Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Visual selective attention in dementia wikipedia , lookup
Premovement neuronal activity wikipedia , lookup
Biology of depression wikipedia , lookup
Executive functions wikipedia , lookup
Synaptic gating wikipedia , lookup
Environmental enrichment wikipedia , lookup
Affective neuroscience wikipedia , lookup
Neuroplasticity wikipedia , lookup
Emotional lateralization wikipedia , lookup
Human brain wikipedia , lookup
Aging brain wikipedia , lookup
Neuroanatomy of memory wikipedia , lookup
Cognitive neuroscience of music wikipedia , lookup
Time perception wikipedia , lookup
Orbitofrontal cortex wikipedia , lookup
Neuroesthetics wikipedia , lookup
Anatomy of the cerebellum wikipedia , lookup
Neuroeconomics wikipedia , lookup
Neural correlates of consciousness wikipedia , lookup
Cortical cooling wikipedia , lookup
C1 and P1 (neuroscience) wikipedia , lookup
Eyeblink conditioning wikipedia , lookup
Feature detection (nervous system) wikipedia , lookup
Prefrontal cortex wikipedia , lookup
Posterior cingulate wikipedia , lookup
Insular cortex wikipedia , lookup
Motor cortex wikipedia , lookup
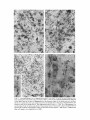
BRAIN RESEARCH 271 R E P R E S E N T A T I O N OF C E N T R A L VISUAL FIELDS IN P R E S T R I A T E C O R T E X OF M O N K E Y S. M. ZEKI* National Center for Prevention and Control of Alcoholism, National Institute of Mental Health at St. Elizabeths Hospital, Washington, D.C. (U.S.A.) (Accepted January 7th, 1969) INTRODUCTION In 1941, Talbot and Marshall 16 reported the manner in which the visual fields are represented in striate cortex of monkey. From this study, later confirmed by Daniel and Whitteridge 6, it appeared that the vertical meridian of the lower and upper visual fields is represented at the striate prestriate boundary behind the lunate and above the inferior occipital sulci respectively with the foveal representation occupying a circular area below the tip of the lunate sulcus and the representation of the horizontal meridian extending backwards from the 'foveal cortex' (see Fig. 10). Since that time the projections of the striate cortex have been studied by Myers lz and by Kuypers et al. 9. Although in both studies the Nauta staining technique was used, the results obtained are not strictly in accord 19. The many disagreements over the anatomy, extent and functions of the prestriate cortex in the monkey have been reviewed at length elsewhere 19. Here it is sufficient to point out that the disagreements have not only been interdisciplinary but also intradisciplinary. For example, among students of cytoarchitectonics, Lashley and Clark 10 and, to a lesser extent, yon Bonin and Bailey 1 were not able to confirm the subdivision into the areas 18 and 19 proposed by Brodmann ~. Lashley and Clark concluded from their architectonic work that the areas 18 and 19 of Brodmann 3 could not be told apart and that, together with the area 7, they should be considered a single functional area. Von Bonin and Bailey 1 conceded that differences in cytoarchitecture may exist but disagreed with Brodmann 3 on the distinguishing cytoarchitectonic criteria and also found the differences to be too gradual and too 'subtle' to justify subdivision into 2 distinct fields. And although they did propose a subdivision into the 2 fields OA and OB, they were unable to clearly specify the boundary between these 2 fields or the anterior boundary of OA, the authors not being able to find any striking cytoarchitectonic differences posterior to the temporal pole. * Visiting Associate: present address: Department of Anatomy, University of Wisconsin, Madison~ Wisc. 53706, U.S.A. Brain Research, 14 (1969) 271-291 272 s.M. ZEKI The studies of fiber degeneration following lesions in striate cortex of monkey have not given consistent results either. With the Marchi myelin stain, Mettler 11 found a limited projection to part of the cortical field lying anterior to the primary visual cortex. Clark 4, using the same technique, could not trace any fibers beyond the opercular lip and concluded that long association fibers emanating from the primary visual cortex do not exist. More recently, the Nauta technique has been used by both Myers 12,13 and his collaborators and Kuypers and his co-workers 9. Kuypers et al. 9 found a projection from the striate cortex to what they term the 'circumstriate cortical belt'. This includes the lunate sulcus, prelunate gyrus and the posterior bank of the superior temporal sulcus in its upper parts. In the Nauta study of Myers a3, however, not all of the 'circumstriate cortical belt' was found to be recipient of fibers from the striate cortex. Myers 13, therefore, proposed a subdivision of prestriate cortex into 4 areas, depending upon their connections with the striate cortex and with homologous areas in the opposite hemisphere. Finally, in the strychnine neuronographic studies of Von Bonin et al. 2 the prestriate cortex was subdivided into areas 18 and 19, although the area 18 in this study was much larger than the area 18 proposed by Brodmann 3. The criterion in this study was whether the opposite hemisphere was fired after strychninization. But since it would be very difficult to strychninize the depth of the lunate sulcus, it is not clear from this study whether the lunate sulcus is in connection with the opposite hemisphere and therefore to be included in the area 18 or whether it has no connections with the opposite hemisphere and therefore to be included in the area 19. These several discrepant findings and the lack of information on the representation of the visual meridians in prestriate cortex of monkey prompted the present study. To obtain a picture of the manner in which the visual field representation is projected onto prestriate cortex, discrete lesions were placed in striate cortex in the regions where the vertical and horizontal meridians are represented, in the 'foveal cortex' of the striate area and in regions between the representation of the horizontal and vertical meridians. Results obtained necessitated a study of the interhemispheric connections of prestriate cortex and this was achieved by section of the splenium and by making discrete lesions in accessible parts of prestriate cortex. M A T E R I A L A N D METHODS Male rhesus monkeys, weighing from 2 to 3 kg, were used in the present study. For cortical lesions, trephine holes followed by limited craniectomies were made and the lesion executed by gentle subpial suction. Great care was exercised not to involve white matter. The lesions that were made were relatively small so that the blood supply to the white matter was not interrupted to any significant extent. Nissl and silver preparations showed that in most of the animals there was no primary involvement or vascular damage to the white matter. Two monkeys in which the white matter was inadvertently involved are not reported in this study. Because of the manner in which both the lunate and inferior occipital sulci fold deeply underneath the operculum, any damage to white matter might involve fibers emanating from prestriate cortex. Brain Research, 14 (1969) 271-291 C E N T R A L V I S U A L FIELDS IN P R E S T R I A T E C O R T E X 273 For sectioning of the splenium, a hole was drilled in the skull and the bone rongeured above the sagittal sinus, one hemisphere retracted and the splenium sectioned by suction and cutting. In all cases, the dura was reapproximated after the lesion was made, covered by gelfoam and the skin closed. In the case of splenial section, only the hemisphere opposite