Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
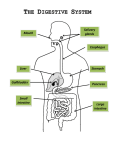
Small bowel physiology Physiology: Small bowel function As chyme enters the small intestine, the intestinal phase of digestion begins. There are some reflexes regulating acidic chyme delivery rate from the stomach to the intestine. The presence of acidic chyme in the duodenum releases the hormone secretin. Secretin inhibits acid production and gastric motility slowing gastric emptying. In addition, secretin stimulates production of pancreatic HCO3 to neutralize acidic chyme that has entered the intestine. If a meal contains fat, the hormone cholecytokinin (CCK) is secreted into the bloodstream. CCK also slows gastric motility and acid secretion. Fat digestion is slower than protein or carbohydrate digestion so it is critical that the stomach only allow small amounts of fat into the intestine at one time. If the meal contains carbohydrates then the hormone GIP is released. Large doses of GIP inhibit gastric acid secretion. The mixture of acid, enzymes, and digested food in chyme usually forms a hyperosmotic solution. Osmoreceptors in the intestinal wall are sensitive to the osmolarity of the entering chyme. When inhibited by high osmolarity, they inhibit gastric emptying in a reflex mediated by some unknown blood born substance. The net result of all three phases of gastric function is the digestion of protein in the stomach by pepsin, the formation of chyme by the action of pepsin, acid peristaltic contractions and controlled entry of chyme into the small intestine so that further digestion and absorption can take place. Most digestion and absorption occurs in the intestine. As chyme from the stomach passes through the small intestine, additional enzymes and fluid are added by the intestinal epithelium, the exocrine portion of the pancreas and the liver. Most digestion and absorption take place in the duodenum and jejunum. The small intestine and its accessories fulfill these roles. Bicarbonate secretion, primarily by the pancreas, neutralize acidic chyme to prevent damage to the intestinal mucosa. Intestinal goblet cells secrete mucus for protection and lubrication. The pancreas and intestinal epithelium secrete fluid containing enzymes for the digestion of proteins, carbohydrates and fats. The liver adds bile a complex mixture that contains wastes to be excreted and agents to aid fat digestion. Motility in the intestine mixes chyme with enzymes and exposes chyme to the mucosa for absorption. Forward movement through the intestine must be slow enough to allow digestion and absorption to go to completion. Absorption in the intestine takes place through transporting absorptive cells called enterocytes. Most digested nutrients are absorbed into the blood of the hepatic portal system. This specialized region of the circulation has two capillary beds: one set to pick up absorbed nutrients at the intestine and the other set to deliver the nutrients directly to the liver. Digested fats, however, go into the lymphatic system rather than into the blood because the intestinal capillaries have a basement membrane around them that most lipids are unable to cross. Endocrine secretions from the intestine coordinate gastric emptying. Other endocrine and neural signals stimulate enzyme and bicarbonate secretion or release of bile from the gall bladder. GIP acts in a fashion to initiate insulin release from the endocrine pancreas. Motility Once food enters the small intestine, it is slowly propelled forward by a combination of peristoltic and segmental contractions. Parasympathetic innervation, gastrin, and CCK all promote intestinal motility. Sympathetic innervation inhibits it. Secretion: secretion in the small intestine comes from the intestinal epithelium, the exocrine pancreas, and the liver. Enzyme secretion The pancreas and intestinal epithelium cells secrete enzymes for the intestinal phase. Pancreatic enzymes enter the intestine in a watery fluid and are active in the lumen. Intestinal enzymes, on the other hand, remain bound to the apical membranes of brush border cells, anchored by transmembrane protein “stalks”. Brush border enzymes are stationary and will not be swept out of the small intestine along with the chyme as it is propelled forward. The brush border enzymes include peptidases, disaccharidases, and a protease called enteropeptidase (enterokinase). A variety of enzymes is secreted by exocrine cells into the pancreas, including pancreatic amylase, lipase and several proteases and peptidases. Some of these enzymes are secreted as inactive zymogens that must be activated upon