Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
History of herbalism wikipedia , lookup
Gartons Agricultural Plant Breeders wikipedia , lookup
Ecology of Banksia wikipedia , lookup
Plant secondary metabolism wikipedia , lookup
Plant nutrition wikipedia , lookup
Plant defense against herbivory wikipedia , lookup
Plant use of endophytic fungi in defense wikipedia , lookup
History of botany wikipedia , lookup
Evolutionary history of plants wikipedia , lookup
Plant breeding wikipedia , lookup
Plant physiology wikipedia , lookup
Ornamental bulbous plant wikipedia , lookup
Plant morphology wikipedia , lookup
Plant evolutionary developmental biology wikipedia , lookup
Plant ecology wikipedia , lookup
Fertilisation wikipedia , lookup
Perovskia atriplicifolia wikipedia , lookup
Pollination wikipedia , lookup
Flowering plant wikipedia , lookup
Chapter 10 – Reproduction in Plants
CHAPTER 10 – REPRODUCTION IN PLANTS
Many plants are able to reproduce both asexually and sexually. Sexually
reproducing plants have alternating haploid and diploid stages in their life cycles,
and this alternation of generations is clearly visible in the life cycle of a liverwort or
a fern. In flowering plants, the haploid stage of the life cycle is much reduced. The
flowers of flowering plants are organs specialised for sexual reproduction on land,
where mechanisms are needed which allow the transfer of male to female gametes
without any danger of them drying out.
ASEXUAL REPRODUCTION IN PLANTS
10.1 Vegetative reproduction
Asexual reproduction is much more common in plants than in animals. Many
species of plants are able to reproduce both sexually and asexually. Growth in a
plant which does not involve the development of structures involved in sexual
reproduction is sometimes known as vegetative growth. So the growth of new
individuals by asexual reproduction is known as vegetative reproduction.
Fig. 10.1 shows asexual reproduction in a strawberry plant. Here, stems called
runners grow horizontally from the parent plant, and put out roots at a node (a place
on the stem from which a leaf stalk grows, and where there is a bud). The roots
grow downwards into the soil, while the bud grows upwards and forms a young
plant. Eventually, the connection between the original parent plant and the daughter
plant withers. By this time, the daughter plant will probably have begun to grow
runners of its own.
Virtually every part of one plant or another can be involved in asexual
reproduction. You may like to try to think of some examples of plants which grow
new individuals from their roots or from their leaves.
The cells in the new individuals produced by asexual reproduction are formed by
mitosis and so, of course, are genetically identical to each other and to their parent.
They form a clone. Horticulturists often wish to produce large quantities of
genetically identical plants, as these will all grow in a similar way and produce a
similar crop. This makes them easier to tend, harvest and market. Such methods as
grafting and taking cuttings have been used for hundreds of years, and still are used
very widely. However, in recent years the technique of tissue culture has been
developed which allows very large numbers of genetically identical new plants to be
produced from a very small number of original cells taken from a parent. It involves
taking a small piece of tissue from the parent plant, and placing it on sterile agar
containing nutrients and plant growth substances which stimulate the cells to divide
by mitosis. They are kept in controlled conditions of light and temperature, and
quickly grow into complete little plants which can later be removed from the agar
and planted into soil.
Asexual reproduction can be advantageous to plants growing in natural conditions,
137
Chapter 10 – Reproduction in Plants
as it can enable rapid coverage of the ground in a suitable environment. However,
the genetic uniformity of the offspring can be a problem, for example if a parasite or
other pest is able to infect these plants. If one plant has no defence against the pest,
then this is true of all of them and the whole population may be killed. This is also a
problem for plant breeders, as genetically uniform crop plants can become
vulnerable to pests and diseases which may sweep through an entire crop. Genetic
variation can help to lessen the probability that such widespread infection will occur,
as at least some of the plants may have a gene which confers resistance to the pest.
In the wild, almost all species of plants are able to reproduce sexually, so ensuring
genetic variation in their offspring. In horticulture and agriculture, plant breeders
maintain stocks of many
different varieties of crop
plants which can be used
in breeding programmes
to introduce new genes
into
commercially
available varieties.