to the one retracted during surgery was used for histology. Animals that were to undergo commissural section were given 15 ml of mannitol intravenously prior to the operation to induce brain shrinkage and minimize thereby retraction of the hemisphere in gaining access to the corpus callosum, in addition, the monkeys were given 0.5 ml of decadron before and after the operation to prevent postoperative edema. All the animals were maintained on either chloromycetin or penicillin antibiotic for 5 postoperative days. Survival periods ranged from 4 to 15 days with the majority surviving around 8 or 9 days (see under individual sections). The monkeys were perfused with formol saline and the brains removed and kept in fresh 1 0 ~ formalin for periods varying from 6 days to 3 weeks. During this time the brains were photographed. The brains were sectioned horizontally on a freezing microtome at 30 # and 3 sections in every 900 # were saved. The sections were stained by a variant of the Fink-Heimer 7 technique for fiber degeneration and by cresyl violet for Nissl substance. The silver staining was performed in a sieve, as described by Wilson and Cragg ~8. The brains were sectioned horizontally. In such sections the sulci are easily identifiable and in addition the lateral striate prestriate boundary is cut at right angles along most of its length. RESULTS A. Characteristics of the degeneration The degeneration seen in this study is characterized as sparse, moderate or heavy and a picture of each type of degeneration is shown in Fig. 11. individual degenerated fibers, mostly of very fine caliber, could be traced in the cortical layers. The pericellular degeneration was limited to the lower 3 cortical layers and only occasionally could a degenerated fiber be seen in the upper cortical layers. An exception to this picture, however, was the region of the opercular lip at the striate prestriate boundary. In this region, degenerated fibers could be seen in the upper cortical layers, including layer 1. But although the degeneration here was very noticeable, it still remained less than the degeneration in the lower cortical layers of this region. The orientation of fibers deserves some comment. In most regions, the fibers passed in all directions without any suggestion that there was a preference for any particular direction. Fibers that were parallel, perpendicular or oblique to the surface could be observed. Some regional differentiation with respect to orientation could, however, be seen. Thus, in the lateral third of the anterior bank of the lunate sulcus and, to a lesser extent, in the posterior part of the prelunate gyrus, fibers that were oriented perpendicular to the surface were particularly prominent (see Fig. l lD). Brain Research, 14 (1969) 271-291 274 s.M. ZEK~ INFERIOR OCCIPITAL SUPERIOR SULCUS TEMPORAL SULCUS Fig. 1. A composite diagram showing the approximate size and location of the lesions made in the striate cortex. A r o u n d the borders of the lesion, the fibers took p r e d o m i n a n t l y two directions, either vertically towards the white matter or horizontally away from the lesion. The former p r e s u m a b l y represent fibers going to distant cortical fields whereas the latter fibers going to adjoining cortical areas. These horizontal fibers could be traced over a distance of 2-3 m m only. lateral / L U N A T E SULCUS posterior anterior MEDIAL CALCARINE SULCUS PARIETO.OCCIPITAL SULCUS medial Fig. 2. The degeneration picture observed in monkey KS 12 at the level indicated. In this and subsequent figures, the dotting in the cortex gives an approximate picture of the density of degeneration but is not intended to reveal the layers of distribution of the degenerated fibers. The uninterrupted line in this and subsequent figures represents the striate cortex. Brain Research, 14 (1969) 271-291 CENTRAL VISUAL FIELDS IN PRESTRIATE CORTEX 275 B. Lesions in the primary visual cortex Discrete lesions were made in the primary visual cortex of 8 monkey hemispheres. None of these lesions involved white matter. In some monkeys the cortical lesion was fairly superficial, involving layers 1-4, whereas in others the lesions were slightly deeper, involving all 6 layers. There did not appear to be any appreciable difference in the ensuing degeneration. Survival in all monkeys with lesions in the striate cortex was limite0 to 8 or 9 days. For ease of reference, the lesions are shown on a composite diagram (Fig. 1). All the lesions were roughly of the same size, being about 6-7 m m long and about 1.5 m m in width. The map provided by Talbot and Marshall 16 was used as a reference in making these lesions. Monkey K S 12 had a lesion that was parallel and posterior to the representation of the vertical meridian of the lower visual field on the striate cortex. In the prestriate cortex, three areas of degeneration could be seen: one just anterior to the striate prestriate boundary at the opercular lip, another in the lunate sulcus and a third area in the posterior bank of the superior temporal sulcus. The degeneration at the striate prestriate boundary was moderate and occupied all the cortical layers. The moderate area of degeneration in the lunate sulcus occupied the medial one-sixth of the anterior bank of the lunate sulcus in more dorsal sections and as the degeneration was traced ventrally, it came to occupy the bottom of the sulcus. In the whole region of degeneration, there was one focus of maximal degeneration surrounded by sparser degeneration in all directions. This picture of maximum degeneration surrounded by sparser degeneration was seen in all areas of degeneration in all the monkeys studied. A third area of degenerated fibers was evident on the posterior bank of the superior temporal sulcus, roughly halfway down the middle. The degeneration in both the lunate sulcus and the superior temporal sulcus was limited to the lower 3 cortical layers. The actual area of distribution of degenerated fibers in all 3 areas was of the order of twice the size of the lesion itself. In monkey K S 21 a lesion was attempted in the representation of the horizontal meridian of the striate cortex, as determined from the Talbot and Marshall in map. The anterior tip of the lesion approached the vertical meridian representation, the lesion having been placed slightly above the representation of the horizontal meridian. The degeneration was almost entirely limited to the posterior bank of the lunate sulcus (Fig. 3). There was a single area of moderate to heavy degeneration occupying the middle third of the posterior bank of the lunate sulcus, flanked on either side by sparser degeneration. When more ventral sections were examined, this area of degeneration moved more medially until in the most ventral sections in which degeneration could be seen, the area of degeneration had shifted almost to the bottom of the lunate sulcus. In addition, at this level, moderate degeneration appeared at the posterior bank of the superior temporal sulcus. The posterior tip of the lesion in this animal was situated in the representation of the parafoveal visual field. The ensuing degeneration suggests that the horizontal meridian is represented in the posterior bank of the lunate sulcus, roughly in the middle, Brain Research, 14 (1969) 271-291 276 s.M. ZEKI Ks21 A. B. Fig. 3. The degeneration picture observed in monkey KS 21 at the levels indicated. and that more medial areas in the depth of the lunate sulcus lead towards the representation of the vertical meridian in the lunate sulcus, as shown by monkey KS 12. The horizontal