Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
Ecological fitting wikipedia , lookup
Introduced species wikipedia , lookup
Overexploitation wikipedia , lookup
Unified neutral theory of biodiversity wikipedia , lookup
Island restoration wikipedia , lookup
Occupancy–abundance relationship wikipedia , lookup
Latitudinal gradients in species diversity wikipedia , lookup
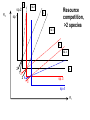
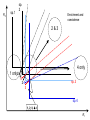
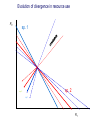
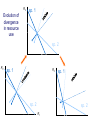
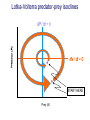
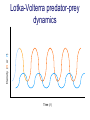
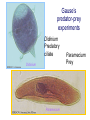
Resource competition among >2 species • One resource – species with lowest R* excludes all others – example: species 1 excludes all others Resource competition among >2 species • Two resources, essential • Constant, homogeneous environment • Two resources - two coexisting species at equilibrium – which two species depends on resource ratios – each species is best competitor for a particular ratio of resources 1 R2 1&2 sp.2 sp.1 Resource competition, >2 species 2 2&3 3 3& 4 2 4 13 24 3 sp.3 sp.4 R1 New effect: spatial variation • Suppose resource ratios vary locally – natural heterogeneity in soil nutrients – consequences for coexistence? • When there is local spatial variation in resource ratios, >2 species can coexist – with local spatial segregation (patchiness) – More species than resources R2 Variation in resource ratios sp.2 sp.1 2 13 24 3 sp.3 sp.4 R1 Spatial variation • • • • Local variation fosters diversity More species than resources possible Dependent on extent of variation Plant communities – often 100’s or 1000’s species – only about 12 essential resources – often patchy • Variance in Resource Ratios Hypothesis (VRR) What is the effect of nutrient enrichment? • Relationship of diversity & productivity • Unimodal vs. Monotonic • Mechanisms producing relationships – Unimodal – particularly decrease in diversity with productivity R2 sp.1 sp. 2 Enrichment and coexistence 2&3 4 only 1 only2 13 24 3 sp.3 sp.4 R1 Nutrient enrichment • Increase all resources uniformly – local variation in resource ratios allows coexistence of fewer species • Increase one resource – necessarily makes resource ratios more extreme – raises, then lowers number of coexisting species • Assumes resources increase without increasing variation • “Paradox of enrichment” – enrichment = reduced diversity Switching resources • Does VRR predict coexistence of many species on 2 switching resources? – species don’t specialize on ratios – each species consumes one resource or the other only • At equilibrium there are 2 species, each consuming and limited by one resourse – Fundamental difference between animal and plant communities Switching resources R2 sp.4 1&4 sp.3 sp. 2 4 sp.1 1 R1 Plants vs. Animals • Plants use essential resources • VRR predicts high species:resources ratio • Animals use switching resources • Theory predicts species:resources ratio =1 Coexistence and evolution • Competitive coevolution – – – – 2 spp. competing for 1 resource cannot coexist if individuals vary in resource use if that variation is heritable competition creates selection • May select for increasing efficiency – selection for better resource use (lower R* ) – a “race” to be most efficient – end result is still exclusion Coexistence and evolution • Competition may select for divergence in resource use – individuals exploiting an alternative resource favored (not affected by competition) – alternative resources could be different spatially, temporally, in size – for substitutable or switching resources – evolution of divergence may avoid exclusion Example: Divergence in prey size freq. of use selection against time freq. of use size of prey size of prey Evolution of divergence in resource use R2 sp. 1 sp. 2 sp. 2 sp. 1 R1 R2 Evolution of divergence in resource use sp. 1 sp. 2 R2 R2 sp. 1 sp. 2 sp. 1 sp. 2 R1 Competitive character displacement • Competition selects for divergence in a morphological feature – presumably results in divergence of resource use – often held to be the best evidence for the importance of competition – Example: Sitta nuthatches Nuthatches – Example: Sitta nuthatches – Asia & Europe – Ranges include regions of allopatry (no contact) – also regions of sympatry (co-occur) – Sitta neurenmayer (Europe) – Sitta tephronata (Asia) – Sympatry in Iran Nuthatches • Bill size – related to prey size – data suggest character displacement on bill size • S. neurenmeyer tephronta • Allopat. 25 mm • Sympat. 