Fig10. 1 Asexual reproduction in a strawberry plant
REPRODUCTION IN LIVERWORTS AND FERNS
Almost all plants, like animals, reproduce sexually - that is, they produce
haploid gametes which fuse to form a diploid zygote which then grows to form an
adult plant. Most of this chapter concentrates on reproduction in flowering plants,
but firstly we will look at sexual reproduction in two less familiar types of plant liverworts and ferns.
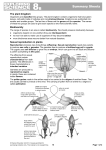
10.2 The life cycle of a liverwort
Liverworts belong to the phylum Bryophyta. They are not very big plants, and you
may never have noticed a liverwort growing. However, once you know what to look
for, they are easy to find. Liverworts grow in damp places, often where it is shady.
You may find them on the banks of streams, on bricks or stones at the base of damp
walls, or on the soil on top of flowerpots.
Fig. 10.2 shows the structure of a common liverwort in Britain called Pellia
epiphylla. The plant which you see growing is haploid - that is, each cell contains a
138
Chapter 10 – Reproduction in Plants
single set of chromosomes rather than the two complete sets which are present in
the diploid cells of almost all adult animals and flowering plants. The structure of this
haploid plant is very simple. Liverworts have no true roots or stems, just a simple
flat body called a thallus. The thallus lies close to the ground, to which it is attached
by single-celled structures called rhizoids. The rhizoids do take up water and
mineral salts from the soil, but the whole thallus can also absorb these substances
when raindrops splash onto it. Most of the cells in the thallus contain chloroplasts
and photosynthesise. Unlike the leaf of a flowering plant the thallus has no waxy
cuticle covering its surface, and so it easily loses water. This is one reason why
liverworts usually grow in damp places. There are no xylem vessels or phloem tubes,
so water cannot easily be transported to different parts of the plant, and the only
support is given by cellulose cell walls and turgor. This, combined with the problems
of water loss, prevents liverworts from growing very big.
The thallus reproduces by producing haploid gametes, and so it is called a
gametophyte ('gamete-producing plant'). On the upper surface of the thallus, male
gametes develop in small depressions called antheridia. As the cells of the thallus
are already haploid, these gametes are produced by mitosis. The male gametes are
long, thin cells with two very long flagella. They are called antherozoids, or sperm.
At the edges of the thallus, the female gametes, called oospheres or eggs, are
formed by mitosis inside archegonia. Whereas large numbers of sperm are made in
each antheridium, only one egg is made in each archegonium.
Fertilisation happens when the sperm swim to the egg while it is still in the
archegonium. They can only do this when there is a film of water on the surface of
the thallus, so that they have something to swim through and so that they do not
dry out. Fertilisation therefore happens in wet weather. Splashing of raindrops may
also be important in transferring the sperm to the eggs, especially from one thallus
to another. The sperm are attracted to the egg by chemicals, especially sugars,
which it secretes.
Fertilisation takes place inside the archegonium, producing a diploid zygote.
The zygote divides by mitosis, eventually producing a diploid structure called a
sporogonium. This stays attached to the thallus which provides it with food. At the
tip of the sporogonium a spherical structure called a spore capsule develops. Inside
this some of the diploid cells, called spore mother cells, divide by meiosis to
produce haploid spores. (A spore is one or more cells, often surrounded by a
protective waterproof covering, which can be dispersed from where it was made and
grow into a new organism.) Because the sporogonium produces spores, it is called a
sporophyte.
After a while, the spore capsule bursts, flinging out the spores. If a spore
lands in a suitable moist place, it germinates. Mitosis takes place, forming a new
haploid thallus. So you can see that there are two alternating stages or 'generations'
in the life cycle of a liverwort - a haploid gametophyte which produces gametes by
mitosis and a diploid sporophyte which produces spores by meiosis. This kind of life
cycle is said to show
Alternation of generations. The life cycle of pellia is shown in Fig. 10.3.
139
Chapter 10 – Reproduction in Plants
Fig-10.2-Reproduction in a liverwort
140
Chapter 10 – Reproduction in Plants
Fig 10.3 Life cycle of a liverwort
10.3 The life cycle of a fern
Ferns belong to the phylum Filicinophyta. Fig. 10.4 shows the structure of the
male fern, Dryopteris filix-mas. This fern is very common, and you can find it growing
in woodland and other shady places all over Britain.