meridian is represented, therefore, between the two representations of the vertical meridian, one at the striate prestriate boundary at the opercular lip and another in the medial one-fourth of the lunate sulcus, ls is not clear from these 2 monkeys how the horizontal meridian is represented between the vertical meridian representation in the lunate sulcus and that at the posterior bank of the superior temporal sulcus. In addition, it should be noted that there was no degeneration whatsoever in the inferior occipital sulcus, that is to say, the prestriate cortex lying anterior to striate cortex that is related to the upper visual fields. In the next monkey, KS 22, a lesion was attempted in the area of representation KS 22 INFERIOROCCIPITALSULCUS , SUPERIOR ~ ~TEMPORAL SULCUS L \\ \ CALCARINESULCUS CALCARINE SULCUS Fig. 4. The degeneration picture observed in monkey KS 22 at the levels indicated. Brain Research, 14 (1969) 271-291 C E N T R A L V I S U A L FIELDS I N P R E S T R I A T E CORTEX 277 of the horizontal meridian. However, in this animal it was desired to place the lesion just below the line given by Talbot and Marshall TM as representing the horizontal meridian. At its anterior end, the lesion approached the representation of the vertical meridian (of the upper visual field). The degeneration in monkey K S 22 was almost entirely limited to the posterior bank of the inferior occipital sulcus. As the sections were traced from ventral to dorsal, the number of degenerated fibers, moderate at first, increased to become very heavy. This single field of degeneration occupied the middle two-thirds of the posterior bank of the inferior occipital sulcus (see Fig. 4). Within this region, however, the density of degeneration at the more medial side was greater than the density of degeneration at the more lateral side. In more dorsal sections, the degeneration became sparser and there was also a shift in the degeneration, the degeneration tending medially towards the bottom of inferior occipital sulcus. In addition, there was an area of moderate degeneration, in more dorsal sections, at the posterior bank of the superior temporal sulcus. Finally, once the lunate sulcus had just appeared, sparse degeneration was evident in the posterior bank of the sulcus which at this level is very shallow (see section A of Fig. 4). Monkey K S 25 had a lesion very similar to monkey KS 22 except that it was more posteriorly placed. The degeneration was exactly similar to the degeneration observed in monkey KS 22 with the following differences : (a) the degeneration was more ventral and (b) the degeneration in the superior temporal sulcus (which becomes anterior at this level) was absent and, instead, there was sparse degeneration in the inferior occipital gyrus. The degeneration observed in monkeys K S 21, KS 22 and K S 25 suggest that projections from striate cortex to prestriate cortex are strictly topographical. Thus, a lesion made just above the representation of the horizontal meridian in striate cortex leads to degeneration in prestriate cortex related to dorsolateral striate cortex (lunate sulcus and the upper parts of the posterior bank of the superior temporal sulcus) and a lesion made below the representation of the horizontal meridian leads to degeneration only in prestriate cortex related to ventrolateral striate cortex (inferior occipital sulcus, lower part of the superior temporal sulcus and the inferior occipital gyrus). In monkey K S 16 a lesion was made well within the area of representation of the lower visual field in the striate cortex (see Fig. 5). The lesion was oblique, with its anterior tip directed towards the representation of the vertical meridian and its posterior tip situated in the parafoveal visual field representation. In addition, there was in this monkey some involvement of prestriate cortex just beyond the striate prestriate boundary and extending some 2 mm anteriorly from that boundary. The dorsalmost section to reveal degeneration showed the degenerated fibers to occupy the prelunate gyrus and extend into the posterior bank of the superior temporal sulcus and anterior bank of the lunate sulcus. This field of degeneration was heaviest along the crown of the prelunate gyrus, tapering off to become sparser along the sulci. Vertically oriented fibers were particularly prominent in this region. A fairly heavy region of degeneration could be seen occupying the middle two-thirds of the posterior bank of the lunate sulcus. Brain Research, 14 (1969) 271-291 278 s.M. ZEK1 Fig. 5. The degeneration picture observed in monkey KS 16 at the levels indicated. In subsequent figures, only the lateral side of the brain is shown. / / / ~" ~ CB: B. C. Fig. 6. The degeneration picture observed in monkey KS 23 at the levels indicated. As the degeneration was traced ventrally through the horizontal sections, the following changes appeared: the degeneration along the crown of the prelunate gyrus was maintained, with the vertically oriented fibers being ever present. This area of degeneration now became separated from the degeneration in the superior temporal sulcus, which became somewhat sparser. It was also separated from the degeneration along the anterior bank of the lunate sulcus which shifted more medially and also became sparser. In the posterior bank of the lunate sulcus, the single area of heavy degeneration was maintained but at more ventral levels another area of degeneration, distinct from the first and moderate, appeared at the bottom of the lunate sulcus. In the most ventral sections that contained degenerated fibers, the moderate degeneration was entirely limited to the depth of the lunate sulcus. The area of degeneration in the Brain Research, 14 (1969) 271-291 C E N T R A L VISUAL FIELDS IN P R E S T R I A T E C O R T E X 279 posterior b a n k o f the superior temporal sulcus was maintained throughout, becoming heavier as more ventral sections were examined*. The lesion in m o n k e y K S 23 was also limited to the area o f representation of the lower visual field in the striate cortex. The lesion was somewhat parallel to the horizontal meridian representation of the striate cortex. Examined f r o m dorsal to ventral t h r o u g h the horizontal sections, the following areas of degeneration appeared: in the most dorsal section to reveal degenerated fibers, moderate degeneration occupied 2 distinct areas in the posterior bank of the lunate sulcus. One area was just beyond the opercular lip, on the lateral side of the posterior bank, while the other was more medially situated. In more ventral sections, the 2 areas o f degeneration coalesced so that a single area o f degeneration was now evident. Within this single field o f degeneration, however, the more lateral third was less heavy than the more medial two-thirds. This picture was maintained until the single field of degeneration separated once more into 2 distinct areas with an intermediate area free of degenerated fibers. The lateral of these 2 areas maintained its position, became sparser and finally disappeared. The more medial became sparser and shifted to the depth o f the lunate sulcus and finally disappeared. Finally, in the most ventral sections, an area of sparse degeneration appeared in the posterior bank of the superior temporal sulcus. In m o n k e y KS 24 a lesion was made above the line o f representation of the horizontal meridian in the striate cortex. The lesion was parallel to this line but was more posterior than the lesion in m o n k e y KS 21. Otherwise, it was of the same dimensions as in m o n k e y KS 21. The