Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
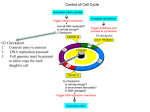
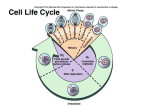
Journal of Experimental Botany, Vol. 54, No. 385, pp. 1125±1126, April 2003 DOI: 10.1093/jxb/erg138 PLANT CULTURE: SEARCHING QUESTIONS Why should we study the plant cell cycle? Dirk InzeÂ1 Department of Plant Systems Biology, Flanders Interuniversity Institute for Biotechnology (VIB), Ghent University, KL Ledeganckstraat 35, B-9000 Gent, Belgium Received 14 January 2003; Accepted 20 January 2003 ABSTRACT Description of the molecular biology of plant and animal cell cycles highlights similarities and critical differences. The cell cycle is a point of control in both growth and development and deepening understanding of underlying processes and mechanisms may have many practical applications. Key words: Animal cell cycle, cell cycle genes, cell proliferation, cell walls, development, growth, plant cell cycle. Introduction Ten years of molecular cell cycle research in plants has given a picture of how plant cells divide that has still to be re®ned. Most interesting was the discovery that many of the fundamental control mechanisms that govern cell division in animals are also conserved in higher plants. Like animals, plants use cyclin-dependent kinases (CDKs), cyclins, CDK inhibitors, retinoblastoma, E2F/DP transcription factors, and WEE kinases to control the progression through the various cell cycle phases (Stals and InzeÂ, 2001). Whereas, in some cases, genes (such as those coding for the A-type CDK, the docking factor CKS, and some cyclins) are functionally interchangeable between organisms, in other cases the conservation is more structural rather than functional. Examples of the latter are the plant CDK inhibitors, now designated Kip-related proteins (KRPs; De Veylder et al., 2001), which share very little sequence homology with their mammalian counterparts, but nevertheless most probably exert similar functions. What does all this mean? Can the plant cell cycle teach anything about the animal cell cycle? How crucial is the plant cell cycle as a point of control in plant development? Below are listed a few thoughts that I feel are worthy of consideration. Cell walls The rigid cell walls of plants prevent division by constriction, therefore plants have developed an intriguing and novel mechanism to construct new cell walls. Early in prophase, a microtubular array, called the preprophase band, positions the future cell wall and, during telophase, another microtubular array, the phragmoplast, ensures the formation of the so-called cell plate that grows from the centre of the cell toward the parental cell walls. Remarkably little is 1 known about the mechanisms that position new cell walls in plants, and the molecular processes that lead to the formation of a phragmoplast and cell plate are currently being discovered. Because the process of cell wall formation in plants and animals is so different, it is not surprising that, already, several new cell cycle regulators without functional homologues in other organisms have been detected in plants. A good example is the existence of a novel, plant-speci®c class of CDKs that control the G2-to-M transition (Porceddu et al., 2001; Sorrell et al., 2001). Cell cycle genes Since the entire genome sequence of Arabidopsis thaliana has become available, it has become clear that the complement of cell cycle genes is even more complex in plants than in mammals. For instance, the number of CDKs, cyclins, and CDK inhibitors (KRPs) exceed those found in humans (Vandepoele et al., 2002). One rather attractive explanation for this remarkable observation is that this high complexity re¯ects the plant's sessile lifestyle. Plants cannot escape adverse conditions and are, therefore, well-adapted to adjust their development in response to changing environmental conditions. Possibly, the high number of cell cycle genes allows ®netuning of development with the probable result that land plants are so successful in colonizing different habitats. More and more data seem to support this view. For example, several cyclins have been shown to be differentially affected by plant growth regulators, such as cytokinins, brassinosteroids, and sucrose (De Veylder et al., 1999; Riou-Khamlichi et al., 2000). An alternative explanation for the reduced number of cell cycle genes in mammals when compared to that in plants is the frequent occurrence of splice variants in mammalian genes, a process that has been rarely reported in plant genes. Cell proliferation The amazingly long lifespan of some plant cells is possibly of far reaching importance. Cells in the meristems of some trees continue, year after year, to produce faithful copies of themselves, without the appearance of cancerous growths. Some scientists will argue that plants cannot form cancers because of the strictly rigid constraints of the tissues in which the cells are embedded. However, many proliferative diseases of plants, such as crown galls formed by Agrobacterium tumefaciens, show that cancerous tissues can easily proliferate. It will be of the utmost interest to understand how plant cells avoid the cell cycle running out of control and this in the Fax: +32 9 264 53 49. E-mail: [email protected] Journal of Experimental Botany, Vol. 54, No. 385, ã Society for Experimental Biology 2003; all rights reserved 1126 Inze absence of known anti-proliferative mechanisms, such as p53 in animals. Obviously, understanding such basic mechanisms might have consequences for the control of cell proliferation in animals. Presumably, in connection with this consideration is that