Ferns, like liverworts, have a clearly marked alternation of generations in
their life cycle. The large fern plant which you see growing is the diploid sporophyte.
It is much larger, and structurally much more complex, than either the thallus or the
sporogonium of a liverwort. Ferns have true roots which can penetrate quite deeply
into the soil. The roots absorb water and minerals as well as anchor the plant. The
male fern has large leaves, called fronds, with a large surface area for the
absorption of sunlight and carbon dioxide for photosynthesis. The fronds are divided
into 'leaflets' called pinnae. The surface of the fronds is covered with a waterproof
cuticle, and there are stomata on the upper and lower epidermis which can be
closed by guard cells to conserve water. Ferns have xylem vessels and phloem
tubes; the xylem vessels transport water to all parts of the plant and also help to
support it. Thus, ferns can grow in drier places than liverworts, and they can grow
larger.
The spores are produced on the undersides of the fronds, in structures called
sporangia. (This name is very similar to, but not the same as, the 'sporogonium' in
which spores are formed in a liverwort - make sure that you do not confuse them!)
The sporangia are found in clusters called sori, and each sorus is covered by a
protective 'hood' called an indusium. Inside the sporangia, diploid spore mother
cells divide by meiosis to produce haploid spores.
The spores are released in dry conditions as a strip of cells called the
annulus, in the wall of the sporangium, curls back and rupture the sporangium. Fig.
10.4 explains how this happens. The spores are carried away on air currents. They
germinate in moist conditions, dividing by mitosis to form a haploid structure which
is quite similar to the thallus of a liverwort; it is called a prothallus. It is about one
centimetre across and very thin, so it is unlikely that you will ever have noticed one.
The prothallus has no cuticle and no supportive or specialised conducting tissue, so it
can only grow in moist conditions.
The prothallus is the gametophyte generation in the fern's life cycle. Male
gametes and female gametes are produced by mitosis inside antheridia and
141
Chapter 10 – Reproduction in Plants
archegonia respectively, both on the underside of the pro thallus. Fertilisation takes
place in a very similar way to liverworts to form a diploid zygote. As in the liverwort,
this remains inside the archegonium as it divides repeatedly by mitosis. It develops
into a new sporophyte. At first, the embryo sporophyte obtains its nutrients from the
prothallus, but once it has formed its first leaves it can photosynthesise for itself.
The gametophyte then dies, and the sporophyte continues to grow to form a new
fern plant.
You can see that the life cycle of Dryopteris (Fig. 10.5) is in many ways
similar to that of Pellia. Both of them show alternation of generations, with a haploid
gameteforming gametophyte alternating with a diploid sporeforming sporophyte.
However, whereas in the liverwort the gametophyte is the 'dominant' stage, in the
fern this role is taken by the sporophyte.
Fig 10.4.1
142
Chapter 10 – Reproduction in Plants
Fig 10.4 Reproduction in a fern
143
Chapter 10 – Reproduction in Plants
Fig 10.5 Life cycle of a fern
SEXUAL REPRODUCTION IN FLOWERING PLANTS
10.4 The structure of a flower
Flowering plants belong to the phylum Angiospermophyta. They differ from
all other kinds of plants in having true flowers.
Fig. 10.6 shows the structure of a generalised flower and also a flower
belonging to the Papilionaceae. The different parts of the flower are arranged in
rings, sometimes called whorls, attached to the top of the flower stalk or receptacle.
These parts are always arranged in the same order, as shown in Fig. 10.6.
The outer whorl is called the calyx and is made up of sepals. In many
flowers these are dull in colour, and their main function is to protect all the other
parts of the flower while it is still a bud. The next whorl is the corolla, made up of
petals. In insect-pollinated flowers the petals are often brightly coloured and
scented, as their function is to attract insects to the flower. The petals advertise the
presence of the flower from a distance, and the colour and scent of the petals may
determine the kind of insect or other animal which is attracted. In tropical countries,
for example, many flowers are pollinated by birds and these often have bright red
flowers. The dark reddish-brown, strangely marked flowers of Stapelia smell like
rotten meat, and attract flies. In many flowers, nectaries secrete a sugar-rich fluid
called nectar, again in order to attract insects for pollination.
