efflux transporters at the outer blood-retinal barrier RPE cells that constitute the outer blood-retinal barrier play an essential role not only in the delivery of nutrients to retina but also in the protection of this tissue from potential toxic effects of endobiotics and xenobiotics when present in systemic circulation. This protective role of RPE involves prevention of entry of such toxic molecules from choroidal blood into retina. Two different mechanisms participate in this process. The toxic endobiotics and xenobiotics in the systemic circulation might gain entry into RPE either by diffusion or by specific influx transporters in the basolateral membrane. These molecules can be effluxed out of RPE back into systemic blood via specific efflux transporters that are expressed in the RPE basolateral membrane. The second mechanism involves transcellular transfer of endobiotics and xenobiotics from subretinal space into systemic blood via concerted actions of influx transporters in the apical membrane and efflux transporters in the basolateral membrane. While it is certainly true that these efflux processes play a beneficial role in the protection of retina from potentially toxic xenobiotics, these processes also pose a major problem in the delivery of therapeutic drugs to retina. Many of the widely used and clinically relevant drugs are substrates for the efflux transporters and therefore such drugs are actively removed from retina through RPE, thus preventing accumulation of these drugs in retina to therapeutically effective concentrations. This hurdle can be overcome however if specific inhibitors of the efflux transporters are co-administered along with the drugs. This provides a means to prevent the removal of the drugs from retina via the efflux transporters in RPE. P-glycoprotein, BCRP, and MRPs P-glycoprotein, encoded by the MultiDrug Resistance gene MDR1, is expressed in RPE. It is an ATP-dependent efflux pump; its expression has been demonstrated in a number of human RPE cell lines (e.g., D407, h1RPE), but interestingly not in ARPE-19 [120-122]. Several classes of drugs, including anticancer agents, antibiotics, steroids, and immunosuppressants are recognized as substrates for P-glycoprotein. Daunomycin, which is used for the management of proliferative vitreoretinopathy, is a substrate for this efflux pump. Treatment of patients with proliferative vitreoretinopathy using daunomycin causes overexpression of P-glycoprotein, thus resulting in Drug delivery to retina 25 multidrug resistance [123]. This is similar to the multidrug resistance observed in certain cancers due to overexpression of this efflux pump. The transporter is expressed more predominantly in the basolateral membrane facing the choroidal circulation where it can mediate active transfer of its substrates from RPE into blood [120]. Interestingly, it is also found in the apical membrane of RPE where its physiologic function remains unknown. The breast cancer