22 mm S. 25 mm 28 mm Prediction of character displacement bill length (mm) S. tephronata S. neurenmayer site (longitude) Actual pattern (Grant 1972) bill length (mm) S. tephronata S. neurenmayer site (longitude) Nuthatches • No shift in cline of bill size when region of sympatry is reached • Bill sizes vary geographically in a continuous fashion • Not much evidence for character displacement Hydrobia snails • intertidal mud snails – particle feeders (diatoms, sediment) • Allopatry – H. ventricosa – H. ulvae mean length = 3.1 mm mean length = 3.3 mm • Sympatry – H. ventricosa – H. ulvae mean length = 2.8 mm mean length = 4.5 mm Hydrobia snails Questions • Character displacement? • Competition for food particles? • Levinton - does particle size affect growth? – larger species does best on larger particles? • Result: No difference in growth for different particle sizes Hydrobia snails: More questions • H. ulvae & H. ventricosa sympatric in lagoons • H. ulvae alone in intertidal • Lagoon H. ulvae – alone … 1.2 X larger than intertidal H. ulvae – w/ H. ventricosa … 1.4 X larger than intertidal H. ulvae • size difference due to physical environment? • lagoons: low reproduction, high growth Character displacement • Classic cases of character displacement now questioned • Probably not a widespread phenomenon • Morphology (size) presumed related to resource use • Competition presumed to be the driving force • Examples of size differences reducing competition? Caribbean Anolis Pacala & Roughgarden 1985 • St. Maarten • A. gingivinus – SVL = 41 mm • A. wattsi – SVL = 38 mm • St. Eustatius • A. bimaculatus – SVL = 53 mm • A. wattsi – SVL = 40 mm Caribbean Anolis • Predict less competition on St. Eustatius • Note: size strongly correlated with prey size Experiment 12 X 12 m enclosures; fenced 1.5 m; clear lizards 60 Ag 100 Aw 60 Ag 60 Ab 100 Aw 60 Ab 60 Ag 100 Aw 60 Ag 60 Ab 100 Aw 60 Ab St. Maarten St. Eustatius Caribbean Anolis • St. Maarten • A. gingivinus + A. wattsi – less food in stomach – lower growth rate (0.5X) – perch height higher (2X) • St. Eustatius • A. bimaculatus + A. wattsi – same amount in stomach – same growth rate – same perch height • compared to A. gingivinus • compared to A. bimaculatus alone alone • Interspecific effect strong • Interspecific effect absent Alternative interpretation • Suppose competition is absent on St. Eustatius – large resource base, abundant food – predators reduce density • A. bimaculatus enclosures – escapes occurred over time – density: 60 45 30 lizards – 1 mo 2 mo – as density drops growth increases; competition Conclusion • Size difference reduced competition • One case, but it shows this effect is possible • Authors do NOT claim size difference evolved due to competition • Has not established that size would evolve in response to competition Morphological evolution & competition (Schluter 1994) Sticklebacks • species complex • extreme body forms Representative limnetic (top) and benthic (bottom) stickleback from Lake Enos in British Columbia, Canada. Click to enlarge. Posted with permission from Paul J. B. Hart and Andrew B. Gill, "Evolution of Foraging Behaviour int the threespine stickleback," in The Evolutionary Biology of the Threespine Stickleback, eds. Michael A. Bell and Susan A. Foster, (Oxford: Oxford University Press), 1994, p. 211. © Oxford University Press – limnetic - feed on plankton (e.g., Daphnia) – benthic - feed on benthic invertebrates see also Robinson & Wilson 1994 Sticklebacks • Morphological intermediates exist • 1 sp. in a lake -- typically intermediate morph • 2 spp. in a lake -- typically 2 morphs • Morphology is related to feeding efficiency and growth • Hypothesis: evolved morphological divergence due to competition (Character displacement) Experiment • 23 X 23 m ponds • Target species intermediate in morphology • produced by hybridization intermediate X intermediate intermediate X limnetic intermediate X benthic Morphology Hypothesis • Competition with a limnetic will have greatest effect on survival and growth of forms morphologically similar to limnetic Target Limnetic Morphology Target Limnetic Morphology Experiment • Hybrids add variation on which selection can work intermediate X intermediate intermediate X limnetic intermediate X benthic Morphology Implication • If hypothesis is supported, selection for character divergence is occurring via competition Experiment Experimental 1800 target 1200 limnetic Control 1800 target X 2 ponds Data collection • 3 months • Collect fish, measure Target • Growth rate reduced by density – competition occurs • Regression of growth vs. morphology • Slope = growth differential between more benthic and more limnetic Results Control Competitor IxB IxI IxL morphology Results • Growth differential – significant for 1 experimental group – nearly so for a 2nd experimental group – clearly not significant for both controls • Survival differential – some evidence for an effect in 1 pond • Target individuals with limnetic morphology fare worst Conclusions • Experimental evidence for character displacement • Caveats: – pseudoreplication – statistical weakness Lake whitefish Coregonus lavaretus dwarf, limnetic benthic Null models in community ecology • Experiments – show that a process occurs – may show it can cause effects on distribution, abundance, fitness of a limited set of species – Does that process structure the community as a whole? – experiments rarely can test that • If interspecific competition is important, what patterns would be predicted for communities? Community patterns • Competition favors differences in resource use among co-occurring species • Predict: co-occurring species should be more different in resource use than expected if species were placed together randomly. • Should be present across