The next whorl is made up of the male parts of the flower and is called the
androecium. It contains several often very many - stamens, each of which has a
stalk called a filament supporting an anther. Inside the anthers, the male gametes
are formed inside pollen grains.
Finally, in the centre of the flower, the female parts or carpels are found.
These make up the gynaecium. There are one or more ovaries, each containing one
or more ovules, inside which the female gametes develop inside an embryo sac. At
the top of each ovary is a style which supports a stigma. The stigma has the
144
Chapter 10 – Reproduction in Plants
function of capturing pollen grains, one of the first stages in the series of events
which will bring the male gametes to the female gametes.
Fig 10.6 –The structure of a flower
10.5 Formation of pollen grains
Male gametes are produced inside pollen grains. Pollen grains are formed
inside the anthers. Each anther contains four compartments called pollen sacs. The
wall of each pollen sac contains several layers of cells. One of the outer layers is
made up of cells with thickened walls and is called the fibrous layer; this helps to
liberate the pollen grains when they are ripe. The innermost layer is called the
tapetum. The cells in this layer help to provide nutrients to the developing pollen
grains.
In the centre of each pollen sac, diploid pollen mother cells divide by
meiosis, each producing four haploid cells. In some species, these stay together in a
group of four called a tetrad, but in others they separate. Each of the haploid cells
develops a tough protective wall around itself, becoming a pollen grain. The wall is
made up of two layers, an outer very tough, waterproof exine and an inner intine.
In place, the exine is absent, leaving a thin area in the wall called a pit. The form
and structure of the exine varies from species to species, and it is possible to identify
a plant just by looking at its pollen grains. They often have spikes or knobs to help
them to stick to the bodies of insects.
145
Chapter 10 – Reproduction in Plants
The haploid nucleus inside each pollen grain divides by mitosis, forming two haploid
cells separated by a very thin cell wall. One of these haploid nuclei is called the
generative nucleus, and the other is the tube nucleus.
When the pollen grains are fully formed, the anthers spilt open in a process called
dehiscence. They split along a line between the two pollen sac on either side
(Fig.10.8), exposing the pollen grains on the surface. Before following the pollen
grains further, we will look at how the female gamete- forming structures develop to
a similar stage.
146
Chapter 10 – Reproduction in Plants
Fig10.8- the production of pollen in an anther
147
Chapter 10 – Reproduction in Plants
10.6 Formation of embryo sac
The female gametes are produced inside structures called embryo sacs which
develop inside the ovules (Fig 10.9)
Ovules are formed inside ovaries. Each ovule is connected to the ovary by a
stalk called the funicle. The ovule has an outer covering, or integuments,
surrounding a tissue made up of relatively undifferentiated cells called the nucellus.
At one end of the ovule, the integuments do not quite meet, leaving an opening
called the micropyle. The other end of the ovule, furthest from the micropyle and
nearest to the funicle, is called the chalaza.
Inside each ovule, a large, diploid, spore mother cell develops. This cell
divides by meiosis to produce four haploid cells. All but one of these degenerates and
the one surviving haploid cell then develops into an embryo sac.
The embryo sac absorbs nutrients from the nucleus and grows larger. Its
nucleus divides by mitosis three times, forming eight haploid nuclei. Two of these
nuclei are found near the centre of the embryo sac, with no cell membranes around
them. The other six arrange themselves at the ends of the embryo sac, three at one
end and three at the other, and usually develop cell membranes. The three haploid
cells at the end nearest the chalaza are called antipodal cells. One of the haploid
cells at the end nearest to the micropyle is a little larger than the other two. This is
the female gamete, the egg cell. The other two cells at this end are called
synergids. The two nuclei in the middle may fused together to form a single diploid
nucleus called the primary endosperm nucleus. Thus, a mature embryo sac
usually contains six haploid nuclei and one diploid nucleus.
10.7 Pollination
Pollination is the transfer of pollen from the anther, where it was made, to a stigma.
Many species of plants have mechanisms which ensure that the pollen is transferred
to a different individual of the same species; this is called cross pollination. In
other species, such as the garden pea, it is usual for pollen to be transferred to the
stigma of the same flower; this is called self pollination.