degeneration in this m o n k e y was restricted almost completely to the posterior b a n k of the lunate sulcus. In the most dorsal sections to reveal degenerated fibers, there were 2 areas of degeneration, the more lateral one being sparse and the more medial one moderate. The lateral area of degeneration occupied a small region beyond the convexity o f the opercular lip** whereas the more medial one was restricted to the medial one-quarter of the posterior b a n k o f the lunate sulcus. Further ventrally, the 2 areas of degeneration coalesced to yield a single area of degeneration. Within this single area, the more medial two-thirds was some 3 times as heavy as the more lateral third. In addition, the degeneration extended medially so that it n o w came to occupy the depth of the lunate sulcus as well. This picture of degeneration was maintained until the n u m b e r o f degenerated fibers decreased and finally disappeared. N o sharp separation between the 2 areas of degeneration at more ventral levels was observed in this monkey, as it was in m o n k e y K S 23. In m o n k e y K S 20 a lesion was attempted in the foveal representation area. The lesion did not involve white matter. The degeneration observed was fairly widespread, * It is almost impossible to section the brain in the zero horizontal plane and sometimes either the anteroposterior tilt or the mediolateral tilt may be exaggerated, as happened in this brain. A sideward bank is indicated by the thickness of the cortex and the configuration of the sulci and gyri, the cortex becoming thicker the greater the tilt. A lunate sulcus that is still deep when the inferior occipital sulcus appears indicates a lateral tilt. Conversely, a very shallow lunate sulcus in the absence of the inferior occipital sulcus would indicate a medial tilt. The tilt in this animal was lateral. ** In this animal there was some lateral tilt. Brain Research, 14 (1969) 271-291 280 s.M. ZEKI including the anterior bank of the lunate sulcus, the prelunate gyrus and the posterior bank of the superior temporal sulcus. In the most dorsal sections, sparse degeneration was limited to the lateral one-third of the anterior bank of the lunate sulcus. As the degeneration was traced ventrally through the horizontal sections, this area of degeneration spread laterally to occupy the posterior half of the prelunate gyrus. At more ventral levels, sparse degeneration appeared at the opercular lip which became increasingly heavy as the region of coexistence of the lunate and inferior occipital sulci was approached. At this level, the degeneration in the prelunate gyrus spread anteriorly to cover the whole of the prelunate gyrus, on the surface. Thus, traced from posterior to anterior, there was heavy degeneration along the opercular lip. The shallow posterior bank of the lunate sulcus was free of degeneration whereas the whole of the anterior bank contained moderate degeneration. The surface of the prelunate gyrus, which at this level is split by the shallow inferior occipital sulcus, was covered throughout by moderate numbers of degenerated fibers although the shallow inferior occipital sulcus itself was entirely free of degenerated fibers. At still more ventral levels, there was a moderate amount of degenerated fibers on the posterior side of the inferior occipital gyrus which, becoming sparser, finally disappeared. Comparison of this picture of degeneration following a lesion in the 'cortical fovea' with the degeneration ensuing following sectioning of the splenium will reveal the remarkable similarity between the two pictures (see Fig. 7). The areas of degeneration outlined here, following a lesion in the area of representation of the fovea in the striate cortex, are also in agreement with the work of Myers 13, the only difference being that in Myers' scheme the degeneration in the inferior occipital gyrus is more widespread ventrally. This minor discrepancy might be due to the more restricted extent of the lesion in this animal. Summary of results of degeneration following striate cortex lesions The degeneration in monkeys with lesions in striate cortex shows quite definitely that the upper and lower field representations in striate cortex project topographically to prestriate cortex. Any lesion made above the line of representation of the horizontal meridian in the striate cortex leads to degeneration in dorsolateral prestriate cortex and any lesion made below the horizontal line in striate cortex leads to degeneration in ventrolateral prestriate cortex only. This is essentially in agreement with the results of Myers 13 and Cowey 5. The results of monkey KS 12 show that there are 3 re-representations of the vertical meridian in prestriate cortex: at the opercular lip, in the lunate sulcus and in the posterior bank of the superior temporal sulcus. Further, the results of monkeys KS 21, 22, 23, 24, 25 and 16 show that areas more medial to the opercular lip in either the lunate sulcus or the inferior occipital sulcus lead towards the representation of the horizontal meridian and that areas still more medially situated in these 2 sulci lead towards the representation of the vertical meridian once more. The foveal representation in prestriate cortex appears to claim a large space and involves considerable parts of the prelunate gyrus as well as the anterior bank of the lunate sulcus and the posterior bank of the superior temporal sulcus. Brain Research, 14 (1969) 271-291 CENTRAL VISUAL FIELDS IN PRESTRIATE CORTEX 281 C. Interhemispheric connections The appearance of the representation of the vertical meridian in the depth of the lunate sulcus and the posterior bank of the superior temporal sulcus made it necessary to study the interhemispheric connections of these areas. Myers ~, in his schema, considered the cortex in the lunate sulcus (his 'striate receptive 19') to be free of connections with the opposite hemisphere. Such a lack of connections between representations of the vertical meridian would be contrary to the generally accepted finding that the midline is interhemispherically connected ~s,17. It was decided, therefore, to repeat this work and, in addition, to make lesions in prestriate cortex to determine whether the connections are homotypical or heterotypical or both. The splenium was sectioned in monkey KS 15 in which survival was 9 days. Gross examination of the post-mortem brain as well as serial Nissl-stained sections revealed that the splenium had been completely sectioned to a distance of about 9 mm from the tip of the splenium. As this paper is concerned with lateral prestriate cortex, the degeneration observed in medial cortical areas will not be reported. The picture of degeneration that emerged is a combination of the degeneration seen following a lesion in the representation of the vertical meridian of striate cortex (KS 12) and the foveal area of representation in striate cortex (KS 20) (see Fig. 7). Thus, in dorsal sections, the opercular lip received a moderate number of degenerated fibers and this degeneration, which involved all cortical layers, extended just beyond the opercular lip. The posterior bank of the lunate sulcus was free of degenerated fibers throughout. Degeneration commenced again in the medial one-quarter of the anterior bank of the lunate sulcus. This was followed by an area free of degenerated fibers which in turn was followed by another area of degeneration occupying the lateral one-quarter of the anterior bank of the lunate sulcus. The surface of the prelunate gyrus was free of degeneration and degenerated fibers were seen