similar species within a community G. E. Hutchinson • Co-occurring European Corixids • Body lengths – ratio of larger to smaller tended to be >1.3 • Morphology as a surrogate for resource use • Origin of idea of limiting similarity Morphological pattern • Predict: co-occurring species should be more different in morphology than expected if species were placed together randomly. • "Community-wide character displacement" • How do you tell? • Null models or Neutral models of communities • Morin 98-103; Chase & Leibold 117-122 Statistical Null hypotheses • Hypothesis of only chance affecting outcome • e.g., c2 for mendelian assortment – coat color… Red – RR • • • • White rr Roan Rr Cross two Roan: Rr x Rr Expect: RR = 0.25; Rr = 0.50; rr = 0.25 observe: RR = 0.26; Rr = 0.38; rr = 0.36 c2 = 7.76, P<0.05 … significant departure from (null) expectation Statistical Null hypotheses • Expected: assumption of random sampling of alleles • P<0.05: results deviating as far (or farther) than observed expected <5% of the times if only random processes are involved • conclude: some non-random process is structuring alleles at this locus • Same general pattern in community ecology, but the model and math are more complex Example – Dytiscid beetles • • • • (Juliano & Lawton 1990) 28 species, Northern England 9 different sites have 8 to 16 species interspecific variation in size and shape Are co-occurring species more different in morphology than expected? Hyphydrus ovatus Hygrotus inaequalis Hydroporus planus Issues for null models • What is the character of interest? – Resource use – Morphology • one variable • many variables • correlation of variables – Co-occurrence (do pairs of species co-occur less often than expected … “forbidden combinations”) Issues for null models • What is the source pool of species? – Islands • Mainland fauna • All species on similar islands – Limits of source pool • Taxonomic • Geographic • Trophic Issues for null models • What is the source pool of species? – Real species (discrete values) • Randomization tests – Statistical distributions (continuous) • Monte Carlo methods; simulations Issues for null models • Identifying the assemblage present – presence/absence – abundance • rare species may transients, not integrated into the community • rarity may be a result of inappropriate morphology or resource use Issues for null models • Test statistic – measure of differences – Size ratios (Univariate only) – Morphological nearest neighbor distance – Minimum spanning tree • Mean vs. Variation – predict mean difference larger than expected – predict variation of difference smaller than expected (regularity of species spacing) – combination Issues for null models • Constraints on randomization – Stratify by other factors, e.g., genera within families – Overall distribution – widespread species more likely to be included – Dispersal ability – good dispersers more likely to be included Source pool : The narcissus effect • Colwell & Winkler 1984 • What if assemblages at all locations are affected by competition – morphologies are more distinct than expected – randomly draw real species … that effect is incorporated into randomly drawn assemblages – real assemblages do not differ from randomly drawn because both include the effect of competition on morphology Source pool issues: The narcissus effect # species # species • Solution? • Synthetic species (unlike any real species, but within the range of variation) • Draw from continuous distributions of morphological variables (match discrete distributions) size size Dytiscid morphology • length, width, depth, head width – correlated in real species • for real species, choose at random, and allocate to community – each species brings correlated morphological measurements • Cannot simply choose length, width, depth, head – omits correlation structure • Canonical discriminant function – produces uncorrelated variables (up to 4) – choose canonical variates Dytiscid beetles Test: randomization • real community with S species – calculate nearest neighbor distance (NND) in morphological space for all species – get mean NND and SD NND • draw S species from pool – calculate NND in morphological space for all species – get mean NND and SD NND • Repeat many (500 or 1000) times • Test stat [Mean NND – SD NND] =D • Is real D large compared to those drawn at random? Test: Monte Carlo • real community with S species – calculate nearest neighbor distance (NND) in morphological space for all species – get mean NND and SD NND • draw S species from distributions of Canonical functions – calculate NND in morphological space for all species – get mean NND and SD NND • Repeat many (500 or 1000) times • Test stat [Mean NND – SD NND] =D • Is real D large compared to those drawn at random? Test statistic • Reject H0 if observed >95% of all others • Result …For one site, there was a significant pattern of large mean NND and large D, but not of small SD NND • Species at one site are more dissimilar than expected by chance – and given average dissimilarity, the are less variable than expected by chance (D) • using synthetic species (vs. real) null hypothesis is rejected slightly more