As neither plants nor pollen grains can move actively from place to place, other
agents are used to transfer the pollen grains. These include animals, such as
insects, birds, bats and other small mammals such as mice, and the wind. A few
aquatic plants make use of water currents to transfer pollen. We will look at how
pollination happens in an insect-pollinated flower and a windpollinated flower.
Insect-pollinated flowers attract insects by providing a 'reward' for them when
they visit the flower. This reward is often either carbohydrate-rich nectar or proteinrich pollen, or sometimes both. The flower advertises the presence of these foods
with brightly coloured petals and/or a scent. The petals are frequently arranged to
provide a landing platform for insects. When the insect arrives at the flower, it
brushes against the anthers as it collects its 'reward'. Some of the pollen grains stick
to its body. The insect flies away, and will often go straight to another similar flower.
Here, some of the pollen grains may brush off its body onto the stigma.
148
Chapter 10 – Reproduction in Plants
149
Chapter 10 – Reproduction in Plants
Fig 10.9 – The development of the embryo sac
It is clear that both the plant and the insect benefit from this arrangement. Insects
and flowers have evolved together over millions of years, and this has resulted in
some very elaborate pollination mechanisms. Some flowers, such as orchids, have
such complex pollination mechanisms that they can only be pollinated by one species
of insect.
150
Chapter 10 – Reproduction in Plants
Wind-pollinated flowers have no need to attract insects, so they do not waste
resources in producing large, brightly coloured petals or nectar. They are usually
relatively small flowers, and are often held on long stalks so that they can easily
catch the wind. Fig. 10.11 shows the structure of a typical grass flower. The
filaments are long and dangle freely out of the flower; they are very flexible, and
move easily in the wind, shaking the pollen free from the anthers. Huge quantities of
tiny, light pollen grains are produced, and you can often see clouds of this pollen
floating up from flowering grasses if you brush against them. (Grass pollen is one of
the main culprits causing hay fever.) The stigmas are long and feathery, and
protrude from the flower. Their large exposed surface increases their chance of
catching pollen grains floating on the wind.
Fig10. 11- Structure of a wind-pollinated flower
10.8 Fertilisation
Pollination results in the arrival of pollen grains on to the stigma. The
pollen grains stick to the surface of the stigma, absorb water and begin to
germinate. This normally only happens if the pollen grain is on the stigma of
the same species of flower, and - in some species - if it is on the stigma of a
different flower of the same species.
The contents of the pollen grain push out through one of the pits in the wall,
forming a pollen tube (Fig. 10.12). The tube grows down through the style
towards an ovule. The tube nucleus remains close to the tip of the tube as it
makes its way through the style. Digestive enzymes are secreted from the tip
of the tube, which is probably directed towards the ovule by chemicals which
the ovule secretes. As the tube grows, the generative nucleus divides by
mitosis, forming two haploid male gametes.
In most plants, the pollen tube enters the ovule through the micropyle,
although in a few species it may digest its way in through the chalaza. Once,
151
Chapter 10 – Reproduction in Plants
the tube has penetrated the ovule, the tube nucleus degenerates as its role is
completed. Behind it, the two male gametes make their way into the embryo
sac.
As you would expect, one of the male gametes fuses with the egg cell, forming a
diploid zygote. But, in plants, a double fertilisation takes place inside the embryo
sac. The other male gamete fuses with the diploid nucleus in the centre of the
embryo sac, forming a triploid nucleus (that is, possessing three sets ofchromosomes). This triploid nucleus is called the endosperm nucleus.
Wind
1.
2.
3.
– pollinated flower
Generally small
Petals green or dull coloured
Do not produce nectar
4. Flower hangs down for easy
shaking
5. Stamens and stigma hang out of
the ring of petals
6. Large number of pollen grains
produced
7. Pollen grains very light with smooth
surface
8. Stigma has feathery branches for
catching pollen
Insect- pollinated flowers
Generally larger
Petals often brightly coloured
Petals have nectarines which produce
nectar
Flower faces upwards
Stamens and stigma inside the ring of
petals
Smaller number of pollen grains produced
Pollen grains heavier with spikes for
sticking to insect
Stigma is like pinhead and lacks
branches.
