Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
Management of acute coronary syndrome wikipedia , lookup
Cardiac contractility modulation wikipedia , lookup
Rheumatic fever wikipedia , lookup
Hypertrophic cardiomyopathy wikipedia , lookup
Heart failure wikipedia , lookup
Coronary artery disease wikipedia , lookup
Electrocardiography wikipedia , lookup
Quantium Medical Cardiac Output wikipedia , lookup
Artificial heart valve wikipedia , lookup
Myocardial infarction wikipedia , lookup
Lutembacher's syndrome wikipedia , lookup
Mitral insufficiency wikipedia , lookup
Atrial septal defect wikipedia , lookup
Heart arrhythmia wikipedia , lookup
Arrhythmogenic right ventricular dysplasia wikipedia , lookup
Dextro-Transposition of the great arteries wikipedia , lookup
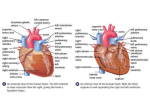
A M . ZOOLOGIST, 8:221-229 (1968). Functional Morphology of the Heart in Mammals Ursula Rowlatt Chicago Zoological Park, Brookfield, Illinois, 60513, and Lincoln Park Zoological Gardens, Chicago, Illinois SYNOPSIS. The heart in all mammals is composed of four separate chambers and in broad outline is very similar in monotremes, marsupials, and placenta! mammals. The functional anatomy of the chambers and valves of the dog's heart has been investigated by Rushmer (1961) and others. They concluded that the atria serve as reservoirs and the ventricles as reciprocating pumps, the right ventricle being a volume pump and the left ventricle a pressure pump. Similar investigations in man, horses, cattle, sheep, and goats suggest that other mammalian hearts act in the same way. However, there is a wide range of variation in heart weight relative to body weight, in heart shape, and in minor points of internal anatomy. The general contention in this paper is that these differences should be considered functionally significant unless proven otherwise. Certain morphological structures such as the valves of the posterior vena cava, Lower's tubercle, the atrial appendages, and the more extreme forms of double ventricular apex may not have functional significance. Verbal descriptive anatomy has probably reached the end of its usefulness and should be replaced with measurements. A correlation of quantitative anatomy and hemodynamics would seem to be the most promising line of future research. The heart of any mammal is composed of four separate chambers arranged in parallel and is very similar in construction in monotremes, marsupials, and placental mammals. A functional explanation can be given in general terms for the gross morphology of each chamber. Such an explanation probably holds good for mammals as a whole and will be given first. However, there are many differences in structural detail between genera, species, and individuals and, if these are to be explained in functional terms, certain evolutionary and morphogenetic processes must be taken into consideration. These will be discussed in the second part of the paper. Most of the following observations are based on the work of Rushmer and coworkers using several different techniques in dogs (Rushmer, 1961). The dynamics of the heart in horses, cattle, sheep, and goats are similar (Sporri, 1965). The atria and contiguous systemic and pulmonary veins act as expanding reservoirs for blood returning from the body and lungs. Semilunar valves preventing backflow are present in the major systemic veins proximal to their entry into the thoracic cavity. There is no valve at the mouth of the anterior vena cava and only in some species is there a valve at the mouth of the posterior vena cava (Eustachian valve) or coronary sinus (Thebesian valve). The muscle fibers of the atria are arranged in two groups: (1) those that are common to both atria, and (2) those that belong to each atrium. The common fibers are superficial (Robb, 1965). Constriction of the muscle bands around the orifices of the systemic veins may reduce if not prevent backflow during atrial systole (Keith, 1904). There are no valves in the pulmonary veins, but a muscular valvular mechanism was postulated by Keith and has been demonstrated in the living dog by Little (1960). Much of the atrial transport of blood is brought about by elastic recoil of the wall. Blood will move either forwards through the atrioventricular valves or backwards along the veins depending on which direction provides the least resistance to flow. Only a small part of the expulsive force of the atria is by active muscular contraction and this occurs at the very end of ventricular diastole. The ventricular myocardium is composed of two thin layers of spiral muscle bundles running more or less at right angles to each