speci®c cells in the meristems of plants have the ability to produce new daughter cells over very long periods. Such `stem' cells might be very interesting to study in greater detail in the future. The cell cycle and development Plants offer a unique possibility to study the integration of cell division, growth, and development of multicellular organisms. Because plants develop mainly post-embryonically, they have an enormous plasticity and seem to tolerate many more changes in numbers and sizes of cells than most other organisms. As such, plants will still be able to reproduce with many fewer cells or with cells that are several times larger than those of control plants. We are just starting to use this plasticity to elucidate the long-standing question as to whether cell division is the driver for growth (cellular theory) or whether cell division just follows a developmental master plan (organismal theory). Most intriguing is that the issue is still unresolved and that experimental data sometimes support the cellular theory and sometimes the organismal theory. For example, overproduction of the transcription factor E2F results in extra cells, thereby creating larger structures (De Veylder et al., 2002). On the other hand, overproduction of KRPs reduces the number of cells in the leaves by a factor of ten; but, this effect is compensated by greatly enhanced cell sizes (Wang et al., 2000; De Veylder et al., 2001). Yet another serious experiment indicates that overexpression of the cyclin genes CYCB1;1, CYCD2;1, and CYCD3;1 just speeds up the cell cycle and, consequently, organ development (Doerner et al., 1996; Cockcroft et al., 2000; Dewitte et al., 2003). How to consolidate all these observations is certainly a major challenge for the near future. Neither the cellular nor the organismal theories are correct and presumably new and more complex concepts will emerge. It is my ®rm belief that understanding how cell division is co-ordinated and integrated into the development of higher plants can help scientists to address similar questions in other organisms. An advantage of working with plants is that they have a very ¯exible development with determinate and indeterminate growing organs and a large reservoir of natural variation. Furthermore, combining a well-developed technology to analyse growth processes (division and expansion) at high spatio/temporal resolution (Beemster and Baskin, 1998) with the emerging genomics tools will yield unprecedented possibilities to anchor the cell cycle into developmental networks. Practical applications A thorough understanding of the plant cell cycle will probably be of major practical importance in enhancing biomass production and yield in major crops. Indeed, augmenting the number of cells in concert with cell growth is expected to increase biomass. In addition, acceleration of the cell cycle has been shown to reduce the lifespan of plants (seed to seed). The systematic functional analysis of cell cycle genes in crops is currently underway. References Beemster GTS, Baskin TI. 1998. Analysis of cell division and elongation underlying the developmental acceleration of root growth in Arabidopsis thaliana. Plant Physiology 116, 1515±1526. Cockcroft CE, den Boer BGW, Healy JMS, Murray JAH. 2000. Cyclin D control of growth rate in plants. Nature 405, 575±579. De Veylder L, Beeckman T, Beemster GTS, Krols L, Terras F, Landrieu I, Van Der Schueren E, Maes S, Naudts M, Inze D. 2001. Functional analysis of cyclin-dependent kinase inhibitors of Arabidopsis. The Plant Cell 13, 1653± 1667. De Veylder L, De Almeida Engler J, Burssens S, Manevski A, Lescure B, Van Montagu M, Engler G, Inze D. 1999. A new D-type cyclin of Arabidopsis thaliana expressed during lateral root primordia formation. Planta 208, 453±462. De Veylder L, Beeckman T, Beemster GTS, de Almeida Engler J, Ormenese S, Maes S, Naudts M, Van Der Schueren E, Jacqmard A, Engler G, Inze D. 2002. Control of proliferation, endoreduplication and differentiation by the Arabidopsis E2Fa/DPa transcription factor. EMBO Journal 21, 1360±1368. Dewitte W, Riou-Khamlichi C, Sco®eld S, Healy JMS, Jacqmard A, Kilby NJ, Murray JAH. 2003. Altered cell cycle distribution, hyperplasia, and inhibited differentiation in Arabidopsis caused by the D-type cyclin CYCD3. The Plant Cell 15, 79±92. Doerner P, Jùrgensen J-E, You R, Steppuhn J, Lamb C. 1996. Control of root growth and development by cyclin expression. Nature 380, 520±523. Porceddu A, Stals H, Reichheld J-P, Segers G, De Veylder L, De Pinho BarroÃco R, Casteels P, Van Montagu M, Inze D, Mironov V. 2001. A plantspeci®c cyclin-dependent kinase is involved in the control of G2/M progression in plants. Journal of Biological Chemistry 276, 36354±36360. Riou-Khamlichi C, Menges M, Healy JMS, Murray JAH. 2000. Sugar control of the plant cell cycle: differential regulation of Arabidopsis D-type cyclin gene expression. Molecular and Cellular Biology 20, 4513±4521. Sorrell DA, Menges M, Healy JMS, Deveaux Y, Amano X, Su Y, Nakagami H, Shinmyo A, Doonan JH, Sekine M, Murray JAH. 2001. Cell cycle regulation of cyclin-dependent kinases in tobacco cultivar Bright Yellow-2 cells. Plant Physiology 126, 1214±1223. Stals H, Inze D. 2001. When plant cells decide to divide. Trends in Plant Science 6, 359±364. Vandepoele K, Raes J, De Veylder L, Rouze P, Rombauts S, Inze D. 2002. Genome-wide analysis of core cell cycle genes in Arabidopsis. The Plant Cell 14, 903±916. Wang H, Zhou Y, Gilmer S, Whitwill S, Fowke LC. 2000. Expression of the plant cyclin-dependent kinase inhibitor ICK1 affects cell division, plant growth and morphology. The Plant Journal 24, 613±623.