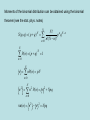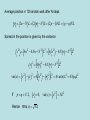Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
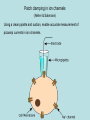
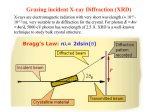
Discussion topic for week 1 • Eukaryotes (multi-cell organisms) evolved into very large sizes whereas prokaryotes (single-cell organisms) remained quite small (about 1 micrometer). What has prevented prokaryotes from growing to larger sizes? Weekly discussion topics are listed in the web page: www.physics.usyd.edu.au/~serdar/bp/bp.html Reminder: please look at the statistical physics notes in the web page and make sure that you have the necessary background. Basic properties of cells (Nelson, chap. 2) • Fundamental structural and functional units • Use solar or chemical energy for mechanical work or synthesis • Protein factories (ribosome) • Maintain concentration differences of ions, which generates a potential difference with outside (-60 mV) • Sensitive to temperature, pressure, volume changes • Respond to changes in environment via sensors and motility • Sense and respond to changes in internal conditions via feedback and control mechanisms (extreme example: apoptosis--cell death) Two kinds of cells: • Prokaryotes (single cells, bacteria, e.g. Escherichia coli) Size: 1 mm (micrometer), thick cell wall, no nucleus The first life forms. Simpler molecular structures, hence easier to study Flagella: long appendages used for moving • Eukaryotes (everything else) Size: 10 mm, no cell wall (animals), has a nucleus, Organelle: subcompartments that carry out specific tasks e.g. mitochondria produces ATP from metabolism (the energy currency) chloroplast produces ATP from sunlight Cytoplasm: the rest of the cell Structure of a typical cell Plasma membrane Molecular parts Electrolyte solution: water (70%) ions (Na+, K+, Cl-,…) Organic molecules Hydrocarbon chains (hydrophobic) Double bonds Functional groups in organic molecules Polar groups are hydrophilic. When attached to hydrocarbons, they modify their behaviour. Four classes of macromolecules: polysaccharides, triglycerides, polypeptides, nucleic acids. (sugars) (lipids) (proteins) (DNA) Simple sugars (monosaccharides): e.g. ribose (C5H10O5), Glucose is a product of photosynthesis Glucose and fructose have the same formula (C6H12O6) but different structure Disaccharides are formed when two monosaccharides are chemically bonded together. Lipids (fatty acids) are involved in long-term energy storage Saturated fatty acids Unsaturated fatty acid (C=C bonds) Phospholipids are important structural components of cell membranes Phosphatide: At normal pH (7), the oxygens in the OH groups are deprotonated, leading to a negatively charged membrane. Phospatidylcholine (PC): The most common phospholipid has a choline group attached ….PO4-CH2-CH2-N+-(CH3)3 Proteins (polypeptides) perform control and regulatory functions (e.g. enzymes, hormones, ). The building blocks of proteins are the 20 amino acids. pH 2 10 pH 2 pH 10 NH3+ - C - COOH NH3+ - C - COONH2 - C - COO- Formation of polypeptides In water: NH 3+ - C - COO - + NH 3+ - C - COO NH 3+ - C - CO - NH - C - COO - + H2 0 Protein structure 3.6 amino acids per turn, r=2.5 Å pitch (rise per turn) is 5.4 Å -helix b -sheet Nucleic acids are formed from ribose+phosphate+base pairs The base pairs are A-T and C-G in DNA In RNA Thymine is substituted by Uracil Adenosine triphosphate (ATP) has three phosphate groups. In the usual nucleotides, there is only one phosphate group which is called Adenosine monophosphate (AMP) Another important variant is Adenosine diphosphate (ADP) B-DNA (B helix) ROM (Read-Only Memory) contains1.5 Gigabyte of genetic information Base pairs per turn (3.4 nm): 10 Primary structure of a single strand of DNA Primary structure of a single strand of RNA Hydrogen bonds among the base pairs A-T and C-G Local structure of DNA Dynamic