other and an intervening constrictor layer. The superficial spiral 221 222 URSULA ROWLATT muscles arise from the atrioventricular rings, run towards the apex, reverse their direction forming a vortex, and return to the base of the heart as the deep spiral layer. This layer forms the trabeculae carneae and papillary muscles. The constrictor bands lying between the spiral layers encircle the basal two-thirds of the ventricles and constitute a much greater proportion of the total thickness of the wall in the left than in the right ventricle. Emptying of the right ventricle is brought about by three mechanisms occurring at about the same time: (1) contraction of the spiral muscles shortens the long axis of the ventricle, pulling the right atrioventricular ring towards the apex, which remains stationary; (2) contraction of the constrictor muscle layer moves the free wall towards the septum in a bellows-like action (the surface area of the ventricle is large compared to its volume so that a small excursion of the free wall will displace a large volume of blood); (3) contraction of the constrictor fibers of the left ventricle will augment the bellows action by pulling on the free wall of the right ventricle at the interventricular groove. This last mechanism has been shown to be capable of adequate ventricular emptying when the free wall has been destroyed by cautery in dogs or by coronary occlusion in man. Presumably it is of even greater significance in animals such as bats, in which the septum bulges well into the right ventricle at all times. The conus of the right ventricle has been shown to behave as a separate chamber. Conal diastole is still occurring when the sinus has begun to contract, and conal systole overlaps sinus relaxation (March, Ross, and Lower, 1962). This dissociation is in keeping with the ontogenetic and phylogenetic history of the conus, which represents the bulbus cordis of lower forms (Keith, 1924). Left ventricular emptying is brought about mainly by a squeezing action caused, by contraction of the constrictor layer. This action reduces the diameter of the chamber and is responsible for the power of ejection. The power of thrust is augmented by the elastic recoil of the aorta after closure of the aortic valve. The spiral muscles shorten the long axis of the chamber, pulling the left atrioventricular ring towards the apex. Rushmer (1961) concluded that the right ventricle is a volume pump ideally suited to the disposal of large and varying amounts of blood against a low peripheral resistance. The left ventricle acts as a pressure pump, capable of forceful contraction against a much higher peripheral resistance. This difference in function is related to the different disposition of musclefibersin each ventricle. The functional anatomy of the atrioventricular valves was demonstrated in real and simulated heart valves by Henderson and Johnson in 1912. They showed that two theoretical actions are possible. There is (1) retrograde flow of blood towards the atria at the beginning of ventricular contraction. This action lifts the valve leaflets from below, bringing them together, but inevitably results in leakage of blood through the center of the closing orifice. However this is prevented by (2) the occurrence at the same time of a suction effect caused by the momentum of the column of blood as it is forced through the valve at the end of atrial contraction. The degree of excursion of the valve leaflets has been studied in intact dogs by placing small metal clips on the atrial side of the leaflets and by recording their movement by cinecardiography. It was seen that the metal clips moved only very short distances during the cardiac cycle presumably due to continuous restraint by the chordae tendineae (Rushmer, 1961). Because both sets of atrioventricular valves look superficially the same in general construction, it has been inferred that their action is similar. However, the morphology of the left atrioventricular apparatus is much more uniform among mammals than is that of the right atrioventricular valve. The number of valve leaflets, the arrangement of trabeculae carneae at the valve ring, and the attachment of the papillary MORTHOLOGY OF THE HEART IN MAMMALS muscles varies greatly from species to species and between individuals of the same species. That each valve is constructed specifically for its own position in the heart is suggested by the course of events in a human congenital anomaly called ventricular inversion. In this abnormality, the heart is normal in all respects except that the ventricle on the right side of the heart is a morphological left ventricle and