Survey
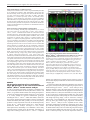
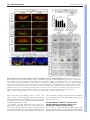
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
RESEARCH ARTICLE 4249 Development 137, 4249-4260 (2010) doi:10.1242/dev.053819 © 2010. Published by The Company of Biologists Ltd The transcription factors Nkx2.2 and Nkx2.9 play a novel role in floor plate development and commissural axon guidance Andreas Holz1, Heike Kollmus1,*, Jesper Ryge2, Vera Niederkofler3, Jose Dias4, Johan Ericson4, Esther T. Stoeckli3, Ole Kiehn2 and Hans-Henning Arnold1,† SUMMARY The transcription factors Nkx2.2 and Nkx2.9 have been proposed to execute partially overlapping functions in neuronal patterning of the ventral spinal cord in response to graded sonic hedgehog signaling. The present report shows that in mice lacking both Nkx2 proteins, the presumptive progenitor cells in the p3 domain of the neural tube convert to motor neurons (MN) and never acquire the fate of V3 interneurons. This result supports the concept that Nkx2 transcription factors are required to establish V3 progenitor cells by repressing the early MN lineage-specific program, including genes like Olig2. Nkx2.2 and Nkx2.9 proteins also perform an additional, hitherto unknown, function in the development of non-neuronal floor plate cells. Here, we demonstrate that loss of both Nkx2 genes results in an anatomically smaller and functionally impaired floor plate causing severe defects in axonal pathfinding of commissural neurons. Defective floor plates were also seen in Nkx2.2+/–;Nkx2.9–/– compound mutants and even in single Nkx2.9–/– mutants, suggesting that floor plate development is sensitive to dose and/or timing of Nkx2 expression. Interestingly, adult Nkx2.2+/–;Nkx2.9–/– compound-mutant mice exhibit abnormal locomotion, including a permanent or intermittent hopping gait. Drug-induced locomotor-like activity in spinal cords of mutant neonates is also affected, demonstrating increased variability of left-right and flexor-extensor coordination. Our data argue that the Nkx2.2 and Nkx2.9 transcription factors contribute crucially to the formation of neuronal networks that function as central pattern generators for locomotor activity in the spinal cord. As both factors affect floor plate development, control of commissural axon trajectories might be the underlying mechanism. INTRODUCTION The spinal cord contains anatomically distinct classes of neurons that contribute to sensory and motor tasks. Neurons that exert and regulate motor control reside in the ventral spinal cord, and their development in the mammalian embryo is largely controlled by genetic programs (for reviews, see Briscoe and Novitch, 2008; Jessell, 2000; Shirasaki and Pfaff, 2002) that specify at least five classes of neuronal progenitor cells. These progenitors give rise to motor neurons (MNs) and four cardinal classes of ventral interneurons, referred to as V0 to V3 neurons (Ericson et al., 1997a; Pierani et al., 1999). The different progenitor cell types are generated in response to the graded activity of the signaling protein sonic hedgehog (Shh), which is first produced by the notochord and subsequently by the overlying floor plate, the most ventral structure of the neural tube (Chiang et al., 1996; Ericson et al., 1996; Marti et al., 1995; Roelink et al., 1995). The concentration gradient of Shh along the dorsoventral (DV) axis of the neural tube leads to regional activation or repression of homeodomain (HD) transcription factors, generating a transcriptional code for distinct 1 Cell and Molecular Biology, University of Braunschweig, Spielmannstraße 7, 38106 Braunschweig, Germany. 2Mammalian Locomotor Laboratory, Department of Neuroscience, Karolinska Institute, 17177 Stockholm, Sweden. 3Institute of Molecular Life Sciences, University of Zurich, 8057 Zurich, Switzerland. 4Institute of Cell and Molecular Biology, Karolinska Institute, 17177 Stockholm, Sweden. *Present address: Department of Infection Genetics, Helmholtz Centre for Infection Research, Inhoffenstraße 7, 38124 Braunschweig, Germany † Author for correspondence ([email protected]) Accepted 11 October 2010 progenitor cell populations (Briscoe and Ericson, 2001; Briscoe et al., 2000; Briscoe et al., 1999; Ericson et al., 1997a; Ericson et al., 1997b; Novitch et al., 2001). The most ventral neuronal population in the spinal cord, referred to as Sim1-expressing (Sim1+) V3 interneurons, is derived from progenitor cells expressing Nkx2.2 and Nkx2.9 HD transcription factors. This cell population is located immediately dorsal to the floor plate and encounters high concentrations of Shh signal. Both Nkx2 genes are structurally, and presumably functionally, related and exhibit very similar spatiotemporal expression patterns in the central nervous system (CNS) in a Shh-dependent manner (Briscoe et al., 1999; Pabst et al., 1998; Pabst et al., 2000). In Nkx2.2-deficient mice, the domain (p3) of early V3 progenitor cells forms and is indistinguishable from wild type between the ventral floor plate and dorsal Pax6expressing cells. However, p3 progenitors in the mutant mouse undergo a ventral-to-dorsal transformation and generate MNs in place of Sim1+ V3 interneurons (Briscoe et al., 1999; Ericson et al., 1997a). This indicates that Nkx2.2 is essential for the correct differentiation of V3 neurons but not for establishing the p3 progenitor domain. Nkx2.9, which is initially co-expressed with Nkx2.2, might substitute for the missing Nkx2.2 function. Slightly later, when expression of Nkx2.9 stops in the neural tube at embryonic day (E) 10.5 (Briscoe et al., 1999), the essential requirement of Nkx2.2 to establish V3 neuronal fate becomes apparent. Alternatively, Nkx2.2 might play no role in setting up p3 progenitor cells and entirely different factors could be responsible. Targeted disruption of the Nkx2.9 gene in mouse causes no obvious neuronal phenotype in spinal cord, consistent with the idea that Nkx2.2, which is expressed for a longer time DEVELOPMENT KEY WORDS: Nkx2.2, Nkx2.9, Floor plate development, Central pattern generators in spinal cord, Mouse 4250 RESEARCH ARTICLE mutant embryos appear to be transformed into MN progenitors leading to considerably reduced expression of various floor platespecific markers. Moreover, we observe axonal pathfinding defects of commissural axons that might be related to the loss of floor plate integrity in mutants. Finally, we demonstrate that Nkx2.2+/–;Nkx2.9–/– compound mutants exhibit locomotor defects in vivo and during fictive locomotor-like activity in vitro, indicating that both HD transcription factors contribute to the development and function of CPG circuits in the spinal cord. MATERIALS AND METHODS Generation and genotyping of the Nkx2.2;Nkx2.9 double-mutant mouse Double-homozygous mice were obtained by crossing Nkx2.2- (Sussel et al., 1998) and Nkx2.9- (Pabst et al., 2003) deficient mutant mice. Doublenull mutants showed perinatal lethality, as already reported for the Nkx2.2–/– mouse (Sussel et al., 1998); however, Nkx2.2+/–;Nkx2.9–/– mutants were viable. Genotyping for Nkx2.9 was performed by PCR as described previously (Pabst et al., 2003). Nkx2.2 genotyping was done by Southern blotting as described (Sussel et al., 1998) or by PCR using the following primers: Nkx2.2 #6 sense, 5⬘-TTCCAAAGGCACCACAAATCGC-3⬘; Nkx2.2 #7 antisense, 5⬘-GGTCTTGGGAGTCAAGTGGATGAAG-3⬘; Nkx2.2 mut#1 antisense, 5⬘-TGAAGAACGAGATCAGCAGCCTCT-3⬘. Embryos were staged by counting the morning of vaginal plug detection as E0.5. Immunohistochemistry Immunohistochemical detection of proteins was performed on fixed tissue sections of mouse embryos as described previously (Pabst et al., 2003). Primary mouse-specific monoclonal antibodies (obtained from the Developmental Studies Hybridoma Bank, University of Iowa) used were: FoxA2 (Foxa2 – Mouse Genome Informatics) clone 4C7 (1:800), Nkx2.2 clone 74.5A5 (1:1000), Nkx6.1 clone F55A10 (1:600), Pax6 (1:600), HB9/MNR2 (Mnx1 – Mouse Genome Informatics) clone 81.5C10 (1:300), Islet1 (Isl1 – Mouse Genome Informatics) clone 39.4D5 (1:500), Lim3 (Lhx3 – Mouse Genome Informatics) clone 67.4E12 (1:500) and Tag1 (Cntn2 – Mouse Genome Informatics) clone 4D7 (1:15). Other primary antibodies used were: rabbit anti-Olig2 (IBL; 1:400), anti-BrdU (Santa Cruz; 1:400) and rabbit anti-Rig1 (a gift from Dr Fujio Murakami, Osaka University, Japan; 1:600) (Tamada et al., 2008). Secondary antibodies used were: anti-mouse IgG-Cy2, anti-mouse IgG1-Cy2, anti-mouse IgG2b-Cy3 and anti-rabbit IgG-Cy5 (all from Jackson ImmunoResearch; 1:600). The slides were counterstained with DAPI (Molecular Probes) and documented on a Zeiss Apotome or Zeiss LSM 