resistance protein BCRP, also known as ABCG2, is expressed in D407 cells but not in any other RPE cell line [122]. RPE in intact retina as well as primary RPE cells express BCRP; the transporter localizes to basolateral membrane of this cell [124]. BCRP is also an efflux pump energized by ATP hydrolysis. There is clear evidence of expression of Multidrug Resistance-associated Protein (MRP) functional activity in ARPE19 cells and in primary cultures of human RPE [125]. The aldose reductase inhibitor BAPSG (N[4-(benzoylamino)phenyl sulfonyl]glycine), which has potential for treatment of diabetic retinopathy, is a substrate for this efflux process [125]. Among the six different genes coding for MRPs, three (MRP1, MRP4, and MRP5) are expressed in all human RPE cell lines [122]. MRPs are also dependent on ATP hydrolysis for active efflux of their substrates. The expression profile of MRPs in intact retina has not been studied. Conclusions and perspectives Even though the neural retina is isolated from systemic circulation via the inner and outer blood-retinal barriers, the plasma membrane transporters expressed at these barrier sites can be exploited for delivery of a wide variety of drugs into retina. An in-depth knowledge of the transporters in the retinal blood vessel endothelial cells and retinal pigment epithelial cells is an absolute requirement to appreciate and exploit the full potential of this approach to deliver therapeutic drugs into the posterior part of the eye for treatment of retinal diseases. Several factors related to specific characteristics of these transporters need to be taken into consideration to maximize their potential use as drug delivery systems. This includes knowledge on relative affinities of these transporters for endogenous substrates versus therapeutic agents to minimize competition, transport capacities to maximize drug delivery for achievement of therapeutically relevant concentrations, and tolerance for structural alterations in substrates to broaden the scope in the design of drugs that can be delivered. We have witnessed a remarkable progress in recent years in the identification and characterization of the transporters in these two cell types that constitute the blood-retinal barriers. This information will be very useful for the future design and development of specific drugs that can be delivered efficiently into retina for therapeutic purposes. 26 Masanori Tachikawa et al. References 1. 2. 3. 4. 5. 6. 7. 8. 9. 10. 11. 12. 13. 14. 15. 16. 17. 18. 19. 20. 21. 22. 23. 24. 