that on the left a morphological right ventricle, as judged by the appearance of the ventricular septum (Lev, 1954). Since the configuration of the valvular apparatus depends on the arrangement of the papillary muscles, which are in themselves part of the morphology of the ventricle from which they arise, the right atrioventricular valve is a morphological mitral valve and the left atrioventricular valve is a tricuspid valve. Such a heart may function properly for many years, but the limiting factor is the developmemt of tricuspid (left atrioventricular) incompetence and final heart failure from the accumulated effects of regurgitation of blood through this orifice. From this one can infer that the exact morphology of the mitral valve is required for proper control of the left atrioventricular orifice and that another apparently similar valve is less efficient. Ventricular inversion is in effect a translocation of morphological units within the heart, and its consideration at this point illustrates the usefulness of studying the abnormal alongside the normal for the better understanding of both. The semilunar valves at the pulmonary and aortic orifices are mechanically much simpler. Each consists of three symmetrical cusps. It has been shown that two cusps would close equally well but would require more elastic stretch to open adequately (Rushmer, 1961). Closure is brought about partly by suction following the initial rush of blood through the orifice during ventricular systole and partly by lateral pressure from eddy currents in the sinuses of Valsalva. Usually two coronary arteries arise from the base of the aorta, but the larger branches 223 of either may have an independent origin as indicated by supernumerary ostia. The coronary arteries and their main branches lie either on the surface of the heart beneath the epicardium or deep to the superficial layer of the myocardium where they lie parallel to the muscle fibers. Further subdivisions arise at right angles to the main branches and run between the muscle fibers to supply all layers of the myocardium. In some mammals, the ventricular septum is supplied by a special septal artery arising from the left coronary artery close to its origin and running on the low pressure side of the septum beneath the endocardium of the right ventricle (see Robb, 1965, for references). The resistance to blood flow in the coronary arteries varies enormously during the cardiac cycle because of the squeezing effect of the surrounding myocardial fibers. Flow is greatest in diastole. Large animals and athletes have a slow heart beat and a long diastole which allows the heart muscle to recover between contractions. Venous drainage is by way of the coronary veins which enter the right atrium by various routes. The Thebesian veins probably play an arterial role and supply the deeper parts of the myocardium when the arteries are closed during ventricular systole (Burton, 1965). However, within this general morphological framework mammalian hearts differ in several important respects. There is a wide range of heart weight relative to body weight. The shape of the heart differs markedly from mammal to mammal. Various points of external and internal anatomy make it possible to identify hearts from distant and closely related genera with a fair degree of confidence. However, it must be emphasized that the intraspecific variation is great and that any morphological detail should be examined in many individuals before it can be considered to be a constant entity. Extra caution should be taken in animals that have been bred selectively such as domestic animals and pets. The ratio of heart weight to body weight (heart ratio) is independent of body weight 224 URSULA ROWLATT within a wide range of body size. Stahl (1965) plotted heart weight against body weight on a log-log scale for figures given by Brody for animals ranging in size from mice (25 g) to steers (500 to 1000 kg). He also obtained an allometric line, indistinguishable statistically from that obtained from Brody's data, using measurements for 321 primates of body weight from 10 g to 100 kg. For many years it has been assumed that animals with a higher ratio of heart weight/body weight have more efficient hearts and possess, therefore, a physiological or environmental advantage over animals of similar body weight but lower heart ratio. Clark (1927) noticed that animals capable of continuous and severe muscular exertion had heart ratios greater than 0.6 and those incapable of such exertion had heart ratios of less than 0.6. Furthermore, the heart ratio of a wild animal such as a rabbit was greater than that of