Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Haemodynamic response wikipedia , lookup
Embodied language processing wikipedia , lookup
Perception of infrasound wikipedia , lookup
Neurophilosophy wikipedia , lookup
Bird vocalization wikipedia , lookup
History of neuroimaging wikipedia , lookup
Sensory substitution wikipedia , lookup
Affective neuroscience wikipedia , lookup
Clinical neurochemistry wikipedia , lookup
Environmental enrichment wikipedia , lookup
Embodied cognitive science wikipedia , lookup
Premovement neuronal activity wikipedia , lookup
Holonomic brain theory wikipedia , lookup
Neurocomputational speech processing wikipedia , lookup
Emotional lateralization wikipedia , lookup
Development of the nervous system wikipedia , lookup
Optogenetics wikipedia , lookup
Neuropsychology wikipedia , lookup
Neurolinguistics wikipedia , lookup
Nervous system network models wikipedia , lookup
Cognitive neuroscience wikipedia , lookup
Sound localization wikipedia , lookup
Neuroanatomy wikipedia , lookup
Brain Rules wikipedia , lookup
Eyeblink conditioning wikipedia , lookup
Animal echolocation wikipedia , lookup
Music psychology wikipedia , lookup
Neuroesthetics wikipedia , lookup
Neuroanatomy of memory wikipedia , lookup
Synaptic gating wikipedia , lookup
Neuroplasticity wikipedia , lookup
Evoked potential wikipedia , lookup
Aging brain wikipedia , lookup
Neuropsychopharmacology wikipedia , lookup
Human brain wikipedia , lookup
Metastability in the brain wikipedia , lookup
Sensory cue wikipedia , lookup
Cortical cooling wikipedia , lookup
Neural correlates of consciousness wikipedia , lookup
Neuroeconomics wikipedia , lookup
Time perception wikipedia , lookup
Feature detection (nervous system) wikipedia , lookup
Inferior temporal gyrus wikipedia , lookup
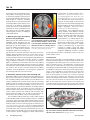
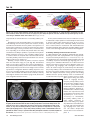
Tot 10 Auditory Brain Development in Children With Hearing Loss – Part One By Jace Wolfe, PhD, & Joanna Smith, MS Editor’s Note: This is the first installment of a two-part article. The conclusion will be published in the November issue. a. D b. Insula (short gyri) Percentral gyrus Central sulcus Postcentral gyrus r. Carol Flexer aptly puts it: It’s Planum temporale all about the brain. We hear with our brain. The ears are Wernicke’s just the way in. Early identifiarea cation of hearing loss and intervention Transverse must occur during the critical period of (Heschl’s) Gyrus language development in the brain. Listening happens in the brain, not in the ears. As pediatric hearing healthcare profesPrimary Auditory Cortex sionals, we are familiar with these mantras and catchphrases. In fact, we have heard and said these slogans so much that they Figure 1a. A visual representation of the classical ascending auditory pathway almost seem like clichés. But there are from the cochlea to primary auditory cortex. Figure 1b: Primary auditory cortex power truths behind clichés that make (Heschl’s gyrus) identified by the red arrow (Reproduced with permission: Bhatthem stand the test of time. Too often we nagar. Neuroscience for the Study of Communicative Disorders. 2nd ed. Wolters lose sight of the exact origins of clichés Kluwer Health, 2012). and buzz-phrases that underlie our lives. In this two-part article, we provide a Percentral a. Insula b. gyrus Central sulcus brief overview of several relevant research (short gyri) Postcentral gyrus studies examining the effects of hearing Planum loss and audiologic intervention on auditemporale tory brain development. This summary includes a survey of research investigating Wernicke’s area auditory brain development in humans and animals, with a focus on the work of Andrej Transverse (Heschl’s) Gyrus Kral, MD, PhD, one of the most prolific scholars in the area of early auditory brain development. His research on deaf white Figure 2a. Secondary auditory cortex identified by red arrows. Figure 2b. Seccats has substantially advanced our un- ondary auditory cortex identified by red arrows (Brodmann’s Areas 41 [primary] derstanding of the influence of deafness and 42 [secondary]; Bhatnagar, 2012). and cochlear implantation on auditory brain development. We would be remiss if we failed to ac10. The Auditory Brain knowledge a number of other brilliant researchers from around The auditory brain extends far beyond Heschl’s gyrus and is the world who have contributed to this line of study (including actually quite complex. In our auditory anatomy courses, we but not limited to Chris Ponton, Jos Eggermont, Anu Sharma, likely learned that auditory signals travel up the brainstem to the David Pisoni, David Ryugo, Bob Harrison, Karen Gordon, thalamus and the primary auditory cortex (otherwise known as Lynne Werner, Nina Kraus, and Patricia Kuhl). Heschl’s gyrus; Figs. 1a and 1b). Heschl’s gyrus resides within the Sylvian fissure and courses medially from the superior temporal gyrus. Tonotopic organization is preserved throughout Dr. Wolfe, left, is the director of this trip, including within the primary auditory cortex; additionaudiology at Hearts for Hearing ally, complex processing, which mediates functions—from basic and an adjunct assistant professor at the University of Oklasimple detection to complex localization and extraction of a