25. Lee, V.H.L., and Hosoya, K. 2001, Drug delivery to the posterior segment (Ryan S.J., Ed.), Retina, 3rd edition, Mosby, St. Louis, 2270. Mannermaa, E., Vellonen, K.S., and Urtti, A. 2006, Adv. Drug Delivery, 58, 1136. Hosoya, K., and Tachikawa M. 2009, Pharm. Res., 26, 2055. Hosoya, K., and Tomi, M. 2005, Biol. Pharm. Bull., 28, 1. Hosoya, K., Tomi, M., Ohtsuki, S., Takanaga, H., Ueda, M., Yanai, N., Obinata, M., and Terasaki, T. 2001, Exp. Eye Res., 72, 163. Bodis-Wollner, I. 1997, Clin. Electroencephalogr., 28, 143. Bhaskar, P.A., Vanchilingam, S., Bhaskar, E.A., Devaprabhu, A., and Ganesan, R.A., 1986, Neurology, 36,1119. Uchino, H., Kanai, Y., Kim, D.K., Wempe, M.F., Chairoungdua, A., Morimoto, E., Anders, M.W., and Endou, H. 2002, Mol. Pharmacol., 61, 729. Kageyama, T., Nakamura, M., Matsuo, A., Yamasaki, Y., Takakura, Y., Hashida, M., Kanai, Y., Naito, M., Tsuruo, T., Minato, N., and Shimohama, S. 2000, Brain Res., 879, 115. Tomi, M., Mori, M., Tachikawa, M., Katayama, K., Terasaki, T., and Hosoya, K. 2005, Invest. Ophthalmol. Vis. Sci., 46, 2522. Goldenberg, G.J., Lam, H.Y., and Begleiter, A. 1979, J. Biol. Chem., 254, 1057. Hosoya, K., Kyoko, H., Toyooka, N., Kato, A., Orihashi, M., Tomi, M., and Tachikawa, M. 2008, Biol. Pharm. Bull., 31, 2126. Nagase, K., Tomi, M., Tachikawa, M., and Hosoya, K. 2006, Biochim. Biophys. Acta, 1758, 13. Baldwin, S.A., Beal, P.R., Yao, S.Y., King, A.E., Cass, C.E., and Young, J.D. 2004, Pflugers. Arch., 447, 735. Yao, S.Y., Ng, A.M., Sundaram, M., Cass, C.E., Baldwin, S.A., and Young, J.D. 2001, Mol. Membr. Biol., 18, 161. Hosoya, K., Minamizono, A., Katayama, K., Terasaki, T., and Tomi, M., 2004, Invest. Ophthalmol. Vis. Sci., 45, 1232. Takata, K., Kasahara, T., Kasahara, M., Ezaki, O., and Hirano, H. 1992, Invest. Ophthalmol. Vis. Sci., 33, 377. Minamizono, A., Tomi, M., and Hosoya, K., 2006, Biol. Pharm. Bull., 29, 2148. Goti, D., Hrzenjak, A., Levak-Frank, S., Frank, S., van der Westhuyzen, D.R., Malle, E., and Sattler, W. 2001, J. Neurochem., 76, 498. Tachikawa, M., Okayasu, S., and Hosoya, K. 2007, Mol. Vis.,13, 2041. Tomi, M., Hosoya, K., Takanaga, H., Ohtsuki, S., and Terasaki, T. 2002, Invest. Ophthalmol. Vis. Sci., 43, 774. Hosoya, K., Saeki, S., and Terasaki, T. 2001, Microvasc. Res., 62, 136. Nakashima, T., Tomi, M., Katayama, K., Tachikawa, M., Watanabe, M., Terasaki, T., and Hosoya, K. 2004, J. Neurochem., 89, 1454. Tachikawa, M., Takeda, Y., Tomi, M., and Hosoya, K. 2009, Invest. Ophthalmol. Vis. Sci., in press. Tomi, M., Terayama, T., Isobe, T., Egami, F., Morito, A., Kurachi, M., Ohtsuki, S., Kang, Y. S., Terasaki, T., and Hosoya, K. 2007, Microvasc. Res., 73, 100. Drug delivery to retina 27 26. Tomi, M., Tajima, A., Tachikawa, M., and Hosoya, K. 2008, Biochim. Biophys. Acta., 1778, 2138. 27. Tomi, M., Kitade, N., Hirose, S., Yokota, N., Akanuma, S., Tachikawa, M., and Hosoya, K. 2009, J. Neurochem., 11, 716. 28. Hosoya, K., Kondo, T., Tomi, M., Takanaga, H., Ohtsuki, S., and Terasaki, T. 2001, Pharm. Res., 18, 1669. 29. Gerhart, D.Z., Leino, R.L., and Drewes, L.R. 1999, Neuroscience, 92, 367. 30. Hosoya, K., Fujita, K., and Tachikawa, M. 2008, Drug Metab. Pharmacokinet., 23, 285. 31. Tomi, M., Arai, K., M. Tachikawa, M., and Hosoya, K. 2007, Neurochem. Res., 32, 1833. 32. Okamoto, M., Akanuma, S., Tachikawa, M., and Hosoya, K. 2009, Neurochem. Int., 55, 789. 