a tame one, and dogs bred for speed had a higher heart ratio than mongrels. Increased heart weight is due to increased left ventricular weight which is itself related to stroke volume. The physiological responses to exercise are complex and variable under different conditions, but, in general, increased cardiac output in response to increased venous return is brought about more significantly by a rise in heart rate than by increased stroke volume. A large heart in a continuously active animal probably merely reflects an increase in general muscle bulk and a large capillary network. Animals that run fast over small distances are not exceptionally muscular and do not possess proportionately large hearts. Clark's observations are correct but the interpretation is misleading. The variation in heart shape in mammals (Fig. 1), although striking, has received no adequate explanation. Casual observation suggests that the shape of the heart follows the shape of the chest. The whale's heart is broad and flat and lies in a broad thoracic cage with an obliquely slanting diaphragm (Slijper, 1962). The ungulate heart is long and narrow as is the thorax. The peculiar shape of the mole's heart is typical of the marked modification of the common mammalian plan in the animal as a whole. In man, the cardiothoracic ratio, as seen radiologically, is sufficiently constant over a large range of body size to be used as an index of cardiac normality (Oberman, Myers, Karunas, and Epstein, 1967). The same general relationship is seen in dogs, but ratios relating cardiac silhouette to chest dimensions have not been found reliable for diagnosis (D. K. Detweiler, personal communication). The most extreme view relating chest to heart size is that of Davis (1964). He studied the hearts of 14 bears, 3 procyonid carnivores, 4 canids, and 7 telids. He found that the right ventricle was broader in bears, procyonids, and dogs than in cats. He observed (p. 567) that: "the heart is part of a regional growth area that includes the entire thorax. Accelerated transverse growth in this area in bears and procyonids affects that part of the heart (the venous portion of the right ventricle) that is undergoing rapid differentiation and growth at the moment when the acceleration manifests itself." "The differences between the right ventricle of bears and procyonids and the right ventricle of other carnivores are an accident of ontogenetic timing. They do not represent functional adaptations." Although ingenious, this explanation is purely speculative. It is based on examination of very few specimens, and thoracic measurements fundamental to the argument are lacking. Cardiac morphology has been evolved in the vertebrates as a whole in response to physiological demands and might reasonably be supposed to be doing so within the mammalian class. In my opinion, a functional explanation for heart shape should be expected until proved to be false. The next important step is to collect numerous data on dimensions of the heart and chest of living and dead mammals to establish if chest and heart size are interdependent or not. Internal as well as external 225 MORPHOLOGY OF THE HEART IN MAMMALS measurements of a dead heart will only be meaningful if fixation is standardized (Glagov, Eckner, and Lev, 1963). The old belief that a slender, pointed heart is character- DOG istic of fast-moving animals could then be tested (Lechner, 1942). By applying Laplace's law, which relates intraluminal pressure, intramural tension, and radius of a GIBBON BAT HEDGEHOG FIG. 1. The hearts of the dog (Cants familiaris), gibbon (Hylobates lar), deer (Dama dama), bat (Eidolon helvum), rat (Rattus ratlu.s), rabbit (Oryctolagus cunicuhis), hedgehog (Erinacem europaea), RAT MOLE DEER RABBIT SLOTH mole (Talpa europaea), and sloth (Choloepas didactylus), to show the variation in the shape o£ mammalian hearts. 226 URSULA ROWLATT chamber, it is likely that a long narrow left ventricle with a low diastolic volume would require less myocardial tension to develop an adequate intraventricular pressure than would a more rounded chamber. Such a heart would be more suitable for sustaining chronic pressure loads as in long-distance running. Many hypotheses such as this can be tested by correlating physiological and anatomical data. Examination of the mammalian heart outside the body shows that there is considerable structural variation in the component parts. The basic supposition is that these features are useful to the animal and are important to the efficient working of the heart. However, the development of the mammalian heart has a long and complicated evolutionary history, and