510 confocal microscope. Galactosidase activity was detected on cryosections (14-16 m) as described (Pabst et al., 2003) and documented with a digital color camera mounted onto a Leica DM RBE microscope. For quantification, specifically marked cells on sections (16 m) of neural tube were photographed on the Zeiss Axioplan 2 fluorescent microscope and counted using Adobe Photoshop software. For each genotype, a minimum of five sections from at least three individual animals was analyzed. Statistical analysis (t-test) and graph preparation was carried out using GraphPad software (San Diego). In situ hybridization and riboprobes Strand-specific digoxigenin-labeled riboprobes were used for in situ hybridization as described previously (Pabst et al., 2003). The following gene fragments from mouse were subcloned into pSP70/71 vectors: sonic hedgehog, nucleotides 1650–2668 (accession number AK077688); slit, ClaI-SmaI restriction fragment of IMAGE clone IRAVp968B10159 (ImaGenes, Berlin); Dbx1, SmaI-HindIII fragment of IMAGE clone IMAGp998C241002 (ImaGenes, Berlin); Chx10 (Vsx2 – Mouse Genome Informatics), HindIII-XhoI Chx10 3⬘UTR fragment of IMAGE clone IRAVp968a0893 (ImaGenes, Berlin). IMAGE clone IRAKp961D0140Q (ImaGenes, Berlin) was used to generate netrin 3⬘ untranslated region (UTR) riboprobes, the FoxA2 riboprobe was described previously (Ang et al., 1993) and a 5⬘ probe of engrailed 1 cDNA was obtained from F. Vauti (University of Braunschweig, Germany) (Wurst et al., 1994). DEVELOPMENT period, provides redundant functions and thereby rescues V3 neurons (Pabst et al., 2003). Thus, the individual phenotypes of single Nkx2.2 and Nkx2.9 mutant mice suggest that the two genes have overlapping roles in neuronal patterning of the ventral spinal cord. In order to clarify the extent of shared functions between the two transcription factors, we examined double-mutant mice lacking both Nkx2.2 and Nkx2.9 proteins with particular focus on their individual and combined roles in early cell lineage decisions in the spinal cord. Although floor plates were reported to be anatomically normal in Nkx2.2 and Nkx2.9 single-mutant mice (Briscoe et al., 1999; Pabst et al., 2003), early expression of both regulators in the floor plate area suggests that they might have a function in development and/or maintenance of this important organizing and signaling center. Nkx2.2 expression in the ventral spinal cord of zebrafish and chicken embryos has been shown to extend beyond the p3 domain into the lateral floor plate (Chapman et al., 2002; Strahle et al., 2004). In the mouse, expression of Nkx2.2 in early floor plate has also been described (Jeong and McMahon, 2005) and preliminary evidence showed that the Nkx2.9 knock-in allele directs lacZ expression in the most ventral part of the neural tube (Pabst et al., 2003). The floor plate is not composed of a single population of uniform cells but consists of different cell groups, most notably the medial and the lateral floor plate cells, which can be distinguished by molecular markers including Nkx2.2 (Placzek and Briscoe, 2005; Strahle et al., 2004). To investigate the hypothesis that both HD transcription factors could play a role in floor plate development, we utilized Nkx2.2;Nkx2.9 doublemutant embryos to analyze the histological and functional consequences of the combined null mutations on floor plate development. The neuronal circuits that generate locomotor behavior in mammals are embedded in the ventral half of the spinal cord (Kiehn, 2006; Kiehn and Kjaerulff, 1998). These networks are commonly called central pattern generators (CPGs), which are composed of excitatory and inhibitory neurons belonging to the four main classes of V0, V1, V2 and V3 interneurons (Briscoe et al., 2000; Goulding and Pfaff, 2005; Jessell, 2000; Kiehn, 2006) with further subtypes within each class (Al-Mosawie et al., 2007; Lanuza et al., 2004; Lundfald et al., 2007). Individual contributions of the various types of interneurons to locomotor CPGs have been investigated by genetic disruption of early cell type-specific transcription factors, selective cell ablation experiments and permanent or acute blockage of synaptic transmission (for reviews, see Goulding, 2009; Kiehn, 2006; Kiehn et al., 2010; Stepien and Arber, 2008). The role of V3 neurons in the CPG was probed by functional interference in two ways. First, a Sim1-Cre-mediated conditionally expressed tetanus toxin light chain permanently blocked synaptic release from V3 cells; secondly, an allostatin receptor-based approach acutely attenuated the activity of V3 neurons in the adult mouse. Both experimental interventions resulted in disruption of the reliable rhythmicity of locomotor activity in vitro without a major effect on left-right alternation (Zhang et al., 2008). Nkx2.2 single- and Nkx2.2;Nkx2.9 doublemutant mice fail to form Sim1+ V3 interneurons thereby providing an alternative model to study the role of V3 neurons in the spinal locomotor network. Here, we report the detailed phenotypic analysis of Nkx2.2;Nkx2.9 double-null embryos. We show that neuronal progenitor cells within the p3 domain are not specified correctly and that putative V3 progenitors are entirely replaced by MN precursors. Likewise, cells of the lateral floor plate in double- Development 137 (24) Nkx2 transcription factors regulate floor plate development RESEARCH ARTICLE 4251 Anterograde labeling of commissural axons Embryos were collected at E12.5, fixed in 4% paraformaldehyde in PBS for 45 minutes then washed in PBS. Subsequently, spinal cords were dissected carefully, dorsal root ganglia were removed and the tubes were cut open at the dorsal midline (roof plate) to obtain ‘open-book’ preparations, as described previously (Perrin and Stoeckli, 2000). Following a second fixation step, DiI solution (5 mg/ml in methanol; Molecular Probes) was injected with finely drawn-out glass capillaries into the soma of dorsal commissural neurons (dl1 neurons) (Zou et al., 2000). DiI was allowed to penetrate axon trajectories in cold PBS for 40-48 hours. Analysis and documentation was performed on a standard fluorescence or confocal microscope. Fictive locomotion in isolated spinal cord preparations RESULTS V3 neuronal progenitor cells are replaced by Olig2+ MN precursors in the ventral neural tube of Nkx2.2–/–;Nkx2.9–/– double-mutant embryos In order to investigate potentially overlapping or redundant roles of the structurally related HD transcription factors Nkx2.2 and Nkx2.9, we generated double-mutant mice lacking both proteins. Crossing the single-knockout mutants (Pabst et al., 2003; Sussel et al., 1998) yielded the expected combinatorial genotypes including the Nkx2.2–/–;Nkx2.9–/– double-null mutants, which were born with no apparent anatomical defects. However, double-null mutants died shortly after birth, possibly as a result of respiratory problems or a lack of pancreatic islet cells, as described previously for the Nkx2.2–/– mutant mouse (Sussel et al., 1998). Because double-null mutant embryos developed to term, neither transcription factor, individually or in combination, can be vitally important during prenatal mouse development. Interestingly, a single Nkx2.2 wild-type allele in Nkx2.2+/–;Nkx2.9–/– compound Fig. 1. Early patterning defect in the ventral neural tube of Nkx2.2–/–;Nkx2.9–/– double-mutant mouse embryos: loss of the p3 progenitor domain. (A-L)Immunohistochemistry for neuronal progenitor cells in the ventral spinal cord of E9.5 (A-D) and E10.5 (E-L) mouse embryos at forelimb (E-H) and hindlimb (I-L) levels. Transverse sections of wild type (A,E,I,G,K), Nkx2.2–/– (B), Nkx2.9–/– (C) and Nkx2.2–/–;Nkx2.9–/– mutant (D,F,H,J,L) animals reveal unaltered expression of Pax6 (green in A-F,I,J) and Nkx6.1 (green in G,H,K,L) in single and double mutants whereas Olig2+ MN progenitor cells (blue) expand more ventrally into the p3 domain in the double mutant (D,F,H,J,L). Olig2+ MN precursors initially occupy the entire width of the neural tube and become restricted to the ventricular zone later. The ventral expansion of the MN progenitor domain in rostral sections of mutant embryos is highlighted by vertical lines (E-H). Scale bars: 50m. mutants was sufficient for postnatal survival. However, most of these animals displayed abnormal locomotion that manifested as a hopping gait. To test the hypothesis that functional redundancy exists between both genes, we analyzed the neuronal patterning in spinal cords of double-homozygous Nkx2.2–/–;Nkx2.9–/– mouse embryos using diagnostic markers, such as the transcription factors Olig2, Pax6 and Nkx6.1, which mark all neuronal progenitors ventral to the p2 domain (Sander et al., 2000). We found that at E9.5 the p3 domain in Nkx2.2 and Nkx2.9 single-knockout mutant mice was located ventral to MN progenitor cells forming the correct dorsal boundary with Olig2-expressing (Olig2+) cells as observed in wild-type embryos (Fig. 1A-C) (Mizuguchi et al., 2001; Novitch et al., 2001; Sun et al., 2001). By contrast, Olig2+ progenitors in double-null Nkx2.2–/–;Nkx2.9–/– embryos at E9.5 expanded into the ventral neural tube, including the p3 domain and, surprisingly, also a major part of the floor plate (Fig. 1D; Fig. 4M). This phenotype persisted in double-mutant embryos at E10.5 when Olig2 expression began to retract from the medial floor plate area (Fig. 1F,H). The ventral border of Pax6-expressing cells and the expression domain of DEVELOPMENT For in vitro recordings, neonatal pups from postnatal day (P) 0 to P2 were used except for one initial litter of E18.5 Nkx2.2 embryos. Nkx2.9–/–;Nkx2.2–/– double-knockout animals die at birth and were not included in this study. All electrophysiological experiments were carried out as described in detail elsewhere (Quinlan and Kiehn, 2007). Fictive locomotion was induced with a mixture of N-methyl-D-aspartic acid (NMDA; 3-7.5 M) and 5-hydroxytryptamine (5-HT; 3-20 M). In a few cases (n5) dopamine (20-50 M) was also added. For blocked inhibition experiments, 10 M picrotoxin and 0.3 M strychnine was applied. The left and right activities of the L2 and L5 ventral roots were recorded with suction electrodes. To estimate the level of coordination, ventral root recordings were analyzed from stable locomotor activity, taken at least 10 minutes after the initial burst of drug-induced activity using circular statistics as described previously (Kjaerulff and Kiehn, 1996; Tresch and Kiehn, 1999). In brief, raw traces of the ventral root activity were rectified and smoothed (moving average) using custom written applications in R software (http://www.r-project.org/). Cycle period onsets were automatically calculated based on the intersection between the smoothed tracing with a horizontal line running through the center of this trace. The cycle periods of the reference trace (LL2) were each normalized to a scale of 360 degrees, with the onset of a locomotor burst in the reference ventral root corresponding to a phase value of zero and the onset of the next burst corresponding to a phase value of 360. The phase delay of the period cycle onset with respect to the reference was calculated for test traces (LL5, RL2 or RL5). Based on these values, an estimate of ventral root coordination was made and locomotor cycles were selected randomly. From these values, a vector representing the level of individual animals was calculated. The angle of the vector represents the average phase delay with respect to the reference root, and the length is proportional to the clustering around this value. Raleigh’s test was used to determine the degree of clustering with statistical significance (P<0.05; illustrated as a blue circle). Of the 58 animals tested, only 32 were included in the final circular statistics because of poorly defined signals in the remaining preparations. 4252 RESEARCH ARTICLE Development 137 (24) Fig. 2. Development of V0, V1 and V2 neurons is unaffected in the ventral spinal cord of Nkx2.2–/–;Nkx2.9–/–and Nkx2.2+/–;Nkx2.9–/–mutant mouse embryos. (A,B)In situ hybridization using probes for Dbx1, En1, Chx10 and Sim1 on transverse sections of E11.5 wild-type, Nkx2.2+/–;Nkx2.9–/– and Nkx2.2–/–;Nkx2.9–/– mutant embryos. No major differences are observed in V0 (Dbx1), V1 (En1) and V2a (Chx10) neurons (A). Notably, Sim1-expressing V3 neurons are present in Nkx2.2+/–;Nkx2.9–/– mutant embryos of the presumptive hopping phenotype (B). (C)Immunofluorescent analysis using Lim3 (green) and Islet1 (red) antibodies on embryos at E12.5 illustrates an increase in the number of somatic motor neurons (yellow) within the medial motor column (MMC) of Nkx2.2+/–;Nkx2.9–/– and Nkx2.2–/–;Nkx2.9–/– mutants. Islet1-positive MN cells of the lateral motor column (LMC) and Lim3+ V2 progenitors (green in encircled area) appear unaffected in double-mutant embryos. DRG, dorsal root ganglia. (D) Quantification of V2 progenitor, MMC and LMC cells demonstrates a significant increase of motor neurons in the MMC in compound and double-null mutants versus wild type. Seven or more independent sections were counted at forelimb level from wild type (n3) and Nkx2.2–/–;Nkx2.9–/– (Nkx2ko; n3) mutant embryos. Data are expressed as mean ± s.d. Scale bar: 50m. Nkx2.9 is activated in the p3 domain, including the floor plate territory, of Nkx2.2–/–;Nkx2.9–/– double-null embryos The model for neuronal patterning in the ventral spinal cord proposes that progenitor cells in the p3 domain enter the lineage of V3 neurons driven by a high concentration of Shh signal and subsequent induction of the transcription factors Nkx2.2 and Nkx2.9 (Dessaud et al., 2007; Ericson et al., 1997b). No other specific markers have been described for these early cells. To confirm that DEVELOPMENT Nkx6.1 were unaffected in double-mutant embryos suggesting that both Nkx2.2 and Nkx2.9 were not essential for control of the expression pattern of those transcription factors (Fig. 1). Thus, molecular markers in the embryonic spinal cord of the Nkx2.2–/–;Nkx2.9–/– double mutant clearly indicated that V3 progenitor cells were absent and replaced entirely by MN precursor cells. This is in contrast to Nkx2.2 and Nkx2.9 single-mutant embryos, which contained V3 progenitor cells in the p3 domain, as observed in wild type (Fig. 1B,C) (Briscoe et al., 1999; Pabst et al., 2003). Taken together, these observations support the idea that Nkx2.2 and Nkx2.9 play redundant roles and that one or both factors is required and sufficient for the formation of the early p3 progenitor domain. In order to confirm that simultaneous depletion of Nkx2.2 and Nkx2.9 did not cause indirect or compensatory effects on the development of the other neuronal cell populations in ventral spinal cord, we performed in situ hybridization with specific probes for Dbx1, En1, Chx10 and Sim1 to identify V0, V1, V2a and V3 neurons, respectively. Our results indicated that the normal dorsoventral distribution, and presumably numbers, of V0, V1 and V2a interneurons were maintained in Nkx2.2–/–;Nkx2.9–/– doublenull embryos at E11.5 (Fig. 2A). Counts of Lim3+ and Islet1expressing cells in Nkx2.2–/–;Nkx2.9–/– embryos at E12.5 confirmed that motor neurons in the medial motor column (MMC) were specifically expanded at the expense of V3 cells (Fig. 2C,D). By contrast, numbers of Lim3+ and Islet1– V2 progenitor cells representing the next dorsal class of interneurons, and Islet1+ neurons of the lateral motor column (LMC) appeared to be unaffected in mutant embryos (Fig. 2C,D). This latter result confirms and expands previously published observations in the Nkx2.2–/– mutant, which demonstrated that ectopic MN progenitors in p3 contribute to the MMC only and not to the LMC (Agalliu et al., 2009). Notably, Nkx2.2+/–;Nkx2.9–/– compound-mutant embryos, most of which developed the hopping gait, also showed a moderate increase of MMC motor neurons (Fig. 2C,D) and a reduction, but not a complete loss, of mature Sim1+ V3 interneurons (Fig. 2B). This decrease was in line with fewer Nkx2.2-expressing V3 progenitor cells, which we identified by immunostaining in the Nkx2.2+/–;Nkx2.9–/– compound mutant (Fig. 4F,L). All other cardinal classes of interneurons in the ventral spinal cord of Nkx2.2+/–;Nkx2.9–/– mutant embryos were indistinguishable from those observed in wild-type animals (Fig. 2A). Fig. 3. The Nkx2.9-lacZ knock-in allele is expressed in the floor plate of E10.5 Nkx2.2;Nkx2.9-deficient embryos. -Galactosidase activity (blue) in whole-mount embryos (left panels) reflecting Nkx2.9 activity in the CNS and in transverse sections at the rostral trunk level (right panels) illustrating that Nkx2.9 is activated in the most ventral neural tube, including the floor plate. This expression domain is established independently of both Nkx2.2 and Nkx2.9 proteins. prospective V3 progenitors were actually generated but take another fate in Nkx2.2–/–;Nkx2.9–/– double-mutant embryos, we utilized the lacZ reporter gene inserted into the Nkx2.9 mutant allele (Pabst et al., 2003). In E10.5 Nkx2.9-deficient embryos, -galactosidase activity was observed throughout the normal Nkx2.9 expression domain regardless of whether or not Nkx2.2 was also present (Fig. 3). This finding suggests that initial Nkx2.9 gene activation is probably directly controlled by the Shh signal and is not subject to feedback control, such as auto- or