33. Barza, M., Kane, A., and Baum, J. 1983, Invest. Ophthalmol. Vis. Sci., 24, 1602. 34. Somervaille, T.C., Hann, I.M, Harrison, G., Eden, T.O., Gibson, B.E., Hill, F.G.., Mitchell, C., Kinsey, S.E., Vora, A.J., and Lilleyman, J.S. 2003, Br. J. Haematol., 121, 280. 35. Engler, C.B., Sander, B., Larsen, M., Koefoed, P., Parving, H.H., LundAndersen, H. 1994, Acta. Ophthalmol., 72, 663. 36. Betz, A.L., and Goldstein, G.W. 1980, Exp. Eye Res., 30, 593. 37. Hosoya, K., Makihara, A., Tsujikawa, Y., Yoneyama, D., Mori, S., Terasaki, T., Akanuma, S., Tomi, M., and Tachikawa, M. 2009, J. Pharmacol. Exp. Ther., 329, 87. 38. Kikuchi, R., Kusuhara, H., Sugiyama, D., and Sugiyama, Y., 2003, J. Pharmacol. Exp. Ther., 306, 51. 39. Nakakariya, M., Shimada, T., Irokawa, M., Koibuchi, H., Iwanaga, T., Yabuuchi, H., Maeda, T., and Tamai, I. 2008, Pharm. Res., 25, 578. 40. Gao, B., Wenzel, A., Grimm, C., Vavricka, S.R., Benke, D., Meier, P.J., and Reme, C.E. 2002, Invest. Ophthalmol. Vis. Sci. 43, 510. 41. Katayama, K., Ohshima, Y., Tomi, M., and Hosoya, K. 2006, J. Neurosci. Methods, 156, 249. 42. Uchida, Y., Kamiie, J., Ohtsuki, S., and Terasaki, T. 2007, Pharm. Res., 24, 2281. 43. Tachikawa, M., Toki, H., Tomi, M., and Hosoya, K. 2008, Microvasc. Res., 75, 68. 44. Leggas, M., Adachi, M., Scheffer, G.L., Sun, D., Wielinga, P., Du, G., Mercer, K.E., Zhuang, Y., Panetta, J.C., Johnston, B., Scheper, R.J., Stewart, C.F., and Schuetz, J.D. 2004, Mol. Cell Biol., 24, 7612. 45. Tomi, M., and Hosoya, K. 2004, J. Neurochem., 91, 1244. 46. Sugiyama, D., Kusuhara, H., Shitara, Y., Abe, T., Meier, P.J., Sekine, T., Endou, H., Suzuki, H., and Sugiyama, Y. 2001, J. Pharmacol. Exp. Ther., 298, 316. 47. BenEzra, D., and Maftzir, G. 1990, Invest. Ophthalmol. Vis. Sci., 31, 1362. 48. Asashima, T., Hori, S., Ohtsuki, S., Tachikawa, M., Watanabe, M., Mukai, C., Kitagaki, S., Miyakoshi, N., and Terasaki, T. 2006, Pharm. Res., 23, 1235. 49. Puchowicz, M.A., Xu, K., Magness, D., Miller, C., Lust, W.D., Kern, T.S., and LaManna, J.C. 2004, J. Cereb. Blood Flow Metab., 24, 449. 50. Dunn, K.C., Aotaki-Keen, A.E., Putkey, F.R., and Hjelmeland, L.M. 1996, Exp. Eye Res., 62, 155. 28 Masanori Tachikawa et al. 51. Sonoda, S., Spee, C., Barron, E., Ryan, S.J., Kannan, R., and Hinton, D.R. 2009, Nat. Protoc., 4, 662. 52. Nakauchi, T., Ando, A., Ueda-Yamada, M., Yamazaki, Y., Uyama, M., Matsumura, M., and Ito, S. 2003, Invest. Ophthalmol. Vis. Sci., 44, 5023. 53. Sellner, P.A. 1986, J. Neurosci., 6, 2823. 54. Pautler, E.L., Tengerdy, C., Beyer, J., and Beezley, D. 1989, Am. J. Physiol., 257, C940. 55. Gandhi, M.D., Pal, D., and Mitra, A.K. 2004, Int. J. Pharm., 275, 189. 56. Rossier, G., Meier, C., Bauch, C., Summa, V., Sordat, B., Verrey, F., and Kuhn, L.C. 1999, J. Biol. Chem., 274, 34948. 57. Yamamoto, A., Akanuma, S., Tachikawa, M., and Hosoya, K. 2010, J. Pharm. Sci., 99, 2475. 58. Ganapathy, V., Ananth, S., Smith, S.B., and Martin, P.M. 2008, Ocular Transporters in Ophthalmic Diseases and Drug Delivery, J. Tombran-Tink and C.J. Barnstable (Eds.), Humana Press, 437. 