several non-functional influences must be taken into account in interpreting structural variation. Various anatomical features have been identified as being primitive. Benninghoff (1933) recognized that concentration of parts of the heart is a fundamental process in vertebrate cardiac evolution and that this process continues in mammals. The sinus venosus has been absorbed into the right atrium in most mammals, but in monotremes and the armadillo, Dasypus, it remains as a small, shallow chamber at the confluence of the three venae cavae. In these animals it is separated from the right atrium by a competent semilunar valve. Two pulmonary veins enter the left atrium by way of a separate vestibule in monotremes and marsupials but open directly into the wall of the atrium in placental mammals. The vestibule is still present in the developing embryo of placental mammals. Often the main tributaries of the pulmonary veins drain independently into the roof of the left atrium, but much variation occurs. The bulbus cordis is always incorporated into the right ventricle, but, as the conus, may be more prominent in some species than others. Even when absorption of the developmental components of the heart is complete, evidence of their former relations is given by the coronary arteries, the main branches of which overlie the primitive subdivisions of the heart. A coarse trabecular pattern in the free wall of the right ventricle is the primitive condition in phylogeny and ontogeny. Intraspecific variation must be taken into consideration as always, but it seems that the wall of the right ventricle of some mammals is strikingly smooth. Other patterns are found fairly consistently in other mammals (Fig. 2) Any functional explanation of the trabeculation should be directed at the smooth rather than the coarse pattern. The right atrioventricular valve in monotremes is mainly muscular as in birds and crocodilians. In marsupial and placental mammals, an occasional chorda tendinea may be muscular, indicating its origin from the muscular wall of the ventricle, but in general the right atrioventricular valve is formed of connective tissue only. Modern mammals are subdivided into three infraclasses on the basis of their prenatal development and by inference on the course and duration of the embryonic circulation. It follows that intra-uterine circulatory requirements determine the morphology of the right atrium and atrial septum. This should be the easiest area of the heart to understand in functional terms but in practice it is the most difficult. The functions of the residual valves of the embryonic sinus venosus and the muscle bands in the wall of the atrium have mystified anatomists since Eustachius described his valve in 1563 and Lower his tubercle in 1669. The copious literature on the comparative anatomy of these structures has been reviewed by Franklin (1948). No clear interpretation of their function has been given to this day, nor has it been established except in rare cases, if these structures are of importance in postnatal life. The sinus valve leaflets in bats and in the mole have migrated into the mouth of the posterior vena cava where they form a semilunar valve that has been shown by 227 MORPHOLOGY OF THE HEART IN MAMMALS YELLOW FRUIT BAT AMERICAN BISON FIG. 2. Different degrees of trabeculation in the wall of the right ventricles of the yellow fruit bat cine-angiography to be competent during adult life. It is important to remember that the heart is beating throughout its transformation from a simple, coiled tube to a complicated, double-circulatory organ. The view, originally suggested by Spitzer in 1923, that the passage of blood through the developing heart is an important factor in moulding the embryonic tissues into the adult shape has received ample recent confirmation (see Jaffee, 1967, for references). Possibly, both the Eustachian valve (Franklin, 1948) and Lower's tubercle may be produced by such hemodynamic forces. The varying prominence of Lower's tubercle in the animals shown in Figure 3 may be related to the angle at which the right anterior and posterior venae cavae enter the heart. A myoendocardial ridge tends to form between two converging streams of blood in the developing heart and this process may continue in postnatal life. The existence of atrial appendages may be an example of the influence of early LAR GIBBON (Eidolon helvum), the American bison bison), and the lar gibbon (Hylobates lar). MAN (Bison WOLF DEER SEA LION FIG. 3. Position and prominence of Lower's Tubercle (shaded) in man (Homo sapiens), the wolf (Canis lupus), the deer (Dama dama), and the sea lion (Zalophus californianus). 