cross-regulation by Nkx2.2 or Nkx2.9. More interestingly, -galactosidase-positive cells were not only found in the p3 domain but also more ventrally within the floor plate region, suggesting that Nkx2.9-expressing cells initially include prospective progenitors of neuronal cell lineages as well as non-neuronal floor plate cells. Typical floor plate markers are reduced in mutant embryos lacking Nkx2.2 and Nkx2.9 In an attempt to assess a function for Nkx2.2 and Nkx2.9 transcription factors in the development of the floor plate we first determined the expression of the forkhead transcription factor FoxA2, a key regulator for floor plate cells in the embryonic spinal cord (Ruiz i Altaba et al., 1995). Immunohistochemistry with a FoxA2-specific antibody on rostral (forelimb) and caudal (hindlimb) transverse sections of the spinal cord from E10.5 mouse embryos revealed almost normal expression in Nkx2.2–/– mutants, and a moderately smaller expression domain particularly on caudal sections of Nkx2.9–/– single mutants (Fig. 4G-J). In the Nkx2.2–/–;Nkx2.9–/– double knockout (Fig. 4C,D) and also in Nkx2.2+/–;Nkx2.9–/– compound-mutant mice (Fig. 4E,F), FoxA2expressing cells and/or the accumulation of FoxA2 protein were significantly reduced (Fig. 4A,B) and replaced by Olig2+ MN progenitor cells that were already present at E9.5 (Fig. 4K-M). Some cells in the ventral midline, however, retained FoxA2 suggesting that RESEARCH ARTICLE 4253 only the lateral and not the medial floor plate acquired an MN fate in the double-null mutant. A similar reduction of floor plate territory was also observed for Shh (data not shown). Incorporation of BrdU and immunostaining for active caspase 3 did not provide evidence for major changes in proliferation or enhanced apoptosis in ventral spinal cords of mutant embryos, supporting the idea of cell-fate conversion (see Fig. S1 in the supplementary material). Counting immunostained cells on multiple sections of the various mutant and wild-type embryos confirmed that floor plates in double-null and Nkx2.2+/–;Nkx2.9–/– compound mutants were significantly smaller than normal (Fig. 4K). Floor plates in both mutant genotypes appeared to be more affected at caudal than rostral levels. To provide further evidence of the mutant floor plate phenotype, we performed in situ hybridization with additional markers and extended the analysis to older embryos (E11.5). At E10.5 and E11.5 hybridization signals for Shh, netrin 1, FoxA2 and Slit1 transcripts were drastically diminished in double-null Nkx2.2–/–;Nkx2.9–/– (Fig. 4W-Z,II-LL) and compound Nkx2.2+/–;Nkx2.9–/– (Fig. 4S-V,EE-HH) mutant embryos in comparison with wild type (Fig. 4O-R,AA-DD). Taken together, these results provide compelling evidence that many cells, particularly in the lateral floor plate, were transformed into MN progenitors in double-null and compound Nkx2.2+/–;Nkx2.9–/– mutant embryos. Aberrant pathfinding of commissural axons in spinal cord of Nkx2.2;Nkx2.9 mutant embryos Besides its role as signaling center for dorsoventral patterning of the neural tube (Jessell, 2000), the floor plate also controls pathfinding of commissural axons (Charron et al., 2003; Matise et al., 1999; Serafini et al., 1994). The massive changes in floor plate morphology and/or specific gene expression in Nkx2.2;Nkx2.9 mutant embryos prompted us to monitor axon guidance by anterograde labeling of neurons in open-book preparations of the neural tube from E12.5 embryos, as described previously (Bourikas et al., 2005). DiI injection into dorsally located somata in spinal cord preparations from wild-type embryos revealed straight and parallel bundles of commissural axons that projected towards the floor plate, crossed the midline and turned rostrally, as expected (Fig. 5A). Although commissural axons in spinal cords of most Nkx2.2–/– and Nkx2.9–/– single-knockout embryos displayed normal projections, 10-20% of these mutants appeared to form less tightly packed axon bundles that often stalled at the midline without reaching the contralateral side (Fig. 5B,C). The most frequent pathfinding errors of commissural axons were detected in spinal cords isolated from Nkx2.2–/–;Nkx2.9–/– double-null mutants and Nkx2.2+/–;Nkx2.9–/– mutants that develop the hopping phenotype (Fig. 5D-F). Approximately 50% of DiI injection sites in doublehomozygous and compound mutants revealed highly abnormal axons with premature ipsilateral turning, random rostral and caudal turning after crossing the floor plate, and increased density of growth cones indicating stalling at the midline (Fig. 5E). Generally, deviations in axon projections appeared to be most complex in double-null mutants, although the numbers of injection sites displaying abnormal axon projections were similar in compound Nkx2.2+/–;Nkx2.9–/– mutants (Fig. 5G). Potential defects in DV pathfinding or fasciculation of axons prior to reaching the floor plate were analyzed on transverse sections of DiI-injected spinal cord preparations. The results confirmed aberrant axonal navigation at the ventral midline and showed that the vast majority of axons reached the ipsilateral floor plate border correctly, even in mice lacking both Nkx2.2 and Nkx2.9 (Fig. 5K,L). This observation was supported by immunofluorescent labeling of transverse DEVELOPMENT Nkx2 transcription factors regulate floor plate development 4254 RESEARCH ARTICLE Development 137 (24) spinal cord sections from wild-type, Nkx2.2+/–;Nkx2.9–/– and Nkx2.2–/–;Nkx2.9–/– embryos using Rig1- (Tamada et al., 2008) and Tag1-specific (Dodd et al., 1988) antibodies (Fig. 5H-J). We found no evidence for any major deviations of dorsoventral axon trajectories before midline crossing. In summary, our data demonstrate severe defects in pathfinding of commissural axons at the ventral midline of the spinal cord in mice that lack one or both of the transcription factors Nkx2.2 and Nkx2.9. The aberrant axonal navigation and the reduced expression of the axonal guidance molecules Shh, netrin 1 and Slit1 in mutant floor plates support the model that correct formation of the ventral commissure requires Nkx.2.2 and/or Nkx2.9 activity. The adult Nkx2.2+/–;Nkx2.9–/– mutant mouse exhibits abnormal locomotor activity and coordination defects during walking Nkx2.2+/–;Nkx2.9–/– mutant mice with one wild-type Nkx2.2 gene copy were viable but ~90% of these compound mutants displayed abnormal patterns of locomotor coordination (Fig. 6A-D). The gait DEVELOPMENT Fig. 4. Reduction of the floor plate in Nkx2.2+/–;Nkx2.9–/– and Nkx2.2–/–;Nkx2.9–/– double-null embryos. (A-J)Immunofluorescence for Nkx2.2 (red) and the floor plate marker FoxA2 (green) on rostral (forelimb; A,C,E,G,I) and caudal (hindlimb; B,D,F,H,J) transverse sections of E10.5 mouse embryos reveals a significantly smaller FoxA2-positive floor plate in Nkx2.2–/–;Nkx2.9–/– (C,D) and Nkx2.2+/–;Nkx2.9–/– (E,F) mutant embryos compared with wild type (A,B) and Nkx2.9–/– (G,H) and Nkx2.2–/– (I,J) single mutants. (K-M)Triple immunofluorescence at E9.5 demonstrates that all of the p3 domain and a major portion of the floor plate were replaced by Olig2-positive (blue) MN progenitors in the Nkx2.2–/–;Nkx2.9–/– mutant. (N)FoxA2-expressing nuclei were counted on 57 (wild type), 14 (Nkx2.2–/–), 30 (Nkx2.9–/–), 23 (Nkx2.2+/–;Nkx2.9–/–) and 43 (Nkx2.2–/–;Nkx2.9–/–) individual sections of three to five embryos per genotype. Note the stronger effect in the caudal spinal cord. Data are expressed as mean ± s.d. (O-LL) In situ hybridization using specific probes for Shh, netrin 1, FoxA2 and Slit1 demonstrate the dramatic loss of these markers in Nkx2.2+/–;Nkx2.9–/– and Nkx2.2–/–;Nkx2.9–/– mouse embryos at E10.5 (O-Z) and E11.5 (AA-LL). Scale bars: 50m. Nkx2 transcription factors regulate floor plate development RESEARCH ARTICLE 4255 Fig. 6. Adult Nkx2.2+/–;Nkx2.9–/– animals display a hopping gait. (A-C)The hopping phenotype of adult mutant mice is illustrated for two animals with either strong (A) or weak (B) deviations from normal locomotor behavior (C). To demonstrate foot prints on a sheet of paper, fore and hind paws were painted with black and red ink, respectively. Tracings are depicted by foot symbols of matching color. Synchronous activation of the hind limbs is highlighted by the blue bars. Locomotor coordination appears to be affected more strongly in hind than fore limbs, and animals can change between normal alternating and hopping locomotion. Occasionally, extra forelimb footsteps disrupt the regular step pattern (arrowhead in A). (D)Percentage of animals with strong and weak gait phenotypes for the available mutant genotypes at the age of 3-6 