again in the middle of the posterior bank of the superior temporal sulcus. $ 15 A. i Fig. 7. Areas of degeneration following section of the splenium. Brain Research, 14 (1969) 271-291 282 s.M. ZEKI Traced in more ventral sections, this picture of the degenerated areas was maintained with the degeneration in the lateral quarter of the lunate sulcus spreading laterally to cover increasing proportions of the prelunate gyrus. At the level of coexistence of the inferior occipital and lunate sulci (Fig. 7, section B), the degeneration along the opercular lip was maintained and the posterior bank of the shallow lunate sulcus was free of degenerated fibers. In the anterior bank, the 2 fields of degeneration united to form a single uninterrupted field of moderate degeneration covering the area from the depth of the lunate sulcus to the lip of the shallow inferior occipital sulcus. The inferior occipital sulcus was free of degenerated fibers but the inferior occipital gyrus received a moderate number of degenerated fibers. Finally, the moderate degeneration at the posterior bank of the superior temporal sulcus was maintained. At still more ventral levels, once the lunate sulcus had been totally replaced by the inferior occipital sulcus (Fig. 7, section C) the picture was very similar to that observed at more dorsal levels. Thus, the opercutar lip received a moderate number of degenerated fibers. The posterior bank of the inferior occipital sulcus was totally free of degenerated fibers and degenerated fibers appeared again roughly in the middle of the anterior bank of the sulcus. The posterior surface of the inferior occipital gyrus contained degenerated fibers in moderate numbers and the moderate degeneration in the posterior bank of the superior temporal sulcus was maintained. It is not necessary to describe the remaining 5 monkeys with lesions in prestriate cortex in great detail as the degeneration seen following these lesions are components of the degeneration seen following section of the splenium. In monkey K S 6 it was attempted to remove the whole of the posterior surface of the lunate sulcus. There was involvement of white matter and, in addition, of cortex over the lateral quarter of the anterior bank of the lunate sulcus. The posterior part of the prelunate gyrus was also involved in its upper parts. Survival was 11 days. In the contralateral hemisphere, the degeneration was limited to the opercular lip and the lateral quarter of the anterior bank of the lunate sulcus. In monkey K S 7 a small lesion was made in the opercular lip; the lesion involved white matter. Survival was 4 days. In the contralateral hemisphere the degeneration was limited to the opercular lip. In monkey K S 10 a lesion was made in the opercular lip, the lesion being much more extensive than that in monkey KS 7. The lesion thus involved the whole dorsolateral surface up to the region of coexistence of the lunate and inferior occipital sulci. There was some involvement of white matter. Survival was 11 days. The degeneration in the contralateral hemisphere was limited to the opercular lip, the degenerated fibers distributing in all cortical layers. Where the lunate and inferior occipital sulci meet, the degeneration became somewhat more widespread, occupying the whole of the narrow prelunate gyrus at this level. In monkey K S 8 a small lesion was made in the posterior part of the prelunate gyrus. Survival was 10 days. Degeneration in the contralateral hemisphere was limited to the lateral quarter of the anterior bank of the lunate sulcus. Finally, in monkey KS 11 a lesion was made in the anterior part of the prelunate gyrus, the lesion occupying the whole dorsolateral surface up to the region of coBrain Research, 14 (1969) 271-291 CENTRAL VISUAL FIELDS IN PRESTRIATE CORTEX 19 I 18 ~1 ,~ o~ el •---.. 44":, ~'::?:~" / I ~ ~/ 283 LOWER VISUAL FIELD OF Vl - UPPER VISUAL ~. FIELD OF Vl zl / 19 ~' 18 Fig. 8. A schematic representation of prestriate cortex and its relation to striate cortex. The prestriate cortex may be completely divided into upper and lower sections, corresponding to the upper and lower retinal quadrants, by an imaginary prolongation of the line of representation of the horizontal meridian in the primary visual cortex (VI). The horizontal meridian re-representation in prestriate cortex is almost at right angles to its representation in striate cortex. This horizontal re-representation may be considered to form the boundary between the areas 18 and 19, as defined in this study. Note that the areas 18 and 19 of this study are within the lunate sulcus. Points further away dorsally (or ventrally) along the vertical meridian in striate cortex project to more dorsal (or ventral) points in prestriate cortex. Points further peripheral along the horizontal line project to more dorsal or ventral levels, depending on whether they are in the upper or lower visual field representation, the projection from the 2 fields of representation being separate. Not shown is the third vertical and horizontal rerepresentation in the superior temporal sulcus. existence of the inferior occipital a n d l u n a t e sulci. Survival was 14 days. I n the contralateral hemisphere, there was no degeneration except at more ventral levels (corres p o n d i n g to the region where the lunate sulcus becomes shallow a n d the inferior occipital sulcus begins). A t this level, the anterior part of the prelunate gyrus was recipient of a sparse n u m b e r of degenerated fibers. DISCUSSION The results given above enable us to present a schema of the r e p r e s e n t a t i o n of central visual fields in prestriate cortex (see Fig. 8). The lateral prestriate cortex m a y be divided completely, by a n i m a g i n a r y line f o r m i n g a n extension of the horizontal m e r i d i a n representation in striate cortex, into upper and lower prestriate cortex related to the lower and upper visual field representation in striate cortex. A n y area above the h o r i z o n t a l m e r i d i a n line in striate cortex projects to upper prestriate cortex only a n d any area below the h o r i z o n t a l line in striate cortex projects to lower prestriate cortex only. This is in agreement with the results of Cowey 5 a n d Myers la. W i t h i n the u p p e r a n d lower prestriate areas, we may distinguish further two areas in which the r e p r e s e n t a t i o n is f r o m vertical to h o r i z o n t a l a n d from h o r i z o n t a l to vertical. We m a y refer to these two areas, provisionally, a s areas 18 and 19 (see section o n N o m e n c l a t u r e ) . The area 18 of prestriate cortex related to the lower visual Brain Research, 14 (1969) 271-291 284 s.M. ZEKI Fig. 9. Diagrammatic representation of the areas 18 and 19 as defined in this study, as these would appear in a horizontal cross-section passing through the lunate (or inferior occipital) sulcus. For further information see text. v, Area of representation of vertical meridian; h, Area of representation of horizontal meridian. field representation in striate cortex extends from the striate prestriate boundary to the middle of the posterior bank of the lunate sulcus, at which point the horizontal meridian is represented. The region between the middle of the posterior bank of the lunate sulcus to the medial one quarter of the anterior bank of the lunate sulcus may be said to be occupied by the area 19. In lower prestriate cortex, related to upper visual field representation in striate cortex, the same relations apply, with the inferior occipital sulcus substituting for the lunate sulcus. Areas 