59. Lam, K.W., Yu, H.S., Glickman, R.D., and Lin, T. 1993, Ophthalmic Res., 25, 100. 60. Khatami, M., Stramm, L.E., and Rockey, J.H. 1986, Exp. Eye Res., 43, 607. 61. Dimattio, J., and Streitman, J. 1991, Curr. Eye Res., 10, 959. 62. Dun, Y., Mysona, B., Van Ells, T., Amarnath, L., Ola, M.S., Ganapathy, V., and Smith, S.B. 2006, Cell Tissue Res., 324, 189. 63. Gnanaprakasam, J.P., Thangaraju, M., Liu, K., Ha, Y., Martin, P.M., Smith, S.B., and Ganapathy, V. 2009, Biochem. J., 424, 243. 64. Bridges, C.C., Kekuda, R., Wang, H., Prasad, P.D., Mehta, P., Huang, W., Smith, S.B., and Ganapathy, V. 2001, Invest. Ophthalmol. Vis. Sci., 42, 47. 65. Bridges, C.C., Hu, H., Miyauchi, S., Siddaramappa, U.N., Ganapathy, M.E., Ignatowicz, L., Maddox, D.M., Smith, S.B., and Ganapathy, C. 2004, Invest. Ophthalmol. Vis. Sci., 45, 2906. 66. Tamai, I., Ohashi, R., Nezu, J., Yabuuchi, H., Oku, A., Shimane, M., Sai, Y., and Tsuji, A. 1998, J. Biol. Chem., 273, 20378. 67. Wu, X., Huang, W., Prasad, P.D., Seth, P., Rajan, D.P., Leibach, F.H., Chen, J., Conway, S.J., and Ganapathy, V. 1999, J. Pharmacol. Exp. Ther., 290, 1482. 68. Nakanishi, T., Hatanaka, T., Huang, W., Prasad, P.D., Leibach, F.H., Ganapathy, M.E., and Ganapathy, V. 2001, J. Physiol., 532, 297. 69. Ganapathy, M.E., Huang, W., Rajan, D.P., Carter, A.L., Sugawara, M., Iseki, K., Leibach, F.H., and Ganapathy, V. 2000, J. Biol. Chem., 275, 1699. 70. Ohashi, R., Tamai, I., Yabuuchi, H., Nezu, J.I., Oku, A., Sai, Y., Shimane, M., and Tsuji, A. 1999, J. Pharmacol. Exp. Ther., 291, 778. 71. Ganapathy, M.E., and Ganapathy, V. 2005, Curr. Drug Targets Immune Endocr. Metabol. Disord., 5, 357. 72. Hatanaka, T., Nakanishi, T., Huang, W., Leibach, F.H., Prasad, P.D., Ganapathy, V., and Ganapathy, M.E. 2001, J. Clin. Invest., 107, 1035. 73. Hatanaka, T., Haramura, M., Fei, Y.J., Miyauchi, S., Bridges, C.C., Ganapathy, P.S., Smith, S.B., Ganapathy, V., and Ganapathy, M.E. 2004, J. Pharmacol. Exp. Ther., 308, 1138. Drug delivery to retina 29 74. Umapathy, N.S., Ganapathy, V., and Ganapathy, M.E. 2004, Pharm. Res., 21, 1303. 75. Miyamoto, Y., Kulanthaivel, P., Leibach, F.H., and Ganapathy, V. 1991, Invest. Ophthalmol. Vis. Sci., 32, 2542. 76. Sivakami, S., Ganapathy, V., Leibach, F.H., and Miyamoto, Y. 1992, Biochem. J., 283, 391. 77. Leibach, J.W., Cool, D.R., Del Monte, M.A., Ganapathy, V., Leibach, F.H., and Miyamoto, Y. 1993, Curr. Eye Res., 12, 29. 78. Bridges, C.C., Ola, M.S., Prasad, P.D., El-Sherbeny, A., Ganapathy, V., and Smith, S.B. 2001, Am. J. Physiol., 281, C1825. 79. El-Sherbeny, A., Naggar, H., Miyauchi, S., Ola, M.S., Maddox, D.M., Martin, P.M., Ganapathy, V., and Smith, S.B. 2004, Invest. Ophthalmol. Vis. Sci., 45, 694. 80. Philp, N.J., Yoon, H., and Grollman, E.F. 1998, Am. J. Physiol., 274, R1824. 81. Philp, N.J., Wang, D., Yoon, H., and Hjelmeland, L.M. 2003, Invest. Ophthalmol. Vis. Sci., 44, 1716. 