months. Nkx2.9+/–, n35; Nkx2.2+/–;Nkx2.9+/–, n50; Nkx2.9–/–, n121; Nkx2.2+/–;Nkx2.9–/–, n171. Locomotor-like activity in isolated spinal cords from newborn Nkx2.2+/–;Nkx2.9–/– mutant mice The abnormal motor behavior of adult Nkx2.2+/–;Nkx2.9–/– mutant mice might reflect defects in spinal neuronal networks, the CPGs generating locomotion. The elements of the mammalian CPGs are embedded in the ventral half of the spinal cord where Nkx2.2 and Nkx2.9 transcription factors are expressed and constitute key developmental regulators for V3 neurons (Briscoe et al., 1999; Pabst DEVELOPMENT Fig. 5. Pathfinding of commissural axons in spinal cords of Nkx2.2 and Nkx2.9 mutant mice is significantly impaired at the ventral midline. (A-F)Confocal images of DiI-labeled commissural axons in open-book explants of wild type (WT; A), Nkx2.2–/– (B) and Nkx2.9–/– (C) single mutants, Nkx2.2+/–;Nkx2.9–/– (D), and Nkx2.2–/–;Nkx2.9–/– (E,F) double mutants. In all panels the anterior end of the spinal cord is at the top. Dotted lines demarcate the floor plate. The axons in the wild type cross the floor plate and turn anteriorly (A), whereas axons in mutants frequently stall at the floor plate, turn randomly before or after the midline, or fail to turn at all (B-F). (G)Quantification of aberrant injection sites for each mutant genotype relative to wild type. DiI injection sites were scored as showing aberrant pathfinding when at least 25% of the labeled axons did not cross the midline, when more than 50% of the axons did not turn into and extend along the longitudinal axis of the spinal cord, or when axons turned into the longitudinal axis prematurely in the floor plate. Data are expressed as mean ± s.d. (H-J)Normal DV trajectories of commissural axons in double mutants is illustrated by immunofluorescence with Rig1 (green) and Tag1 (red) antibodies. (K,L)Transverse sections of DiI-injected spinal cords illustrate regular DV projections until axons reach the ipsilateral floor plate border in Nkx2.2+/–;Nkx2.9–/– (K) and Nkx2.2–/–;Nkx2.9–/– (L) mutant mice. Horizontal and vertical dashed lines indicate the midline and ventral border of the spinal cord, respectively. Insets show lower magnification of DiI-injected sites. ML, midline. abnormalities were observed predominantly for hindlimbs as continuous hopping with no alternating activity of left and right legs, or as less frequent, intermittent hopping. To illustrate the locomotor phenotype, fore- and hindpaws were painted with black and red ink, respectively, and color-coded footprints were recorded on a sheet of paper. More than 60% of the Nkx2.2+/–;Nkx2.9–/– mutants showed severe locomotion defects of hindlimbs and less stereotypic movements of forelimbs, including extra steps between the normal step cycle (Fig. 6A). Approximately 30% of mutants displayed occasional hopping with only a few interspersed episodes of abnormal leg movements (Fig. 6B). These locomotor phenotypes were confirmed for a large number of mice (more than 70 animals for each genotype) and were never observed in wild type or heterozygous single and double mutants. However, in ~5% and 20% of homozygous Nkx2.9 single mutants we also observed the strong and weak hopping phenotypes, respectively (Fig. 6D). 4256 RESEARCH ARTICLE Development 137 (24) et al., 1998; Pabst et al., 2003) and floor plate, as shown here. In order to determine the functional contribution of Nkx2.2 and Nkx2.9 to the development of the locomotor CPG in addition to the maturation of the V3 population, we performed in vitro recordings in spinal cord preparations from newborn mutant mice carrying various allelic combinations of Nkx2.2 and Nkx2.9 targeted gene disruptions. Locomotor-like activity was induced by bath application of NMDA and 5-HT. Because it was generally difficult to induce fictive locomotion in spinal cords from Nkx2.9-deficient animals, dopamine (20-50 M) was added in some of these experiments (n5). Electrical activities of ventral roots on both sides of the spinal cord were recorded at lumbar level L2 to evaluate left-right coordination. Activities of the ipsilateral roots at L2 and the more caudal L5 were also measured to assess the pattern of motor activity for flexor (L2) and extensor (L5) muscles. Examples of ventral root recordings from Nkx2.2–/– and Nkx2.2+/–;Nkx2.9–/– compoundmutant spinal cords are shown in Fig. 7A and Fig. 7B, respectively. Average circular plots of left-right (LL2-RL2) and flexor-extensor (iL2-iL5) coordination for homozygous and heterozygous Nkx2.2 mutant animals with normal Nkx2.9 wild-type genes revealed no systematic change in the drug-induced locomotor-like activity (Fig. 7C) compared with wild-type animals (Fig. 7A; data not shown). These results suggest that blocking the formation of V3 neurons during embryonic mouse development causes no significant effect on left-right and flexor-extensor coordination. By contrast, Nkx2.9–/– animals and, most prominently, the Nkx2.2+/–;Nkx2.9–/– compound mutants showed markedly reduced coordination of drug-induced locomotor-like activity with increased variability in both left-right and flexor-extensor coordination, as illustrated by the circular plots (Fig. 7D). Unfortunately, double-homozygous Nkx2.2–/–;Nkx2.9–/– null mutants could not be included in the analysis because of their early postpartum lethality. The observed increase in variance of locomotor-like activity was mainly attributable to two phenomena. First, left-right coordination in these animals often drifted in phase DEVELOPMENT Fig. 7. Nkx2.9-deficient mice display defects of left-right and flexor-extensor coordination during locomotor activity in the isolated spinal cord. (A,B)Representative raw traces of ventral root recordings during drug-induced locomotor activity in spinal cords of Nkx2.2–/– (A) and Nkx2.2+/–;Nkx2.9–/– (B) newborn mice. Lower panel in B shows traces from the boxed area with higher resolution of the time axis. Activities were recorded in ventral roots from left (L) and right (R) lumbar segments L2 (LL2, RL2, respectively; flexor-dominated), and the left lumbar segment L5 (LL5; extensor-dominated). Each locomotor cycle is indicated in reference to LL2 by alternating grey and white background. The onset of locomotor cycles at RL2 and LL5 are shown as raster plots above the corresponding recordings. The superimposed thin red line shows the rectified smoothed ventral root activity. (C,D)Circular phase diagrams of left-right (L-R) and intersegmental flexor-extensor (iL2-iL5) coordination show no significant uncoupling in Nkx2.2+/– (green arrow in C; n5) and Nkx2.2–/– (blue arrow in C; n2) pups. By contrast, a number of Nkx2.9-deficient animals, both Nkx2.9–/– single mutants (red arrow in D; n6) and Nkx2.2+/–;Nkx2.9–/– compound mutants (black arrow in D; n11), exhibited insignificant coupling between left-right and flexor-extensor ventral root activities (dots falling within the blue circle). One Nkx2.2+/–;Nkx2.9–/– double mutant showed hopping with phase values around zero. Each dot in the graphs represents the average phase value calculated from 30 locomotor cycles for each individual animal. The blue circle marks the significance level with P<0.05 outside the blue circle. (E)Representative recording from an isolated spinal cord of an Nkx2.2+/–;Nkx2.9–/– pup. Fast GABAergic and glycinergic synaptic transmission was blocked with picrotoxin (10M) and strychnine (0.3M), respectively. Spontaneous slow synchronous bursts of activity occur in all ventral roots. RESEARCH ARTICLE 4257 between the locomotor cycles on each side of the spinal cord leading to a relaxed coordination that lacked clear segmental alternation. Second, a double burst activity was sometimes seen in L5 (seen as two bursts per L2 cycle, one in and one out of phase with L2; Fig. 7B). Thus, it seemed that the regular out-of-phase flexor-extensor activity was maintained but complemented with additional activation of the L5 ventral root in synchrony with the ipsilateral rostral L2 root in these mutants. In summary, Nkx2.9–/– mice and, even more significantly, the Nkx2.2+/–;Nkx2.9–/– mutants displayed a high degree of uncoupling of left and right side activity and aberrant flexor-related bursting in the otherwise extensor-dominated L5 root. Seventy-five percent of V3 cells form a population of excitatory commissural neurons (CINs) (Zhang et al., 2008). This suggests that crossed excitatory drive might be reduced in Nkx2.2+/–;Nkx2.9–/– compound mutants. Therefore, we examined whether these animals were still able to coordinate left-right activities using the remaining excitatory CINs. To test this, we recorded locomotor-like activity in the presence of picrotoxin and strychnine, which