and flexible structure Bends, twists and knots Essential for packing 1 m long DNA in 1 mm long nucleus Central dogma Tools of Molecular Biology • X-ray diffraction • Nuclear magnetic resonance (NMR) spectroscopy • Electron microscopy • Atomic force microscopy • Mass spectrometry • Optical tweezers (single molecule exp’s) • Patch clamping (conductance of ion chanels) • Computational tools (molecular dynamics, bioinformatics, etc.) See, Methods in Molecular Biophysics by Serdyuk et al. for detailed discussion of these methods Mass spectrometer Charged biomolecules are accelerated and injected to the velocity selector which has transverse E and B fields. Velocity selector Only those which have velocity v= E/B will pass through. In the next chamber, there is only a B field, which bends the beam by r = mv/Bq. The mass is accurately determined from the measured radius of gyration. Optical tweezers Single molecule experiment using optical tweezers. Increasing the force on the bead triggers unfolding of RNA (Bustamante et al, 2001). Patch clamping in ion channels (Neher & Sakmann) Using a clean pipette and suction, enable accurate measurement of picoamp currents in ion channels. X-ray diffraction Basics 1. Accelerating charges emit radiation dP e2 2 2 a sin 3 d 4c Larmor’s formula, non-relativistic Where a is the acceleration of the charge and is the angle between the acceleration and radiation vectors. • Maximum radiation occurs in the direction perpendicular to a. • The only way to increase the intensity of radiation is via a. Generic x-ray tubes use bremstrahlung (breaking radiation) Isotropic, only selected wavelengths, low intensity Synchrotrons accelerate electrons around a circular path (relativistic) Directional, continuous, intense (one is operating in Melbourne now!) 2. Charged particles scatter incident radiation X-rays are electromagmetic radiation with l 10 nm E E0 exp[i(k.r - t )], k 2 / l , ck Where E is the the electric field amplitude, k is the wave vector and is the frequency. An EM wave scattered by a charged particle has the amplitude q2 E ' E0 2 sin mc Where is the angle between incident and scattered radiation. Because nuclei are much heavier than electrons, they can be ignored. Note the q dependence; light atoms (e.g. H, He) are much harder to see. Scattering from a collections of atoms is descibed using form factors f (q) (r ) exp[ iq.r ] dV Where is the charge density, q is the momentum transfer in the scattering, i.e. q = k-k'. Thus form factor is just the Fourier transform of the charge density X-ray scattering provides information on f, which is then inverted via inverse Fourier transform to find the electron density maps (r ) 1 (2 ) 3 f (q) exp[ -iq.r ] dV X-ray scattering from a single atom Atom in space 1D cut in FT 2D cut in FT X-ray scattering from two atoms Braggs law: nl = 2d.sin() Atoms in space 1D cut in FT 2D cut in FT X-ray scattering from 5 atoms in a row Atoms in space 1D cut in FT 2D cut in FT X-ray scattering from a lattice of atoms Atoms in space reciprocal space X-ray scattering from a monoclinic lattice (75 degrees) Atoms in space reciprocal space X-ray scattering from a square box Atoms in space reciprocal space X-ray scattering from a circular box Atoms in space reciprocal space Random Walks and Diffusion (Nelson, chap. 4) Friction: when an object moves faster than its fair share (i.e. Ekin>3kT/2) its kinetic energy is degraded by the surrounding molecules. Examples of kinetic energy: a) 1 kg ball with speed 1 m/s: Ekin= 0.5 J ≈ 1020 kT Average speed after equilibration: v 1.6 kT / m 10-10 m/s b) 1 ng cell with speed 1 mm/s: Ekin= 0.5 x 10-18 J ≈ 100 kT Average speed after equilibration: v 10 - 4 m/s 0.1 mm/s At that speed, the cell could move 10 times its size in 1 second! Mesoscopic objects in liquid execute a random motion called Brownian (Dr Robert Brown, 1828). Brownian motion arises from random