arrival in the intestine. Brush border enteropeptidase converts inactive trypsinogen to active trypsin. Trypsin then converts the other inactive pancreatic enzymes to their active forms. What signals release of the enzymes? The brush border enzymes are bound to the membrane and are always in place. The pancreas’ enzymes respond to distension and the presence of food in the intestine. The pancreatic reflex is mediated by both the nervous system and by the hormone CCK. Bicarbonate secretion: Bicarbonate secretion into the small intestine is essential to neutralize the highly acidic chyme that enters from the stomach. Most of the bicarbonate comes from the pancreas, which secretes a watery solution of NaHCO3. The cells that produce bicarbonate are different than the enzymesecreting cells and they respond to different signals. Bicarbonate secretions are released in response to neural stimuli and the hormone secretion. In the pancreas, as in the renal tubule cells and red blood cells, bicarbonate products require high levels of the enzyme carbonic anhydrous. Bicarbonate produced from CO2 and water is secreted by a ClHCO3 antiport protein into the pancreatic duct for delivery to the small intestine. A basolateral Na-H antiport transports H into the extracellular fluid. The H thus reabsorbed helps balance the HCO3 input into the blood by parietal cells. Sodium secretion in these tissues is a passive process driven by electrochemical gradients. To create this gradient the epithelia cells transport Cl from the extracellular fluid into the lumen. The basolateral membrane contains NA, K, 2Cl symport proteins that bring Cl into the cell. On the apical membrane, a gated channel known as cystic transmembrane regulator or CFTR chloride channel, allow Cl to enter the lumen. The chloride reenters the cell in exchange for HCO3. (20-21) The movement of negative charge from the ECF to the lumen attracts positive Na, which moves down the electrochemical gradient through leaky junctions between the cells. This type of transepithelial movement pattern is called a paracellular pathway. The secretion of Na and HCO3 into the lumen creates an osmotic gradient so water follows by osmosis. The net result is secretion of a watery sodium bicarbonate solution from the pancreas and intestine. Bile secretion Bile is a non-enzyme solution secreted from the hepatocytes or liver cells. The key components of bile are (1) bile salts for fat digestion, (2) bile pigments, such as bilirubin, that are the waste products of hemoglobin degradation, (3) and cholesterol. Bile acids are steroids that combine with amino acids to form bile salts. Bile salts are detergents with polar side chains that allow them to interact with both lipids and water, creating water soluble droplets. Bile is secreted into hepatic ducts that lead to the gall bladder, which shoves and concentrates the solution. During a meal contraction of the gall bladder sends bile into the duodenum through he bile duct along with a watery solution of bicarbonate and digestive enzymes from the pancreas. Gall bladder contraction is stimulated by CCK. Carbohydrate digestion and absorption Carbohydrate digestion is the small intestine converts digestible polysaccharides and disaccharides into monosaccharides so that they can be absorbed. Amylase from pancreas continues to digest starch into maltose. Maltose disaccharides from food, such as sucrose and lactose must then be digested by the brush border disaccharidase (maltase, sucrase and lactase) to complete the digestion process. The end product of carbohydrate digestion is glucose, galactose and fructose. Intestinal glucose and galactose absorption uses the same transporters as the proximal renal tubule. As apical Na – glucose symport protein and a basolateral GLUT2 facilitates diffusion transport. The glucose transporters move galactose as well as glucose. Most fructose absorption on the other hand, is not sodium dependent. Fructose moves by facilitated diffusion on the GLUT 5 transporter. If glucose is the major metabolic substrate for aerobic respiration, why don’t intestinal epithelial cells use the glucose they absorb for their own metabolism? Metabolism of intestinal cells differs from that of most other cells. They do not use glucose as their preferred energy substrate. Current studies suggest that these cells use the amino acid glutamine as their main source of energy, thus allowing glucose to pass unchanged into the blood stream. Protein digestion and absorption Bicarbonate in the intestine neutralizes acidic chyme which increases luminal pH. Pepsin is inactivated at this higher pH, but other proteases from