block the two main classes of inhibitory receptors, -aminobutyric acid (GABA) and glycine receptors, respectively. In spinal cords of wild-type mice, these experimental conditions led to synchronous activities in all ventral roots (Hinckley et al., 2005). The same result was obtained with spinal cords from the Nkx2.2+/–;Nkx2.9–/– mutant (Fig. 7E). Taken together, the electrophysiological data indicate that developmental loss of V3 neurons alone probably has little impact on druginduced locomotor-like activity, whereas complete disruption of the Nkx2.9 gene and partial loss of Nkx2.2 results in substantial locomotor deficits, both in vitro and in vivo. shown that Nkx2.2 is able to repress the expression of Olig2 (Novitch et al., 2001). The ectopic expression of Olig2 in Nkx2.2 and Nkx2.9-depleted cells supports the idea of transcriptional repression of MN lineage-specific genes, such as Olig2. In this scenario, formation of motor neurons would be driven by Shh signaling through the direct induction of Olig2 (Dessaud et al., 2007). Support for this view also comes from the temporal sequence of marker gene expression in the early ventral neural tube, in which the basic progenitor organization is probably already set by E8.5-9.0 by Shh signaling from the notochord (Ericson et al., 1996; Jeong and McMahon, 2005). Olig2 expression is first seen in the most ventral part of the early neural tube and precedes that of Nkx2.2 in an area that soon after will acquire floor plate identity. Olig2 expression is probably repressed by Nkx2.2 and/or Nkx2.9 in early p3 progenitors and, as we show here, in prospective floor plate cells. Nkx2.2 has also been suggested to repress the expression of Pax6 in p3 cells (Briscoe et al., 2000). However, we find no evidence for a ventrally expanded Pax6 domain in the spinal cord of double-null mutant embryos, in agreement with previous observations for the homozygous Nkx2.2 mutant (Briscoe et al., 1999). In conclusion, we propose a model in which Nkx2.2 and Nkx2.9 genes collaborate to serve a dual function during development of V3 interneurons. First, the repressive activity of Nkx2.9 and/or Nkx2.2 leads to the determination of the early V3 progenitor cell lineage by preventing Olig2 expression, as shown here. Second, continued activity of, presumably, Nkx2.2 is essential to mediate differentiation of progenitors into Sim1+ V3 interneurons (Briscoe et al., 1999). DISCUSSION Individual targeted disruptions of the genes encoding the related HD transcription factors Nkx2.2 (Briscoe et al., 1999; Sussel et al., 1998) and Nkx2.9 (Pabst et al., 2003) suggested that both proteins play essential and partially redundant roles for the development of distinct neuronal populations in hindbrain and ventral spinal cord. Here, we report three novel findings in double-mutant mice. First, V3 neural progenitors cells are converted to the MN cell lineage in the spinal cord of mouse embryos that lack both transcription factors. Second, development of a structurally and functionally intact floor plate is impaired in the absence of Nkx2.2 and Nkx2.9, which might cause the observed severe axonal pathfinding defects of commissural neurons. Third, the majority of adult Nkx2.2+/–;Nkx2.9–/– mutant mice exhibits abnormal locomotor behavior, such as a hopping gait and increased variability of leftright and flexor-extensor coordination in drug-induced locomotorlike activity of isolated spinal cords. The transcription factors Nkx2.2 and Nkx2.9 are essential for normal locomotor behavior Unexpectedly, we observed that the majority of adult compound Nkx2.2+/–;Nkx2.9–/– mutant mice exhibited a strong walking impairment. This phenotype was supplemented by the finding that neonatal mutant mice of the Nkx2.9-null genotype showed substantially abnormal drug-induced locomotor-like activity, which was even more pronounced in Nkx2.2+/–;Nkx2.9–/– mutant animals. Our data therefore indicate the importance of Nkx2.9 and Nkx2.2 factors for a fully functional CPG network. In vitro locomotor experiments revealed that compound Nkx2.2+/–;Nkx2.9–/– mutant mice possibly have more than one defect in locomotion, as leftright and flexor-extensor motor activity patterns were disturbed. Mutant animals displayed alterations of the interplay between L2 and L5 ventral roots. Although the normal out-of-phase activity of the ipsilateral L2 and L5 segments seemed to be preserved in mutant animals, it was complemented by an additional activation of L5 in synchrony with the ipsilateral L2 activation. This extensorrelated double burst presents a CPG phenotype not previously described for genetic perturbation experiments in any of the V0-V3 groups. In normal animals, the alternation of motor bursts on both sides of the spinal cord is coordinated through the activity of CINs. Proper left-right alternation is fundamental for coordinated walking behavior in mammals, which is thought to be mediated through inhibitory and excitatory pathways (Butt and Kiehn, 2003; Jankowska, 2008; Kiehn, 2006; Quinlan and Kiehn, 2007). Support for this view was obtained by the analysis of a mouse mutant in which the V0 class was perturbed by genetically ablated Dbx1, the fate-determining transcription factor in V0 neurons. This gene mutation caused disturbances in left-right coordination in the spinal cord (Lanuza et al., 2004). Rhythmic coupling of left-right motor Cell fate determination of neuronal progenitors in the p3 domain requires either Nkx2.2 or Nkx2.9 The present study provides evidence that neuronal progenitor cells in the p3 domain acquire a more dorsal fate in the absence of Nkx2.2 and Nkx2.9. This observation, together with previous studies, clearly indicates that at least one of the two transcription factors is required and sufficient for correct cell lineage determination in this part of the spinal cord, supporting the model of functional redundancy between both genes. In the double-null mutant, but not in single-mutant or wild-type embryos, cells in the p3 region express Olig2, which identifies them as prospective MN precursors that replace the normal V3 progenitors. Recent lineage tracing studies demonstrated that Nkx2.2 and Olig2 cell lineages share a common origin (Dessaud et al., 2007). Furthermore, it was DEVELOPMENT Nkx2 transcription factors regulate floor plate development bursts seems to be coordinated by ipsilaterally projecting interneurons, as genetic depletion for excitatory Chx10-positive V2a neurons also disrupted left-right alternation (Crone et al., 2008; Crone et al., 2009). V2a interneurons synapse (among other targets) on V0 commissural interneurons (Crone et al., 2008) emphasizing the complexity of the neuronal network that is required to drive the left-right alternation. Mice with a conditional block of neurotransmission in all V3derived interneurons have been generated recently by selective expression of the tetanus toxin light chain subunit (Zhang et al., 2008). The regularity and robustness of the locomotor rhythm in these mutant mice was severely perturbed, particularly the duration of individual motor bursts, and the entire step cycle period of fictive locomotion appeared to be highly variable (Zhang et al., 2008). This is in contrast to wild-type mice and even mutants with complete loss of left-right alternation, in which the length of motor bursts per segment on either side of the spinal cord were identical and well coordinated (Fawcett et al., 2007; Kullander et al., 2003; Wegmeyer et al., 2007). Current knowledge suggests that V3derived neurons ensure normal walking gait by controlling and balancing the rhythmic motor outputs of independent locomotor centers on each side of the spinal cord as well as flexor-extensorrelated motor bursts. However, the Sim1+ neurons seem to play a minor role in the general left-right alternation of the locomotor cycle and flexor-extensor coordination (Zhang et al., 2008). Interestingly, in the present study fictive locomotion in spinal cords from Nkx2.2-deficient neonates that lack V3 neurons was not significantly different from wild type, suggesting that compensatory mechanisms during prenatal development of the CPG circuitry might obscure the phenotype. Because Nkx2.2deficient mice also contain higher numbers of MNs, it seems unlikely that an increased pool of MNs contributes significantly to the locomotor phenotype. Development of the floor plate and formation of the spinal commissure are defective in Nkx2.2;Nkx2.9 mutant mice Nkx2.2+/–;Nkx2.9–/– double-mutant and to a lesser extent Nkx2.9 single-mutant mice show clear defects in locomotor activity both in vitro and in vivo, although they generate all cardinal