228 URSULA ROWLATT certain physical proportions within the heart. Quantitation is needed to verify this point. Lastly, consideration of the events of cardiac phylogeny and ontogeny may help to separate morphological features that are probably ancestral from those that may have functional significance. The greatest need is for correlation of in vivo physiological observations, such as those provided by angiocardiography and telemetry, with the presence of structures that seem to suggest, on theoretical grounds, a functional rather than a non-functional origin. PLACENTAL MAMMAL MARSUPIAL FIG. 4. Position of the right atrial appendage relative to the root of the aorta. events in the development of the heart on adult morphology. No function has been ascribed convincingly to these structures, and they may be removed without ill effect during cardiac surgery. Probably they are formed by pressure on the developing atrium by the truncus arteriosus as the heart tube bends to accommodate itself within the pericardium. The presence of a double right atrial appendage in marsupials may be a refinement of this process. In these animals, the aorta lies farther to the right of the pulmonary trunk than in placental mammals, allowing a further compression of atrial tissue between the roots of the great vessels (Fig. 4). The markedly bifid apex of the manatee heart may be caused by asynchronous development of the two aneurysmal out-pouches of the ventricle which are destined to be the definitive ventricles (Robb, 1965). A similar explanation may be given for less marked degrees of duplication of the apex of the heart. In summary, mammalian hearts are more similar than they are dissimilar. However, variations in relative size, shape, and details of anatomy exist between species. Differences in heart shape are probably related to the extent of the capillary bed and in particular to that of the total skeletal muscle mass. Differences in shape may be related directly or indirectly to the shape of the chest or may better be correlated with exercise potential by virtue of REFERENCES BenninghofF, A. 1933. Das Herz, p. 467-556. In L. Bolk, E. Goppert, E. Kallius, and W. Lubosch, [ed.], Handbuch der vergleichenden Anatomie der Wirbeltiere, vol. 6. Urban und Schwarzenberg, Berlin and Wien. Burton, A. C. 1965. Physiology and biophysics of the circulation, Year Book Medical Publishers, Inc., Chicago, 217p. Clark, A. J. 1927. Comparative physiology of the heart. Cambridge Univ. Press, Cambridge. I57p. Davis, D. D. 1964. Anatomy of the heart in bears (Ursidae), and factors influencing the form of the mammalian heart. Morphol. Jahrb. 106:553-568. Franklin, K. J. 1948. Cardiovascular studies. Blackwell Scientific Publications, Oxford. xvi+306p. Glagov, S., F. A. O. Eckner, and M. Lev. 1963. Controlled pressure fixation apparatus for hearts. Arch. Pathol. 76:640-646. Henderson, Y., and F. E. Johnson. 1912. Two modes of closure of the heart valves. Heart 4:69-82. Jaffiee, O. C. 1967. The development of the arterial outflow tract in the chick embryo heart. Anax. Rec. 158:35-42. Keith, A. 1904. Abstract of the Hunterian lectures on the evolution and action of certain muscular structures o£ the heart. Lancet 1:555-559, 629-632, 703-707. Keith, A. 1924. Fate of the bulbus cordis in the human heart. Lancet 11:1267-1273. Lechner, W. 1942. Herzspitze und Herzwirbel. Anat. Anz. 92:249-283. Lev, M. 1954. Pathologic diagnosis of positional variations in cardiac chambers in congenital heart disease. Lab. Invest. 3:71-82. Little, R. C. 1960. Volume pressure relationships of the pulmonary-left heart vascular segment. Circulation Res. 8:594-599. March, H. W., J. K. Ross, and R. R. Lower. 1962. Observations on the behavior of the right ventricular outflow tract, with reference to its developmental origins. Am. J. Med. 32:835-845. Oberman, A., A. R. Myers, T. M. Karunas, and F. MORPHOLOGY OF THE HEART IN MAMMALS H. Epstein. 1967. Heart size of adults in a natural population Tecumseh, Michigan. Ckculation 35:724-733. Robb, J. S. 1965. Comparative basic cardiology. Grune and Stratton, New York. xxvii-)-602p. Rushmer, R. F. 1961. Cardiovascular dynamics. W. B. Saunders, Philadelphia and London. 503p. Slijper, E. J. 1962. Whales. Hutchison, London. 475p. 229 Spitzer, A. 1923. Uber den Bauplan des normalen und missbildeten Herzens (Versuch einer phylogenetischen Theorie). Virchows Arch. Pathol. Anat. 243:81-272. Sporri, H. 1965. Studies of cardiac dynamics in animals (horses, cattle, sheep, goats). Ann. New York Acad. Sd. 127:379-392. Stahl, W. R. 1965. Organ weights in primates and other mammals. Science 150:1039-1042.