the pancreas and brush border continue to digest protein. The primary products of protein digestion are free amino acids, dipeptides, and tripeptides all of which can be absorbed. There are multiple amino acid transport systems in the intestine. Most free amino acids are carried by Na dependent co-transport proteins similar to those in the proximal tubule of the kidney. A few amino acid transporters are H dependent. Di- and tripeptides are carried into the mucosal cell using H dependent co-transport. Once inside the epithelial cell di and tripeptides have two possible fates. Most are digested by cytoplasmic peptidases into amino acids, which are transported across the basolateral membranes and into the circulation undigested . Di-and tripeptides are transported intact across the basolateral membrane. Some larger peptides are absorbed by transcytosis after binding to a receptor on the luminal surface of the epithelium, Peptide fragments may act as antigens and stimulate antibody formation and result in allergic reaction. Consequently, the intestinal absorption of large peptides may be a significant factor in the development of food allergies and intolerance. Fat digestion and absorption Fats enter the small intestine from the stomach in the form of a coarse emulsion created by acid lipase digestion and mechanical mixing. In the duodenum bile salts coat the coarse emulsion to help stabilize the droplets. The digestion of triglycerides in the emulsion is carried out by pancreatic lipase. Lipase with a protein cofactor co-lipase from the pancreas is able to penetrate the bile salt coating. Co-lipase displaces some bile salts allowing lipase access to fats inside the bile salt coating. Lipase digests triglycerides into monoglycerides and fatty acids. As enzymatic and mechanical digestion proceed, fatty acids, bile salts, monoglycerides and phospholipids and cholesterol form small cylinders called micelles. Fats are absorbed by simple diffusion across the apical membrane of the intestinal cells. At the brush border surface, fatty acids and monglycerides move out of micelles and into the epithelial cells. Cholessterol is transported on a specific energy dependent membrane transporter. Once in the cytoplasm of the epithelial cells, monoglycerides and fatty acids move to the smooth endoplasmic reticulum where they are resynthesized into triglycerides. Triglycerides then combine with cholesterol and protein to form large droplets called chilomicrons Because of their large size, chilomicrons are packaged into secretory vesicles and leave cells by exocytosis. Once in the extracellular space their size prevents them from entering the capillaries. Instead chilomicrons are absorbed into the lymph vessel of the villi called . They pass through the lymphatic system and finally enter the venous system just before it flows into the heart. Bile salts are not altered during fat digestion. In the terminal ileum they are transported intact back into the hepatic portal circulation. This recirculation of bile salts is essential to maintain fat digestion because the body’s pool of bile salts will cycle from two to five times for each meal. Bilirubin is not reabsorbed and passes into the large intestine for excretion. Vitamins and Minerals In general the fat soluble vitamins (A,D,E and K0 are absorbed along with fats. The water soluble vitamins are absorbed by mediated transport. The major exception is vitamin B12 also known as cobalamin. The intestinal transporter for B12 recognizes the vitamin only when it is complexed with intrinsic factor. This transporter is found only in the ileum. Mineral absorption usually occurs by active transport. Iron and calcium are two of the few substances whose intestinal absorption is linked to their concentrations in the body. Absorption of ions and water About 5.5 liters of fluid and food per day enter the small intestine. Another 3.5 liters of pancreatic and intestinal secretions are added for a total of 9 liters in the lumen. Of this total, most (7.5 liters) is reabsorbed from the small intestine. The absorption of organic nutrients which takes place mostly in the duodenum and jejunum creates an osmotic gradient for water absorption. Na is actively reabsorbed across the intestinal epithelium using transporters similar to the kidney. One apical transporter carries Cl and Na. Another is a Na-K-2Cl symporter that brings all three types of ions into the cell junctions in the small intestine. K and Cl also pass through the cell junctions By the end of the ileum about 1.5 liters of unabsorbed chyme remains. It passes into the large intestine though the ileocecal valve.