82. Deora, A.A., Philp, N.J., Hu, J., Bok, D., and Rodriguez-Boulan, E. 2005, Proc. Natl. Acad. Sci. U.S.A., 102, 16245. 83. Enerson, B.E., and Drewes, L.R. 2003, J. Pharm. Sci., 92, 1531. 84. Morris, M.E., and Felmlee, M.A. 2008, AAPS J., 10, 311. 85. Martin, P.M., Dun, Y., Mysona, B., Ananth, S., Roon, P., Smith, S.B., and Ganapathy, V. 2007, Invest. Ophthalmol. Vis. Sci., 48, 3356. 86. Miyauchi, S., Gopal, E., Fei, Y.J., and Ganapathy, V. 2004, J. Biol. Chem., 279, 13293. 87. Gopal, E., Fei, Y.J., Miyauchi, S., Zhuang, L., Prasad, P.D., and Ganapathy, V. 2005, Biochem. J., 388, 309. 88. Martin, P.M., Gopal, E., Ananth, S., Zhuang, L., Itagaki, S., Prasad, B.M., Smith, S.B., Prasad, P.D., and Ganapathy, V. 2006, J. Neurochem., 98, 279. 89. Ganapathy, V., Thangaraju, M., Gopal, E., Martin, P.M., Itagaki, S., Miyauchi, S., and Prasad, P.D. 2008, AAPS J., 10, 193. 90. Gopal, E., Miyauchi, S., Martin, P.M., Ananth, S., Roon, P., Smith, S.B., and Ganapathy, V. 2007, Pharm. Res., 24, 575. 91. Thangaraju, M., Karunakaran, S., Itagaki, S., Gopal, E., Elangovan, S., Prasad, P.D., and Ganapathy, V. 2009, Cancer 115, 4655. 91a. Babu, E., Ananth, S., Veeranan-Karmegam, R., Coothankandaswamy, V., Smith, S.B., Boettger, T., Ganapathy, V., and Martin, P.M. 2011, Invest. Ophthalmol. Vis. Sci., 52, 5749. 92. Itagaki, S., Gopal, E., Zhuang, L., Fei, Y.J., Miyauchi, S., Prasad, P.D., and Ganapathy, V. 2006, Pharm. Res., 23, 1209. 93. Zhao, R., Matherly, L.H., and Goldman, I.D. 2009, Expert Rev. Mol. Med., 11, e4. 94. Huang, W., Prasad, P.D., Kekuda, R., Leibach, F.H., and Ganapathy, V. 1997, Invest. Ophthalmol. Vis. Sci., 38, 1578. 95. Smith, S.B., Kekuda, R., Gu, X., Chancy, C., Conway, S.J., and Ganapathy, V. 1999, Invest. Ophthalmol. Vis. Sci., 40, 840. 96. Chancy, C.D., Kekuda, R., Huang, W., Prasad, P.D., Kuhnel, J.M., Sirotnak, F.M., Roon, P., Ganapathy, V., and Smith, S.B. 2000, J. Biol. Chem., 275, 20676. 30 Masanori Tachikawa et al. 97. Umapathy, N.S., Gnanaprakasam, J.P., Martin, P.M., Mysona, B., Dun, Y., Smith, S.B., Ganapathy, V., and Prasad, P.D. 2007, Invest. Ophthalmol. Vis. Sci., 48, 5299. 98. Bridges, C.C., El-Sherbeny, A., Ola, M.S., Ganapathy, V., and Smith, S.B. 2002, Curr. Eye Res., 24, 129. 99. Ganapathy, V., Smith, S.B., and Prasad, P.D. 2004, Pflugers Arch., 447, 641. 100. Qiu, A., Jansen, M., Sakaris, A., Min, S.H., Chattopadhyay, S., Tsai, E., Sandoval, C., Zhao, R., Akabas, M.H., and Goldman, I.D. 2006, Cell, 127, 917. 101. Zhao, R., Min, S.H., Wang, Y., Campanella, E., Low, P.S., and Goldman, I.D. 2009, J. Biol. Chem., 284, 4267. 102. Bozard, B.R., Ganapathy, P.S., Duplantier, J., Mysona, B., Ha, Y., Roon, P., Smith, R., Goldman, I.D., Prasad, P., Martin, P.M., Ganapathy, V., and Smith, S.B. 2010, Invest. Opthalmol. Vis. Sci., 51, 3226. 103. Voloboueva, L.A., Liu, J., Suh, J.H., Ames, B.N., and Miller, S.S. 2005, Invest. Ophthalmol. Vis. Sci., 46, 4302. 104. Jia, L., Liu, Z., Sun. L., Miller, S.S., Ames, B.N., Cotman, C.W., and Liu, J. 2007, Invest. Ophthalmol. Vis. Sci., 48, 339. 