18 and 19 of prestriate cortex related to the lower and upper visual fields form triangles with the upper and lower 18 and 19 meeting each other at their apices. The apex itself is occupied by the area of foveal representation which extends outwards (anteriorly) beyond the confines of 18 and 19, as defined above. The manner in which the vertical meridian representation at the striate prestriate boundary and that at the lunate sulcus come closer together at more ventral levels to give this pyramidal appearance can be seen in monkey KS 12. In this animal, which had a lesion just behind the representation of the vertical meridian in striate cortex, the two areas of degeneration at the opercular lip and the medial quarter of the anterior bank of the lunate sulcus came increasingly closer together at more ventral levels. This may be in part a secondary phenomenon due to the manner in which the lunate sulcus becomes shallower at more ventral levels. There appears, however, to be a real shift in position of the vertical meridian representation at the lunate sulcus. For in monkey KS 12, the area of degeneration shifted from the medial quarter of the anterior bank of the lunate sulcus to the depth of the sulcus. Other evidence for the approximation of the two representations of the vertical meridian comes from monkey KS 15 in which the splenium was sectioned. In this animal, the two vertical meridian representations, which were separated by the whole of the posterior bank of the lunate sulcus, came much closer together at more ventral levels once this sulcus had become shallower. Brain Research, 14 (1969) 271-291 C E N T R A L V I S U A L FIELDS IN P R E S T R I A T E C O R T E X 285 The exact position of the representation of the vertical meridian in the lunate sulcus cannot be determined in an anatomical study of this nature. But the position of the lesion in monkey KS 12 and the ensuing degeneration, in the lunate sulcus, together with the degeneration in monkey KS 15 following section of the splenium, would indicate that the vertical meridian in the prestriate cortex related to the lower visual field is represented in the medial quarter of the anterior bank of the lunate sulcus. This is the position in which degeneration is observed following sectioning of the splenium and if it is accepted that only the midline (i.e., the areas of representation of the vertical meridian and the foveal area) projects interhemispherically 17, then it would be reasonable to suppose that this is the position of the representation of the vertical meridianin the lunate sulcus. That the degeneration in monkey KS 12 occupied a somewhat more medial position may be explained by the fact that the lesion in this animal was not at the representation of the vertical meridian in striate cortex but just posterior to it. Although no lesion was made in the area of representation of the vertical meridian in the inferior striate cortex (upper visual field), by the same reasoning applied above the representation of the vertical meridian in prestriate cortex related to the upper visual field falls in the medial one third of the anterior bank of the inferior occipital sulcus. This is the region in which degeneration is observed following sectioning of the splenium. As in prestriate cortex related to lower visual field, the two representations of the vertical meridian in prestriate cortex related to the upper visual field come closer together from ventral to dorsal. The evidence for this is the manner in which the two areas of degeneration at the opercular lip and in the inferior occipital sulcus come closer together at more dorsal levels, where the inferior occipital sulcus begins and is therefore shallow. The re-representation of the horizontal meridian in prestriate cortex is more complex. The degeneration in monkey KS 21 was restricted to the posterior bank of the lunate sulcus. In this animal the lesion fell just above the horizontal line in striate cortex. In monkey KS 22, the degeneration was restricted to the posterior bank of the inferior occipital sulcus and the lesion in this animal was just below the horizontal line in striate cortex. The conclusion therefore is that the horizontal meridian is represented in prestriate cortex related to the upper visual field and independently in prestriate cortex related to lower visual field. This is not to say that each point on the horizontal line in striate cortex projects to two points. Rather, any point above the zero horizontal line projects only to prestriate cortex related to the lower visual field and forms a re-representation of the horizontal meridian there. In the same manner, any point below the horizontal line in striate cortex projects to prestriate cortex related to upper visual field and forms a re-representation of the horizontal meridian there. If the horizontal re-representation in upper and lower prestriate cortex were joined together, then the line so formed would be almost at right angles to the horizontal line in striate cortex (see schema of Fig. 8). It is not clear from this study how the zero horizontal line is represented in prestriate cortex. It would be practically impossible to make a lesion restricted purely to the zero horizontal meridian in striate cortex in an anatomical study. Brain Research, 14 (1969) 271-291 286 S.M. ZZ~:I Fig. 10. Diagram to show the manner in which the distance between the vertical and horizontal meridian representations varies. This distance is much smaller between points a and b than between points a" and b'. Consequently, any lesion placed between a and b would remove much more of the visual field between the vertical and horizontal meridians than an equal sized lesion placed more peripherally. The dashed lines indicate the representation of the vertical and horizontal meridians in striate cortex. The foveal area appears to claim a large field of prestriate cortex. Thus, it extends into prelunate gyrus and the lateral one-third of the anterior bank of the lunate sulcus. In addition, it extends ventrally into the inferior occipital gyrus and appears to send a finger like projection for a short distance around the striate prestriate boundary (see degeneration in monkey KS 20). This projection of the cortical fovea is essentially in agreement with the projection obtained by Myers 13. The somewhat more restricted projection in this study may be due to the more restricted extent of the lesion in the 'cortical fovea' in KS 20. If it is accepted, however, that the midline projects interhemispherically, then the results of monkey KS 20, together with the results of monkey KS 15, would confirm the results of Myers 13 where the foveal representation in prestriate cortex is concerned. It is of some interest to enquire why the lesions in monkeys KS 21, KS 22 and KS 25, which were so close to the line of representation of the horizontal meridian in striate cortex, led to such widespread degeneration in the posterior banks of the lunate and inferior occipital sulci. In the first place, if there are indeed two areas (18 and 19) which have a common boundary at which the horizontal meridian is represented (that is to say the two areas each have an independent representation of the horizontal meridian at the boundary at which they meet), then it would seem reasonable to assume that a lesion near the horizontal line in striate cortex would lead to degeneration in two areas which, because of their proximity, cannot be easily told apart. Fig. 10 may supply another partial answer. All the visual field from vertical to horizontal is represented