homa Health Sciences Center signal of interest from competing noise—occurs in groups of and Salus University. Ms. Smith, neurons at all levels of the auditory nervous system. right, is a founder and the exFrom the primary auditory cortex, auditory signals travel to the ecutive director of Hearts for secondary auditory cortex, which has less defined boundaries Hearing in Oklahoma City. 14 The Hearing Journal October 2016 Tot 10 and components compared with the primary auditory cortex. Figure 2a shows an elementary example of many areas in temporal lobe and beyond that are typically thought to comprise secondary auditory cortex. It is well known that the secondary auditory cortex plays a prominent role in our ability to understand speech. For instance, in 1874, Wernicke noted that an insult to Brodmann area 22, located in the secondary auditory cortex (Fig. 2b), results in an inability to understand speech. representation or feature extraction (Allen. IEEE Trans. on Speech and Audio Processing 1994;2[4]:567; Kral. Brain Res Rev 2007;56[1]:259; Kral. e-Neuroforum 2015; 6:21). Research has shown that infants as young as 4 months begin to attend to the phonemes of his/her primary language while showing weaker responses to phonemes of foreign languages (Dahaene-Lambertz. Trends Neurosci 2006;29[7]:367). Researchers have suggested that the primary auditory cortex detects acoustic features that an individual deems important, and the secondary and higher-order areas 9. When all goes right, the auditory combine these features into meaningful Figure 3. PET scan image illustrating brain’s areas glow bright! representations (i.e., auditory objects; Kral. When a person has sufficient access to a broad bilateral activation of primary Neuroscience 2013;247:117; Kral, 2007). intelligible speech throughout the first and secondary auditory cortices while Higher-order auditory areas contain pluripfew years of life, the auditory areas of the a post-lingually deafened adult with a otent neurons that respond to multiple brain light up like Time Square in re- left ear CI listens to running speech modes of stimulation (e.g., a neuron that sponse to auditory stimulation (Fig. 3). (Reproduced with permission: Green. responds to auditory, visual, and tactile Green et al. used positron emission Hear Res 2005;205[1-2]:184). stimulation), possibly enabling multi-modal tomography (PET) scan testing to imintegration. Additional research is needed age the areas of the brain that were responsive when postto fully understand the roles of pluripotent neurons in the seclingually deafened adults listened to speech while using a ondary auditory cortex. cochlear implant (CI; Hear Res 2005;205[1-2]:184). To clarify, the participants had normal hearing during childhood, lost 7. I like bacon! their hearing as adults, and received a CI after a variable We have yet to develop a full understanding of exactly how range of duration of deafness (1 to 48 years). As shown in and where auditory objects are represented in the brain. DeFigure 3, a broad area of activation was seen in the auditory riving higher-order meaning from the sound we hear is cerareas of the brain. Specifically, activity in response to auditory tainly a complex process. “Fundamentally, everything that stimulation was observed both in primary and secondary audicomes into our minds reduces to patterns of neural activity,” tory cortices. Also of note is that this broad area of auditory according to Kai-How Farh, MD, a clinical geneticist at Bosactivation occurred bilaterally, even though the participant ton Children’s Hospital. In other words, each cognitive expewas listening with a CI only on the left ear. rience is represented by a unique network of neurons that produces the reality we perceive. For instance, when we 8. Secondary auditory cortex is the launching pad. hear the word “yellow,” a certain set of neurons responds Secondary auditory cortex is like the launching pad of the auacross the brain to produce the experience we associate ditory area of the brain. The complex connections between with the word. Figure 5a illustrates a network of neurons that the secondary auditory cortex and the rest of the brain are not may move across the brain to represent the word “yellow.” entirely elucidated, but research shows that the secondary Most of the responsive neurons reside in the primary and auditory cortex has multiple connections to other areas of the secondary cortices, but there are also neurons that respond brain and back to the primary auditory cortex in the form of from the frontal and parietal lobes (and possibly even in the efferent tracts. The connections between secondary auditory cortex and other areas of the brain are often referred to as Superior longitudinal fasciculus intra-hemispheric connections. An example is the arcuate fasArcuate fasciculus ciculus, which connects the temporal and the frontal lobe (Fig. 4). Numerous others exist, as well as connections to areas inferior to the cerebrum, such as the hippocampus. The secondary auditory cortex also sends a robust number of efferent fibers back to primary auditory cortex (i.e., feedback projections). It has been proposed that efferent fibers from the Uncinate fasciculus secondary auditory cortex likely play a role in tuning the primary auditory cortex, to focus on primary signals of interest (David. Inferior longitudinal fasciculus Proc Natl Acad Sci USA 2012;109:2144). Typical real-world environments are fraught with a cacophony of speech and environmental noises. For successful communication to occur, the Figure 4. Illustration of intrahemispheric fiber tracts providauditory system must be able to focus on the acoustic elements ing connections between different areas within one hemiof the listener’s spoken language—an ability called feature sphere of the cerebrum (Bhatnagar, 2012). 