frequently (narcissus effect) Real species Synthetic species Null distribution and real communties Other results • Significant – Hawks (Accipter spp.) – Middle eastern cats – Some tiger beetle (Carabidae) assemblages – Desert Rodents • Not significant – Birds (Tres Marias & Channel Islands) – Most tiger beetle assemblages – Multiple passerine bird assemblages What does it show? • A significant result establishes that there is a pattern, consistent with prediction. • Does not establish what the mechanism is. • Experiments to test mechanisms where patterns exist – e.g., experiments like Pacala & Roughgarden Exploitation mostly predation Exploitation mostly predation • Predator: kills and eats victim • Parasite: lives intimately with victim and usually does not necessarily kill victim • Herbivore/Carnivore distinction not that important for dynamics Exploitation • How does the presence / absence of a predator affect: – species populations – assemblages of prey species – evolution of prey • Does predation contribute to community patterns? Predation & population dynamics • • • • • • Predators eat prey; prey die due to predation How does this affect population dynamics? Lotka-Volterra predator-prey model Starting point N = number in prey population P = number in predator populatio Lotka-Volterra predator-prey • Without predation, prey grow exponentially dN / dt = r1 N • • • • Predation is an increasing function of N & P Effect of predation on prey population = C1 NP C1 is the capture efficiency So, with predation… dN / dt = r1 N - C1 NP Lotka-Volterra predator-prey • Without prey, predators starve to death exponentially dP / dt = - r2 P • • • • Predation is an increasing function of N & P Effect of predation on predator population=C2 NP C2 = product of capture & conversion efficiencies So, with prey … dP / dt = C2 NP - r2 P Lotka-Volterra predator-prey: Equilibrium predictions • At equilibrium • dN / dt = 0 and dP / dt =0 • there is a specific, constant density of predators, above which prey cannot increase • there is a specific, constant density of prey, below which predator cannot increase Lotka-Volterra predator-prey isoclines PREY ISOCLINE PREDATOR ISOCLINE dN / dt = 0 dP / dt = 0 dP / dt < 0 dN / dt < 0 dN / dt > 0 Prey (N) dP / dt > 0 Prey (N) Lotka-Volterra predator-prey isoclines dP / dt = 0 dN / dt = 0 equilibrium Prey (N) Lotka-Volterra predator-prey isoclines START HERE Prey (N) Density (N or P) Lotka-Volterra predator-prey dynamics Time (t ) Predator-prey cycles in real data • Hare & Lynx • What assumptions are built into Lotka-Volterra predator-prey models? Simplifying Assumptions • Simplifying Environmental – Constant in time – Uniform or random in space • Simplifying Biological – – – – Individuals are identical & constant in time Exponential prey growth Prey limited only by predation Predator growth dependent only on predation Explanatory Assumption • Predators and prey encounter each other at random, like bimolecular collisions – Frequency of encounter proportional to product of densities • Individual predator feeding rate increases linearly as N increases – No limit on increase in feeding rate Unrealistic elements • No limits on prey except predation – expect real prey may be limited by food, space, etc. when abundant – upper limit ( K ) for prey even with no predators • Predators do not saturate with prey – expect real predators to hit a maximum number eaten – expect an upper limit for predators with maximal food (KP ) Gause’s predator-prey experiments Didinium Predatory ciliate Didinium Paramecium Paramecium Prey Density (N or P) Didinium - Paramecium predator-prey experiment Paramecium Didinium Time (t ) Gause’s Predator-Prey experiments • • • • • Predator and prey in a simple environment No cycles (stable or otherwise) Predator exterminates prey Predator dies out shortly after Inconsistent with Lotka-Volterra predator-prey models Gause’s Modified Predator-Prey experiments • Regular immigration of Paramecium • Produces cycles of predator & prey • Consistent with Lotka-Volterra predator-prey models? • No – violates simplifying assumptions – prey population now not soley governed by exponential growth and predation Huffaker’s Predator-Prey experiments • Mites – predator Typhlodromus – prey Eotetranychus • on oranges • With oranges evenly spread on a tray – no cycles – prey extinction, then predator extinction Huffaker’s modified Predator-Prey experiments • Add barriers to dispersal • rubber balls, vaseline – cycles • Confirms Lotka-Volterra prediction? • NO – violates simplifying environmental assumption Predator-Prey models & experiments: Conclusions • • • • Lotka-Volterra models are largely inadequate lab systems meeting assumptions -- no cycles Stable oscillations when system is “fixed” Conceptual error: – Design experiments to meet assumptions, then test predictions – Don’t manipulate experiments until they confirm theory Improved Predator-Prey models • Self limitation of prey and predators • Asymptotic prey consumption by predators • Spatial refuges for prey • graphical approach – Rosezweig & MacArthur (1963) • mathematical approach – Williams (1980) Grover (1997) – Gilpin & Ayala (1973) Populus 5.4