105. Janoria, K.G., Boddu, S.H., Wang, Z., Paturi, D.K., Samanta, S., Pal, D., and Mitra, A.K. 2009, J. Ocul. Pharmacol. Ther., 25, 39. 106. Cundy, K.C., Branch, R., Chernov-Rogan, T., Dias, T., Estrada, T., Hold, K., Koller, K., Liu, X., Mann, A., Panuwat, M., Raillard, S.P., Upadhyay, S., Wu, Q.Q., Xiang, J.N., Yan, H., Zerangue, N., Zhou, C.X., Barrett, R.W., and Gallop, M.A. 2004, J. Pharmacol. Exp. Ther., 311, 315. 107. Koepsell, H., Lips, K., and Volk, C. 2007, Pharm. Res., 24, 1227. 108. Rajan, P.D., Kekuda, R., Chancy, C.D., Huang, W., Ganapathy, V., and Smith, S.B. 2000, Curr. Eye Res., 20, 195. 109. Han, Y.H., Sweet, D.H., Hu, D.N., and Pritchard, J.B. 2001, J. Pharmacol. Exp. Ther., 296, 450. 110. Zhang, N., Kannan, R., Okamoto, C.T., Ryan, S.J., Lee, V.H.L., and Hinton, D.R. 2006, Invest. Ophthalmol. Vis. Sci., 47, 287. 111. Duncan, K.G., Hosseini, K., Bailey, K.R., Yang, H., Lowe, R.J., Matthes, M.T., Kane, J.P., LaVail, M.M., Schwartz, D.M., and Duncan, J.L. 2009, Br. J. Ophthalmol., 93, 1116. 112. Tserentsoodol, N., Gordiyenko, N.V., Pascual, I., Lee, J.W., Fliesler, S.J., and Rodriguez, I.R. 2006, Mol. Vis., 12, 1319. 113. Reboul, E., Klein, A., Bietrix, F., Gleize, B., Malezet-Desmoulins, C., Schneider, M., Margotat, A., Lagrost, L., Collet, X., and Borel, P. 2006, J. Biol. Chem., 281, 4739. 114. During, A., Doraiswamy, S., and Harrison, E.H. 2008, J. Lipid Res., 49, 1715. 115. Hu, H., Miyauchi, S., Bridges, C.C., Smith, S.B., and Ganapathy, V. 2003, Biochem. J., 375, 17. 116. Chothe, P., Thakkar, S.V., Gnanaprakasam, J.P., Ananth, S., Hinton, D.R., Kannan, R., Smith, S.B., Martin, P.M., and Ganapathy, V. 2010, Invest. Ophthalmol. Vis. Sci., 51, 413. Drug delivery to retina 31 117. Thakkar, S.V., Miyauchi, S., Prasad, P.D., and Ganapathy, V. 2008, Drug Metab. Pharmacokinet., 23, 254. 118. Ito, A., Yamaguchi, K., Onogawa, T., Unno, M., Suzuki, T., Nishio, T., Suzuki, T., Sasano, H., Abe, T., and Tamai, M. 2002, Invest. Ophthalmol. Vis. Sci., 43, 858. 119. Ito, A., Yamaguchi, K., Tomita, H., Suzuki, T., Onogawa, T., Sato, T., Mizutamari, H., Mikkaichi, T., Nishio, T., Suzuki, T., Unno, M., Sasano, H., Abe, T., and Tamai, M. 2003, Invest. Ophthalmol. Vis. Sci., 44, 4877. 120. Kennedy, B.G., and Mangini, N.J. 2002, Mol. Vis., 8, 422. 121. Constable, P.A., Lawrenson, J.G., Dolman, D.E., Arden, G.B., and Abbott, N.J. 2006, Exp. Eye Res., 83, 24. 122. Mannermaa, E., Vellonen, K.S., Ryhanen, T., Kokkonen, K., Ranta, V.P., Kaarniranta, K., and Urtti, A. 2009, Pharm. Res., 26, 1785. 123. Esser, P., Tervooren, D., Heimann, K., Kociok, N., Bartz-Schmidt, K.U., Walter, P., and Weller, M. 1998, Invest. Ophthalmol. Vis. Sci., 39, 164. 124. Gnana-Prakasam, J.P., Reddy, S.K., Veeranan-Karmegam, R., Smith, S.B., Martin, P.M., and Ganapathy, V. Invest. Ophthalmol. Vis. Sci., 52, 9279. 125. Aukunuru, J.V., Sunkara, G., Bandi, N., Thoreson, W.B., and Kompella, U.B. 2001, Pharm. Res., 18, 565.