16 The Hearing Journal October 2016 Tot 10 a. b. sensor motor area sensor motor area frontal eye field frontal eye field frontal lobe parietal lobe prefrontal area Broca’s Area (in left hemsphere) visual visual association temporal lobe auditory auditory association frontal lobe parietal lobe prefrontal area Broca’s Area (in left hemsphere) visual visual association temporal lobe auditory auditory association Figure 5. A cartoon representation neural responsiveness (i.e., a neural network or connectome) in response to a. the word “Yellow,” and b. the sound of bacon frying in a pan. Note, the yellow stars are intended to represent groups of neurons firing in different areas of the brain (Bhatnagar, 2012). occipital lobe or multimodal areas of secondary auditory cortex). Engagement of the frontal lobe allows us to extract higherorder meaning from the word “yellow.” For instance, we may conclude that we dislike the color yellow or that yellow is our favorite color. Furthermore, we may associate the color yellow with a traffic light, a canary, a favorite shirt, or a banana. Neurons responding in the frontal and parietal lobes also likely contribute to our ability to produce or speak the word “yellow.” Finally, pluripotent neurons in the secondary auditory cortex or neurons within the occipital lobe interconnected with the secondary auditory cortex also respond to allow us to form an image of yellow in our mind’s eye. Similarly, a unique network or pattern of neurons respond when we hear bacon frying in a pan (Fig. 5b). That distinct sizzle in a pan elicits responses from neurons throughout primary and secondary auditory cortices. Even without seeing the bacon frying in the pan, we can form an image of it in our mind’s eye because of integration between auditory-responsive neurons in the secondary auditory cortex and visually responsive neurons. We remember how bacon tastes and feels, and may even begin to salivate as we hear the frying sound, all because of the integration between neurons. In short, each sound that comes into our minds from our ears is reduced to a unique pattern of neural activity. For that sound to possess higher-order meaning and come to life, it has to travel from the primary to the secondary auditory cortex and form a neural network or connectome with multi-modal areas throughout the brain. Once again, the secondary auditory cortex serves as the launching pad for this interaction and integration. 6. Primary auditory cortex was born to hear. In 1999, Nishimura and colleagues published their groundbreaking research that employed PET scan testing of brain responses to a variety of stimuli (Nature 1999;397[6715]:116). The participants were pre-lingually deafened adults who had no auditory experience and used sign language prior to receiving a CI in adulthood. Nishimura et al. observed neural activity in the brain in response to three different stimuli: running speech, sign language, and meaningless hand movements. As shown in Figure 6, neural activity was observed in the primary auditory cortex contralateral to the implanted ear (but not in the primary auditory cortices of both hemispheres) when the subjects listened to running speech. This finding indicates that the primary auditory cortex is hard-wired for sound. Even when a person does not have access to intelligible speech during the critical Horizontal sections relative to the intercommissural plane: period of language develop10 mm below 4 mm above 8 mm above ment, the primary auditory cortex still responds to auditory stimuli, as seen in young adults initially introduced to sound via CI after a lifetime of deafness. However, extended duration of deafness likely reduces the responsiveness and reorganization of the primary auditory cortex (Kral. Trends Neurosci 2012;35[2]:111). Nonetheless, the primary auditory cortex remains largely responsive to sound. This finding may explain Figure 6. PET scan imaging results showing neural responses in the brain of a pre- why adults who never had access lingually deafened adult with CI after auditory deprivation since birth. Responses are to sound and communicated via observed in a. the occipital lobe (in blue) in response to meaningless hand movesign language their entire lives ments, b. only the secondary auditory cortex (in yellow) in response to a story told can still detect whisper-level with sign language, and c. only in the primary auditory cortex contralateral to the imsounds with use of a cochlear implanted ear when a story is spoken while the participant used the cochlear implant for plant. the left ear (Reproduced with permission: Nishimura. Nature 1999;397[6715]:116). 18 The Hearing Journal October 2016