between any two points at the vertical and horizontal lines of striate cortex. Since these lines come closer together, the whole of the visual field representation between the vertical and horizontal meridian representations is restricted to a smaller cortical space in central regions as compared to more peripheral regions. Thus, a lesion made more anteriorly Brain Research, 14 (1969) 271-291 CENTRAL VISUAL FIELDS IN PRESTRIATE CORTEX 287 would be expected to remove more of the visual field representation between the vertical and horizontal meridians than a similar sized lesion placed more peripherally. Since the vertical and horizontal meridians are topographically represented in prestriate cortex, it would a priori be expected that a lesion placed in striate cortex between the representation of the vertical and horizontal meridians would lead to two distinct regions of degeneration in prestriate cortex, one in area 18 and one in area 19. Actually, this result is obtained but not with great clarity. In monkey KS 16, two regions were evident. In monkey KS 23 two regions were also evident but they coalesced to form a single field of degeneration which occupied the posterior bank of the lunate sulcus. The distinguishing feature in this animal was the heavier degeneration in the more medial part of this field of degeneration as compared to the more lateral part. In monkey KS 24 in which a lesion was made parallel to the representation of the horizontal meridian but more dorsal and posterior than in monkey KS 21, there was a single field of degeneration in the posterior bank of the lunate sulcus which extended to the depth of the sulcus. In this field of degeneration, yet again, the only distinguishing feature was the markedly heavier degeneration on the more medial side compared to the more lateral side. As the lesion in this animal was placed between the vertical and horizontal meridian representation (though nearer to the horizontal), two distinct areas of degeneration were expected. Instead, only one appeared though the expected shift to the bottom of the lunate sulcus also appeared. The most probable conclusion is that a single area in striate cortex does not project in a strictly topographical sense to the areas 18 and 19 as here defined. Rather, any point in striate cortex projects to multiple points in areas 18 and 19 and only a gross form of topography is maintained. This supports the results of Cowey 5 who, using punctiform stimulation of the retina in the squirrel monkey, found that prestriate receptive fields were larger than those in the striate area and that adjacent points in the striate area projected to overlapping areas in prestriate cortex. Of the projections into the two distinct areas, the results of monkey KS 22, KS 23, KS 24 and KS 25 would suggest that the area 19 receives a heavier projection than the area 18. It is not clear if and how the horizontal meridian is represented between the two representations of the vertical meridian at the anterior bank of the lunate sulcus and at the posterior bank of the superior temporal sulcus (and also in the anterior bank of the inferior occipital sulcus and the posterior bank of the superior temporal sulcus in its lower parts). It was thought that because so much of the area between the two representations of the vertical meridian is occupied by 'foveal cortex', the representation of the horizontal meridian may have been displaced more dorsally or ventrally. However, no degeneration at more dorsal levels, or even medially, appeared in monkeys KS 21 and KS 24 and no degeneration at more ventral levels appeared in monkeys KS 22 and KS 25. It is possible that there is no such re-representation or that it is the horizontal meridian representation at the posterior bank of the lunate and inferior occipital sulci that project to form a horizontal re-representation at the prelunate and inferior occipital gyri. As no lesions were made in the posterior banks of these two sulci in this study, no definite conclusions can be drawn. In the one animal with a lesion extending from peripheral to vertical in striate cortex (KS 16) in which Brain Research, 14 (1969) 271-291 Fig. 11. Photomicrographs of the degeneration seen in this study. A shows the sparse degeneration seen in the depth of the lunate sulcus in monkey KS 12. B shows the moderate degeneration observed in the lateral part of the area of degeneration in the posterior bank of the lunate sulcus in monkey KS 23. C is taken from the same section but from the medial side of the area of degeneration in the posterior bank of the lunate sulcus. The degeneration here is heavy ( × 250). D is a photograph of a section taken from the lateral one-third of the anterior bank of the lunate sulcus in monkey KS 16, showing the vertically oriented fibers which are prominent in this region ( x 400). Inset C, Degenerated fiber as it appears at a magnification of 1000. C E N T R A L VISUAL FIELDS IN P R E S T R I A T E C O R T E X 289 there was degeneration in the prelunate gyrus, there was a slight involvement of the striate prestriate boundary. Myers 13 has claimed that this boundary (his 'juxta-striate 18') projects to the prelunate gyrus (his 'area 18 proper') and this would be confirmation of his finding. If this is so, however, it must be assumed that this projection is devoid of all except the grossest topography since the degeneration in the prelunate gyrus of monkey KS 16 was very widespread whereas the actual involvement at the striate prestriate boundary was very small. Finally, it is not very clear from this study how the projections to the posterior bank of the superior temporal sulcus from the striate cortex are organized. All the lesions approaching the vertical line of representation in striate cortex led to degeneration here even when no degeneration was found at the vertical meridian representation at the striate prestriate boundary, the anterior bank of the lunate sulcus or the anterior bank of the inferior occipital sulcus. For example, in monkey KS 21, no degeneration was apparent at the striate prestriate boundary or in the anterior bank of the lunate sulcus; yet there was a moderate amount of degeneration in the posterior bank of the superior temporal sulcus. Exactly the same picture appeared in monkey KS 22 in which the lesion was just below the line of representation of the horizontal meridian in striate cortex. On the other hand, in both monkeys KS 24 and KS 25, in which the lesions were more peripherally situated, some degeneration also appeared in the posterior bank of the superior temporal sulcus. In monkey KS 24 this was sparse but well localized while in monkey KS 25 it was very sparse and appeared on the surface of the inferior occipital gyrus (the superior temporal sulcus becomes very anterior at the level at which degeneration was seen in this monkey). Degeneration in the posterior bank of the superior temporal sulcus was also observed in monkey KS 12 with a lesion just behind the representation of the vertical meridian in striate cortex and in monkey KS 20 with a lesion in the 'cortical fovea'. No readily visible organization can be determined from this study and it remains for future work to determine if such an organized projection to this cortical field does exist. It is of some interest to compare the subdivisions proposed here with those given by Myers 13. Essentially, Myers 13 subdivides his prestriate cortex into four areas depending upon whether they receive fibers from striate cortex and from opposite prestriate cortex. His 'juxta striate 18' which is the