classes of neurons in the ventral neural tube, including some Sim1+ V3 neurons (Fig. 2) (Pabst et al., 2003). How then can the abnormal locomotor phenotype be explained? Nkx2.2 and Nkx2.9 are transiently expressed in cells that will contribute to the floor plate (Chamberlain et al., 2008; Jeong and McMahon, 2005). We demonstrate here that targeted disruption of both genes has a major impact on floor plate development most profoundly in Nkx2.2–/–;Nkx2.9–/– mutants but also in Nkx2.2+/–;Nkx2.9–/– animals (Fig. 4). Several known guidance molecules for commissural axons (Evans and Bashaw, 2010) showed reduced expression in mutants. Specifically, chemoattractants and repellents, like Shh (Charron et al., 2003), netrin (Kennedy et al., 1994; Kennedy et al., 2006; Serafini et al., 1994), and Slit (Hammond et al., 2005; Long et al., 2004) were significantly diminished in the floor plate of double mutants. Tracing of the trajectories of dorsal commissural neurons (presumably dl1 neurons) in open-book preparations of mutant spinal cords provided evidence of disturbed navigation of axons (Fig. 5), which normally extend towards the ventral midline attracted by guidance cues derived from the floor plate (Salinas, 2003). As axons contact the floor plate, they become unresponsive to these attractive signals and gain sensitivity to repellent molecules, which allows complete crossing of the midline and subsequent turning in an anterior direction (Stoeckli, Development 137 (24) 2006). Shh in the floor plate serves a dual function by first attracting commissural axons and then inducing the switch to repulsion that is important to permit entry into the contralateral hemisphere of the neural tube (Parra and Zou, 2010). Shh is reduced in the floor plate of the Nkx2 mutants, which might partly explain the aberrant pathfinding behavior of commissural axons observed in this study. As defects in axonal guidance and the locomotor phenotype appear to be correlated in Nkx2.2+/–;Nkx2.9–/– mutant mice, it is not unreasonable to propose that altered guidance cues for commissural axons could lead to wiring problems in the CPG circuitry and cause the abnormal locomotor behavior of these mutants. Although we have not yet tested which type of commissural neurons might be affected in the hopping mutant, V0 and dl6 interneurons are good candidates (Dougherty and Kiehn, 2010; Goulding, 2009; Kiehn et al., 2010). The reduction of V3 interneurons alone does not provide a sufficient explanation for the locomotor phenotype, as they are not required for left-right alternation (Zhang et al., 2008). Nevertheless, it cannot be ruled out entirely that subtle changes in the function of mutant V3 neurons contribute to the defective gait. Acknowledgements We thank Iris Linde for expert technical assistance, R. Hänsch for advice on confocal microscopy, S. Schildt and K. Sawinski for experimental help, and Dr Fujio Murakami for the Rig1 antibody. This work was supported by a grant of the Deutsche Forschungsgemeinschaft to H.H.A. and A.H., and grants from the Swedish Research Council and NIH (R01NS0407909) to O.K. Deposited in PMC for release after 12 months. Competing interests statement The authors declare no competing financial interests. Supplementary material Supplementary material for this article is available at http://dev.biologists.org/lookup/suppl/doi:10.1242/dev.053819/-/DC1 References Agalliu, D., Takada, S., Agalliu, I., McMahon, A. P. and Jessell, T. M. (2009). Motor neurons with axial muscle projections specified by Wnt4/5 signaling. Neuron 61, 708-720. Al-Mosawie, A., Wilson, J. M. and Brownstone, R. M. (2007). Heterogeneity of V2-derived interneurons in the adult mouse spinal cord. Eur. J. Neurosci. 26, 3003-3015. Ang, S. L., Wierda, A., Wong, D., Stevens, K. A., Cascio, S., Rossant, J. and Zaret, K. S. (1993). The formation and maintenance of the definitive endoderm lineage in the mouse: involvement of HNF3/forkhead proteins. Development 119, 1301-1315. Bourikas, D., Pekarik, V., Baeriswyl, T., Grunditz, A., Sadhu, R., Nardo, M. and Stoeckli, E. T. (2005). Sonic hedgehog guides commissural axons along the longitudinal axis of the spinal cord. Nat. Neurosci. 8, 297-304. Briscoe, J. and Ericson, J. (2001). Specification of neuronal fates in the ventral neural tube. Curr. Opin. Neurobiol. 11, 43-49. Briscoe, J. and Novitch, B. G. (2008). Regulatory pathways linking progenitor patterning, cell fates and neurogenesis in the ventral neural tube. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 363, 57-70. Briscoe, J., Sussel, L., Serup, P., Hartigan-O’Connor, D., Jessell, T. M., Rubenstein, J. L. and Ericson, J. (1999). Homeobox gene Nkx2.2 and specification of neuronal identity by graded Sonic hedgehog signalling. Nature 398, 622-627. Briscoe, J., Pierani, A., Jessell, T. M. and Ericson, J. (2000). A homeodomain protein code specifies progenitor cell identity and neuronal fate in the ventral neural tube. Cell 101, 435-445. Butt, S. J. and Kiehn, O. (2003). Functional identification of interneurons responsible for left-right coordination of hindlimbs in mammals. Neuron 38, 953-963. Chamberlain, C. E., Jeong, J., Guo, C., Allen, B. L. and McMahon, A. P. (2008). Notochord-derived Shh concentrates in close association with the apically positioned basal body in neural target cells and forms a dynamic gradient during neural patterning. Development 135, 1097-1106. Chapman, S. C., Schubert, F. R., Schoenwolf, G. C. and Lumsden, A. (2002). Analysis of spatial and temporal gene expression patterns in blastula and gastrula stage chick embryos. Dev. Biol. 245, 187-199. Charron, F., Stein, E., Jeong, J., McMahon, A. P. and Tessier-Lavigne, M. (2003). The morphogen sonic hedgehog is an axonal chemoattractant that collaborates with netrin-1 in midline axon guidance. Cell 113, 11-23. DEVELOPMENT 4258 RESEARCH ARTICLE Chiang, C., Litingtung, Y., Lee, E., Young, K. E., Corden, J. L., Westphal, H. and Beachy, P. A. (1996). Cyclopia and defective axial patterning in mice lacking Sonic hedgehog gene function. Nature 383, 407-413. Crone, S. A., Quinlan, K. A., Zagoraiou, L., Droho, S., Restrepo, C. E., Lundfald, L., Endo, T., Setlak, J., Jessell, T. M., Kiehn, O. et al. (2008). Genetic ablation of V2a ipsilateral interneurons disrupts left-right locomotor coordination in mammalian spinal cord. Neuron 60, 70-83. Crone, S. A., Zhong, G., Harris-Warrick, R. and Sharma, K. (2009). In mice lacking V2a interneurons, gait depends on speed of locomotion. J. Neurosci. 29, 7098-7109. Dessaud, E., Yang, L. L., Hill, K., Cox, B., Ulloa, F., Ribeiro, A., Mynett, A., Novitch, B. G. and Briscoe, J. (2007). Interpretation of the sonic hedgehog morphogen gradient by a temporal adaptation mechanism. Nature 450, 717720. Dodd, J., Morton, S. B., Karagogeos, D., Yamamoto, M. and Jessell, T. M. (1988). Spatial regulation of axonal glycoprotein expression on subsets of embryonic spinal neurons. Neuron 1, 105-116. Dougherty, K. J. and Kiehn, O. (2010). Functional organization of V2a-related locomotor circuits in the rodent spinal cord. Ann. New York Acad. Sci. 1198, 8593. Ericson, J., Morton, S., Kawakami, A., Roelink, H. and Jessell, T. M. (1996). Two critical periods of Sonic Hedgehog signaling required for the specification of motor neuron identity. Cell 87, 661-673. Ericson, J., Briscoe, J., Rashbass, P., van Heyningen, V. and Jessell, T. M. (1997a). Graded sonic hedgehog signaling and the specification of cell fate in the ventral neural tube. Cold Spring Harbor Symp. Quant. Biol. 62, 451-466. Ericson, J., Rashbass, P., Schedl, A., Brenner-Morton, S., Kawakami, A., van Heyningen, V., Jessell, T. M. and Briscoe, J. (1997b). Pax6 controls progenitor cell identity and neuronal fate in response to graded Shh signaling. Cell 90, 169180. Evans, T. A. and Bashaw, G. J. (2010). Axon guidance at the midline: of mice and flies. Curr. Opin. Neurobiol. 20, 79-85. Fawcett, J. P., Georgiou, J., Ruston, J., Bladt, F., Sherman, A., Warner, N., Saab, B. J., Scott, R., Roder, J. C. and Pawson, T. (2007). Nck adaptor proteins control the organization of neuronal circuits important for walking. Proc. Natl. Acad. Sci. USA 104, 20973-20978. Goulding, M. (2009). Circuits controlling vertebrate locomotion: moving in a new direction. Nat. Rev. Neurosci. 10, 507-518. Goulding, M. and Pfaff, S. L. (2005). Development of circuits that generate simple rhythmic behaviors in vertebrates. Curr. Opin. Neurobiol. 