kicks of molecules (Einstein, 1905) Random walk in 1D Toss a coin and take a step (of length L) to the right if it is heads, and to the left if it is tails. If we get n heads after N throws, the position will be x nL - ( N - n) L (2n - N ) L Repeating this experiment many times, we will get a distribution of positions in the range [-NL, NL]. Since x and n have a 1-to-1 correspondence, the same distribution applies to that of heads & tails. This is given by the binomial distribution: Given that the probability of throwing a head is p and tail q (p+q=1), that of n heads out of N trials is N! P ( n) p n q N -n n!( N - n)! Moments of the binomial distribution can be obtained using the binomial theorem (see the stat. phys. notes) N S ( p, q ) ( p + q ) N n 0 N! p n q N -n n!( N - n)! N N P ( n ) ( p + q ) 1 n 0 N n nP(n) pN n 0 n 2 N n 2 P ( n) n 2 + Npq n 0 var( n) n 2 - n 2 Npq Average position in 1D random walk after N steps x 2n - N L 2 n - N L (2 p - 1) NL ( p - q) NL Spread in the position is given by the variance x 2 4n 2 - 4 Nn + N 2 L2 4 n 2 - 4 N n + N 2 L2 x 2 4n var( x) x 2 - x 2 4 n2 - n If p q 1 / 2, 2 - 4 N n + N 2 L2 2 L 2 4 var( n) L2 4 NpqL2 x 0, var( x) x 2 NL2 Hence rms( x) N L Connection with the molecular world: Molecular collisions occur randomly. Nevertheless we can still define a mean collision time (Dt) and a mean free path (L), which allows us to introduce time via t NDt or N t / Dt 2 L x 2 (t ) NL2 t Dt We define D L2 2Dt as the diffusion coefficient The mean-square displacement becomes x 2 2 Dt Generalisation to 2D and 3D is straightforward 2D : r 2 x 2 + y 2 x 2 + y 2 4 Dt 3D : r 2 x 2 + y 2 + z 2 6 Dt Examples of 1D random walk Squared displacement Mean-square displacement for a single random walk for 30 random walks In both graphs, the lines describe the diffusive motion, It is satisfied only for the ensemble average. x 2 2 Dt Example of 2D random walk Perrin’s experimental data for Brownian motion of a colloid particle (size: 0.075 mm) t=300 t=300 N=300 N=7500 Dt=1 Dt=1/25 L=1 L=1/5 D=0.5 D=0.5 Computer simulation of random motion in 2D Mean collision times of molecules in liquids are of the order of picosec. Thus in macroscopic observations, N is a very large number. Large N limit of the binomial distribution is Gaussian (see stat. phys.) P ( n ) P ( n )e where -( n - n ) 2 2 Npq 1 -( n - n ) 2 e 2 2 2 1 1 n Np , P(n ) , Npq 2 2Npq For the position variable x (2n - N ) L, we have x Npq 2 L x - x ( n - n ) 2 L, P( x) 1 2 x e -( x - x ) 2 2 x2 x (2n - N ) L (2 p - 1) NL ( p - q) NL Other examples of random walk: 1. Polymer conformations They have a random coil structure Single step size 3L rms distance for N links: 3L rrms 3N L Mass is proportional to N and diffusion coeff. is proportional to 1/r N-1/2 (for close packing, N-1/3) 2. Stock market 3. Gambling -0.57 (fit to exp) -0.5 Gambling as an example of biased random walk Biasing is worst in poker machines Roulette provides one of the least biased form of gambling Chances of winning with red or odd 100 x 18/37 ~ 49% Friction Macroscopic observation: motion of an object in a viscous medium is damped by a force proportional to its speed: dv m F - v dt dv F 0 vter dt (terminal velocity) For a spherical object, the friction coefficient is given by Stokes formula 6R where R is the radius of the object and is the viscosity of the medium. Typical values for (kg/ms): air: 10-5, water: 10-3, oil: 0.1 For a cell in air, vter ≈ 5 cm/s Microscopic interpretation: motion of the object is modified by random molecular collisions. We model this motion via 1D random walk subject to an external force f. In between collisions, the object moves by 1 f 2 Dxi vi Dt + Dt 2m Dx Dv Dt + in one step 1 f 2 average over many steps Dt 2m Since v is randomly oriented Dv=0. Introduce the drift velocity as Dx Dt vd f Dt 2m L2 Combine with D , 2Dt 2m Dt D m L2 Dt 2 m v 2 kT Einstein relation