striate prestriate boundary is recipient of fibers from the striate cortex and also projects to the opposite 'juxtastriate 18'. In his scheme, the lunate sulcus is recipient of fibers from the striate cortex only, there being no connections with the opposite hemisphere. This area forms his 'striate receptive 19'. This is followed on the surface of the prelunate gyrus by two areas: 'area 18 proper', in connection with the opposite hemisphere and situated in the posterior part of the prelunate gyrus and the 'area 19 proper' which has no connections with the opposite hemisphere and is situated in the anterior part of the prelunate gyrus. The present scheme differs from that proposed by Myers 13 in that the lunate sulcus (as well as the inferior occipital sulcus) is subdivided into (at least) two areas in which there is an orderly representation from vertical to horizontal and then back to vertical. The areas in which the midline is represented are connected with the ,Brain Research, 14 (1969) 271-291 290 s.M. ZEKI opposite hemisphere. Thus, the distinction between the two areas in the present study is made on the manner in which the visual fields are re-represented in prestriate cortex and not on the basis of whether they are in connection with the opposite hemisphere only or with striate cortex only. NOMENCLATURE The prestriate cortex of monkey has been studied by several different methods which have yielded different results. Is is not surprising, therefore, that different authors have used different terminologies to name these areas. It is suggested that the terms 'secondary visual cortex', 'visuo-psychic band' and 'peri- and parastriate cortex' be dropped altogether as somewhat meaningless terms. These terms have found fairly wide usage but unfortunately do not refer to any specific area defined by a specific technique and with specific limits. The terms 'fields OA and OB' should be used when reference is made to the subdivisions proposed by Von Bonin and Bailey 1, based on the cytoarchitectonic method, and should not be used as synonyms for the areas ' 18 and 19' of Brodmann 3. The term 'prestriate cortex' may be used as a general term to indicate the cortical field receiving fibers from the striate cortex, without indication of the subdivisions that exist in this cortical field. The terms 'areas 18 and 19' deserve special comment. These are the numbers used by Brodmann 3 to designate the two areas surrounding the striate cortex in the monkey. Since then, 'areas 18 and 19' have found usage in the cat so that 18 and 19 in this species 14 have become the anatomical terms for the physiologically defined V2 and V3 (ref. 8). If future physiological studies show two areas with different physiological properties corresponding to the two areas defined here as 18 and 19, then these terms may be used as the anatomical terms for the physiological V2 and V3 in monkey. When reference is made to the 18 and 19 of Brodmann 3, then, this should be clearly stated, as the provisional 18 and 19 proposed here are different to the 18 and 19 of Brodmann 3. SUMMARY 1. The organization of the monkey prestriate cortex has been studied by tracing degenerated fibers following discrete lesions in the striate cortex. 2. It has been shown that there are at least two distinct areas in prestriate cortex. Proceeding rostrally from the striate prestriate boundary there is first an area 18 in which there is a sequential representation of visual field from vertical meridian to horizontal meridian and an area 19 in which the representation is from horizontal to vertical. 3. These two areas may be further subdivided into two subareas each, corresponding to the re-representation of the upper and lower visual fields. 4. The fovea has been found to be represented in prestriate cortex at a position at which the areas 18 and 19 of the upper and lower visual fields juxtapose. 5. The areas proposed in this paper are discussed in relation to previous subdivisions of prestriate cortex. Brain Research, 14 (1969) 271-291 CENTRAL VISUAL FIELDS IN PRESTRIATE CORTEX 291 ACKNOWLEDGEMENTS I w i s h to t h a n k D r . J a c k H. M e n d e l s o n a n d D r . N a n c y K. M e l l o for the facilities t h e y m a d e a v a i l a b l e a n d for t h e i r e n c o u r a g e m e n t d u r i n g the c o u r s e o f this w o r k . T h i s r e s e a r c h was c o n d u c t e d at the N a t i o n a l C e n t e r for P r e v e n t i o n a n d C o n t r o l o f A l c o h o l i s m , N a t i o n a l I n s t i t u t e o f M e n t a l H e a l t h , d u r i n g the a u t h o r s t e n u r e as a V i s i t i n g A s s o c i a t e . T h i s p r o j e c t was s u p p o r t e d by the N a t i o n a l I n s t i t u t e o f M e n t a l Health. REFERENCES 1 BONIN, G. VON, AND BAILEY, P., The Neocortex of Macaca Mulatta, Univ. of Illinois Press, Urbana, 1947. 2 BONIN, G. VON, GAROL, H. W., AND MCCULLOCH, W. S., The functional organization of the occipital lobe, Biol. Syrup., 7 (1942) 165-192. 3 BRODMANN, K.., Beitrfige zur histologischen Lokalisation der Grosshirnrinde, J. Psychol. Neurol. (Lpz.), 4 (1905) 176-226. 4 CLARK, W. E. LEG., Observations on the association fibre system of the visual cortex and the central representation of the retina, J. Anat. (Lond.), 75 (1941) 225-235. 5 COWEY, A., Projection of the retina on to striate and prestriate cortex in the sqnirrel monkey, Saimiri sciureus, J. Neurophysiol., 27 (1964) 366-396. 6 DANIEL, P. M., AND WHITTERIDGE, O., The representation of the visual field on the cerebral cortex in monkeys, J. Physiol. (Lond.), 159 (1961) 203-221. 7 FINK, R. P., AND HEIMER, L., Two methods for selective silver impregnation of degenerating axons and their synaptic endings in the central nervous system, Brain Research, 4 (1967) 369-374. 8 HUBEL,D. H., AND WIESEL, T. N., Receptive fields and functional organization in two non-striate visual areas (18 and 19) of the cat, J. Neurophysiol., 28 (1965) 229-289. 9 K_UYPERS, H. G. J. M., SZWARCBART, M. K., MISHKIN, M., AND ROSVOLD, H. E., Occipitotemporal cortico-cortical connections in the rhesus monkey, Exp. Neurol., 11 (1965) 245-262. 10 LASHLEY,K. S., AND CLARK, G., The cytoarchitecture of the cerebral cortex of Ateles: a critical examination of architectonic studies, J. comp. Neurol., 85 (1946) 223-305. 11 METTLER,F., Corticofugal fiber connections of the cortex of Macaca mulatta. The occipital region, J. comp. Neurol., 61 (1935) 221-256. 12 MYERS, R. E., Commissural connections between occipital lobes of the monkey, J. comp. NeuroL, 118 (1962) 1-16. 13 MYERS,R. E., Organization of visual pathways. In E. G. E'rTLINGER(Ed.), Functions of the Corpus Callosum, Churchill, London, 1965, p. 133. 14 OTSUKA, R., UND HASSLER, R., Ober Aufbau und Gliederung der corticalen Sehsph/ire bei der Katze, Arch. P~Tchiat. Nervenkr., 203 (1962) 212 234. 15 PANDYA,D. P., AND VtGNOLO, L. A., Interhemispheric neocortical projections of somatosensory areas 1 and lI in the rhesus monkey, Brain Research, 7 (1968) 300-303. 16 TALBOT, S. A., Arid MARSHALL, W. H., Physiological studies on neural mechanisms of visual localization and discrimination, Amer. J. Ophthal., 24 (1941) 1255-1264. 17 WI~ITTERIDCE,D., Area 18 and the vertical meridian of vision. In E. G. ETTLIN~ER(Ed.), Functions of the Corpus Callosum, Churchill, London, 1965, p. 115. 18 WILSON, M. E., AND CRAGG, B. G., Projections from the lateral geniculate nucleus in the cat and the monkey, J. Anat. (Lond.), 101 (1967) 677-692. 19 ZEKI, S. M., The secondary visual cortex of the monkey, Brain Research, 13 (1969) 197-226. Brain Research, 14 (1969) 271-291