15, 14-20. Hammond, R., Vivancos, V., Naeem, A., Chilton, J., Mambetisaeva, E., Andrews, W., Sundaresan, V. and Guthrie, S. (2005). Slit-mediated repulsion is a key regulator of motor axon pathfinding in the hindbrain. Development 132, 4483-4495. Hinckley, C., Seebach, B. and Ziskind-Conhaim, L. (2005). Distinct roles of glycinergic and GABAergic inhibition in coordinating locomotor-like rhythms in the neonatal mouse spinal cord. Neuroscience 131, 745-758. Jankowska, E. (2008). Spinal interneuronal networks in the cat: elementary components. Brain Res. Rev. 57, 46-55. Jeong, J. and McMahon, A. P. (2005). Growth and pattern of the mammalian neural tube are governed by partially overlapping feedback activities of the hedgehog antagonists patched 1 and Hhip1. Development 132, 143-154. Jessell, T. M. (2000). Neuronal specification in the spinal cord: inductive signals and transcriptional codes. Nat. Rev. Genet. 1, 20-29. Kennedy, T. E., Serafini, T., de la Torre, J. R. and Tessier-Lavigne, M. (1994). Netrins are diffusible chemotropic factors for commissural axons in the embryonic spinal cord. Cell 78, 425-435. Kennedy, T. E., Wang, H., Marshall, W. and Tessier-Lavigne, M. (2006). Axon guidance by diffusible chemoattractants: a gradient of netrin protein in the developing spinal cord. J. Neurosci. 26, 8866-8874. Kiehn, O. (2006). Locomotor circuits in the mammalian spinal cord. Annu. Rev. Neurosci. 29, 279-306. Kiehn, O. and Kjaerulff, O. (1998). Distribution of central pattern generators for rhythmic motor outputs in the spinal cord of limbed vertebrates. Ann. New York Acad. Sci. 860, 110-129. Kiehn, O., Dougherty, K. J., Hagglund, M., Borgius, L., Talpalar, A. and Restrepo, C. E. (2010). Probing spinal circuits controlling walking in mammals. Biochem. Biophys. Res. Commun. 396, 11-18. Kjaerulff, O. and Kiehn, O. (1996). Distribution of networks generating and coordinating locomotor activity in the neonatal rat spinal cord in vitro: a lesion study. J. Neurosci. 16, 5777-5794. Kullander, K., Butt, S. J., Lebret, J. M., Lundfald, L., Restrepo, C. E., Rydstrom, A., Klein, R. and Kiehn, O. (2003). Role of EphA4 and EphrinB3 in local neuronal circuits that control walking. Science 299, 1889-1892. Lanuza, G. M., Gosgnach, S., Pierani, A., Jessell, T. M. and Goulding, M. (2004). Genetic identification of spinal interneurons that coordinate left-right locomotor activity necessary for walking movements. Neuron 42, 375-386. Long, H., Sabatier, C., Ma, L., Plump, A., Yuan, W., Ornitz, D. M., Tamada, A., Murakami, F., Goodman, C. S. and Tessier-Lavigne, M. (2004). Conserved RESEARCH ARTICLE 4259 roles for Slit and Robo proteins in midline commissural axon guidance. Neuron 42, 213-223. Lundfald, L., Restrepo, C. E., Butt, S. J., Peng, C. Y., Droho, S., Endo, T., Zeilhofer, H. U., Sharma, K. and Kiehn, O. (2007). Phenotype of V2-derived interneurons and their relationship to the axon guidance molecule EphA4 in the developing mouse spinal cord. Eur. J. Neurosci. 26, 2989-3002. Marti, E., Bumcrot, D. A., Takada, R. and McMahon, A. P. (1995). Requirement of 19K form of Sonic hedgehog for induction of distinct ventral cell types in CNS explants. Nature 375, 322-325. Matise, M. P., Lustig, M., Sakurai, T., Grumet, M. and Joyner, A. L. (1999). Ventral midline cells are required for the local control of commissural axon guidance in the mouse spinal cord. Development 126, 3649-3659. Mizuguchi, R., Sugimori, M., Takebayashi, H., Kosako, H., Nagao, M., Yoshida, S., Nabeshima, Y., Shimamura, K. and Nakafuku, M. (2001). Combinatorial roles of olig2 and neurogenin2 in the coordinated induction of pan-neuronal and subtype-specific properties of motoneurons. Neuron 31, 757771. Novitch, B. G., Chen, A. I. and Jessell, T. M. (2001). Coordinate regulation of motor neuron subtype identity and pan-neuronal properties by the bHLH repressor Olig2. Neuron 31, 773-789. Pabst, O., Herbrand, H. and Arnold, H. H. (1998). Nkx2-9 is a novel homeobox transcription factor which demarcates ventral domains in the developing mouse CNS. Mech. Dev. 73, 85-93. Pabst, O., Herbrand, H., Takuma, N. and Arnold, H. H. (2000). NKX2 gene expression in neuroectoderm but not in mesendodermally derived structures depends on sonic hedgehog in mouse embryos. Dev. Genes Evol. 210, 47-50. Pabst, O., Rummelies, J., Winter, B. and Arnold, H. H. (2003). Targeted disruption of the homeobox gene Nkx2.9 reveals a role in development of the spinal accessory nerve. Development 130, 1193-1202. Parra, L. M. and Zou, Y. (2010). Sonic hedgehog induces response of commissural axons to Semaphorin repulsion during midline crossing. Nat. Neurosci. 13, 29-35. Perrin, F. E. and Stoeckli, E. T. (2000). Use of lipophilic dyes in studies of axonal pathfinding in vivo. Microsc. Res. Tech. 48, 25-31. Pierani, A., Brenner-Morton, S., Chiang, C. and Jessell, T. M. (1999). A sonic hedgehog-independent, retinoid-activated pathway of neurogenesis in the ventral spinal cord. Cell 97, 903-915. Placzek, M. and Briscoe, J. (2005). The floor plate: multiple cells, multiple signals. Nat. Rev. Neurosci. 6, 230-240. Quinlan, K. A. and Kiehn, O. (2007). Segmental, synaptic actions of commissural interneurons in the mouse spinal cord. J. Neurosci. 27, 6521-6530. Roelink, H., Porter, J. A., Chiang, C., Tanabe, Y., Chang, D. T., Beachy, P. A. and Jessell, T. M. (1995). Floor plate and motor neuron induction by different concentrations of the amino-terminal cleavage product of sonic hedgehog autoproteolysis. Cell 81, 445-455. Ruiz i Altaba, A., Placzek, M., Baldassare, M., Dodd, J. and Jessell, T. M. (1995). Early stages of notochord and floor plate development in the chick embryo defined by normal and induced expression of HNF-3 beta. Dev. Biol. 170, 299-313. Salinas, P. C. (2003). The morphogen sonic hedgehog collaborates with netrin-1 to guide axons in the spinal cord. Trends Neurosci. 26, 641-643. Sander, M., Paydar, S., Ericson, J., Briscoe, J., Berber, E., German, M., Jessell, T. M. and Rubenstein, J. L. (2000). Ventral neural patterning by Nkx homeobox genes: Nkx6.1 controls somatic motor neuron and ventral interneuron fates. Genes Dev. 14, 2134-2139. Serafini, T., Kennedy, T. E., Galko, M. J., Mirzayan, C., Jessell, T. M. and Tessier-Lavigne, M. (1994). The netrins define a family of axon outgrowthpromoting proteins homologous to C. elegans UNC-6. Cell 78, 409-424. Shirasaki, R. and Pfaff, S. L. (2002). Transcriptional codes and the control of neuronal identity. Annu. Rev. Neurosci. 25, 251-281. Stepien, A. E. and Arber, S. (2008). Probing the locomotor conundrum: descending the ‘V’ interneuron ladder. Neuron 60, 1-4. Stoeckli, E. T. (2006). Longitudinal axon guidance. Curr. Opin. Neurobiol. 16, 35-39. Strahle, U., Lam, C. S., Ertzer, R. and Rastegar, S. (2004). Vertebrate floor-plate specification: variations on common themes. Trends Genet. 20, 155-162. Sun, T., Echelard, Y., Lu, R., Yuk, D. I., Kaing, S., Stiles, C. D. and Rowitch, D. H. (2001). Olig bHLH proteins interact with homeodomain proteins to regulate cell fate acquisition in progenitors of the ventral neural tube. Curr. Biol. 11, 1413-1420. Sussel, L., Kalamaras, J., Hartigan-O’Connor, D. J., Meneses, J. J., Pedersen, R. A., Rubenstein, J. L. and German, M. S. (1998). Mice lacking the homeodomain transcription factor Nkx2.2 have diabetes due to arrested differentiation of pancreatic beta cells. Development 125, 2213-2221. Tamada, A., Kumada, T., Zhu, Y., Matsumoto, T., Hatanaka, Y., Muguruma, K., Chen, Z., Tanabe, Y., Torigoe, M., Yamauchi, K. et al. (2008). Crucial roles of Robo proteins in midline crossing of cerebellofugal axons and lack of their upregulation after midline crossing. Neural Dev. 3, 29. Tresch, M. C. and Kiehn, O. (1999). Coding of locomotor phase in populations of neurons in rostral and caudal segments of the neonatal rat lumbar spinal cord. J. Neurophysiol. 82, 3563-3574. DEVELOPMENT Nkx2 transcription factors regulate floor plate development Wegmeyer, H., Egea, J., Rabe, N., Gezelius, H., Filosa, A., Enjin, A., Varoqueaux, F., Deininger, K., Schnutgen, F., Brose, N. et al. (2007). EphA4dependent axon guidance is mediated by the RacGAP alpha2-chimaerin. Neuron 55, 756-767. Wurst, W., Auerbach, A. B. and Joyner, A. L. (1994). Multiple developmental defects in Engrailed-1 mutant mice: an early mid-hindbrain deletion and patterning defects in forelimbs and sternum. Development 120, 2065-2075. Development 137 (24) Zhang, Y., Narayan, S., Geiman, E., Lanuza, G. M., Velasquez, T., Shanks, B., Akay, T., Dyck, J., Pearson, K., Gosgnach, S. et al. (2008). V3 spinal neurons establish a robust and balanced locomotor rhythm during walking. Neuron 60, 84-96. Zou, Y., Stoeckli, E., Chen, H. and Tessier-Lavigne, M. (2000). Squeezing axons out of the gray matter: a role for slit and semaphorin proteins from midline and ventral spinal cord. Cell 102, 363-375. DEVELOPMENT 4260 RESEARCH ARTICLE