152
Chapter 10 – Reproduction in Plants
Fig 10.12 - Fertilization
153
Chapter 10 – Reproduction in Plants
Fig10. 14- The development of the seed
Development of a seed
Once fertilisation has taken place, the ovule is called a seed. It remains attached to
the parent plant and continues to receive nutrients from it as it develops. Fig. 10.14
154
Chapter 10 – Reproduction in Plants
shows how a seed develops.
The diploid zygote divides by mitosis to form an embryo plant, attached to the
wall of the ovule by a large basal cell and a little column of cells called the
suspensor. The embryo develops a radicle or embryo root, a plumule or embryo
shoot, and two cotyledons. In some species, the cotyledons will become the first
leaves, the 'seed leaves', of the young plant when the seed germinates, while in
others the plumule is the first part to reach the light and photosynthesise. In many
species, the cells of the cotyledons build up large stores of food within the seed, such
as starch, which will be used by the embryo in the early stages of germination.
The triploid endosperm nucleus also divides by mitosis, forming a tissue called the
endosperm which surrounds the developing embryo. The function of the endosperm
is to provide nourishment for the embryo. In some seeds, the endosperm has
completed its function within a few days of fertilisation and disappears. In others,
such as cereal grains it remains as the main storage tissue to provide nutrients
during germination.
While all this is happening, the integuments of the ovule are developing into the
testa of the seed. This involves thickening and toughening, as waterproof
substances such as lignin are laid down in the cell walls. The small gap in the
integuments, the micropyle, remains as a tiny hole in the testa
The wall of the ovary also undergoes changes after fertilisation. The ovary
becomes a fruit, and its wall becomes the pericarp of the fruit. The seeds are, of
course, contained within the fruit, and the fruit is often adapted to disperse the
seeds away from the parent plant. The pericarp may become fleshy and sweettasting to attract animals, or it may develop hooks and spines to stick to their hair.
Wind-catching projections may form, or the fruit may become dry and hard, later
splitting forcefully and throwing the seeds in all directions. In many species, other
parts of the flower are involved in these developments; in dandelions, for example,
the calyx becomes the 'parachute' of the fruit. In general, however, the various parts
of the flower have completed their role by now, and they wither and fall off as the
seeds develop inside the fruits.
10.10 Seed dormancy and germination
In some species of plants, such as the common garden weed groundsel, the
seeds are ready to germinate as soon as they leave the parent plant. More
commonly, the seeds are in a state of suspended growth, called dormancy, when
they are shed. Towards the end of their development, while still attached to the
parent plant, water is withdrawn from them until their water content is as little as
5% of that of other plant tissues. This dryness helps seeds to survive for many years
in very hostile conditions, such as drought, or extreme heat or cold. The relative
concentrations of hormones, such as ABA, gibberellin and cytokinin, are also thought
to keep the seed in a state of dormancy.
The exact conditions required for a seed to break dormancy and begin to
germinate differ from species to species. Some require particular types of light;
almost all require exposure to a particular range of temperature, an adequate supply
of oxygen, and water. As germination begins, water is absorbed into the seed and
hydrolytic enzymes, such as amylases, break down the food stores into smaller
soluble molecules. These are transported to the developing embryo in which the
nutrients are used to supply materials and energy to allow the cells to divide and
lengthen.
155
Chapter 10 – Reproduction in Plants
First the ridicule and then the plumule - or the cotyledons followed by the
plumule - emerge from the seed. The radicle is positively geotropic and grows
downwards. It can then absorb more water from the soil, as well as inorganic ions,
allowing the rest of the seedling to grow even more rapidly. The plumule is
negatively geotropic and positively phototropic, and grows up above the soil.
Up until this point, all of the growth of the seedling depended on the use of
nutrients such as starch and protein which were stored in either the cotyledons or
endosperm. Some of these nutrients are used to form new molecules and structures
such as cellulose in cell walls. Some of them are respired, at first anaerobically
(because oxygen cannot easily penetrate the seed) and then aerobically, so that
some of the carbon atoms from them are released as carbon dioxide. The dry mass
of a germinating seedling therefore drops during these early stages of germination.
Once the plumule gets above ground, however, photosynthesis can begin. Now
carbon atoms from carbon dioxide in the air are fixed and become incorporated into
new molecules in the plant, so its dry mass begins to increase (Fig. 10.15).
Fig10.15 changes in dry mass during germination of a seed
156