Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Cooperative binding wikipedia , lookup
Protein domain wikipedia , lookup
Nuclear magnetic resonance spectroscopy of proteins wikipedia , lookup
Protein mass spectrometry wikipedia , lookup
Intrinsically disordered proteins wikipedia , lookup
Protein purification wikipedia , lookup
Protein–protein interaction wikipedia , lookup
Western blot wikipedia , lookup
List of types of proteins wikipedia , lookup
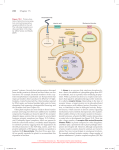
Receptor 接受器or受器 Intracellular receptor Intercellular signaling in animals by receptors 1 Two classes of receptors have two basic structural plans in cell membrane: 1. Trimeric G protein-linked receptors (GPCR) (e.g. glucagon-, serotonin- adrenalin-receptors) Ionotropic Metabotropic 2. Ion-channel receptors (ligand-gated ion-channels, e.g. the acetylcholine receptor) iontropic receptor G-protein coupled receptor Tyrosine kinase linked receptor Instrinsic enzymatic receptor 3. Receptors lacking intrinsic catalytic activity but directly associated with cytosolic protein tyrosine kinases 4. Receptors with intrinsic enzymatic activity (e.g. guanylate cyclase activity, protein phosphatase, serine/threonine kinase or tyrosine kinase activity) 5. Cell adhesion molecules 6. Intracellular receptors Four classes of cell-surface receptors 1. Ligand-gated ion channels (ionotropic) exist in cell membrane. e.g. nAChR 2. G-protein coupled receptors (metabotropic) exist in cell membrane e.g. mAChR 3. Tyrosine Kinase linked Receptor exist in cell membrane e.g. cytokine receptors 4. Instrinsic emzymatic receptor. Receptor Tyrosine kinase (RTK) can autophosphorylation. All receptors are proteins made up of varying numbers of subunits or transmembrane domains. Cell-surface receptors belong to four major classes GPCRs are involved in a range of signaling pathways, including light detection, odorant detection, and detection of certain hormones and neurotransmitters Many different mammalian cell-surface receptors including GPCRs are coupled to a trimeric signal-transducing G protein – made of an alpha, beta and gamma subunit complex Ligand binding activates the receptor, which activates the G protein, which activates an effector enzyme to generate an intracellular second messenger – e.g. adenylyl cyclase – converts ATP to cAMP depending on regulation at the effector enzyme – this pathway can be either activated or inhibited – by the type of G protein activated by the hormone-receptor complex – Gs proteins result in stimulation of the effector enzyme – Gi proteins result in inhibition of the effector enzyme adenylyl cyclase (AC) 2 NO-self enzymatic activity Four classes of cell-surface receptors -ligand binding changes the confirmation of the receptor so that specific ions flow through it the resultant ion movement alters the electric potential across the plasma membrane -found in high numbers on neuronal plasma membranes e.g. ligand-gated channels for sodium and potassium -also found on the plasma membrane of muscle cells -binding of acetylcholine results in ion movement and eventual contraction of muscle -lack intrinsic catalytic activity -binding of the ligand results in the formation of a receptor dimer (2 receptors) -this dimer than activates a class of protein called tyrosine kinases -this activation results in the phosphorylation of downstream targets by these tyrosine kinases (stick phosphate groups onto tyrosines within the target protein) -receptors for cytokines such as, interferons Itself enzymatic activity Signal transduction Cascade -also called receptor tyrosine kinases OR ligand-triggered protein kinases -similar to tyrosine-linked receptors - ligand binding results in formation of a dimer -BUT: they differ from tyrosine-linked receptors – intrinsic catalytic activity -means that ligand binding activates it and the activated receptor acts as a kinase -recognize soluble or membrane bound peptide/protein hormones that act as growth factors e.g. NGF, PDGF, insulin -binding of the ligand stimulates the receptor’s tyrosine kinase activity, -results in phosphorylation of multiple amino acid residues within its target such as serine and threonine residues -this phosphorylation activates downstream targets -its targets are generally other protein kinases –which phosphorylate their own downstream targets (other kinases??) 3 Itself enzymatic activity NO-self enzymatic activity Itself enzymatic activity Itself enzymatic activity Itself enzymatic activity NO-self enzymatic activity Cell adhesion molecular 4 Four classes of cell-surface receptors Nucleus or cytoplasmic receptor G-protein coupled receptors (GPCRs) Ligand binding activates a G-protein which in turn activates or inhibits an enzyme that generates a specific second messenger estrogen receptor 類酯醇X受體(Pregnane X receptor PXR How G proteins were discovered Gilman & Ross studying connection between adrenalin receptors and the enzyme adenylate cyclase which makes cyclic AMP ATP adenylate cyclase cAMP Expt 1 Expt 2 adrenalin adrenalin cyc- + wild-type extract (untreated) cyc- 可能野生種的 AC所導致 cAMP增加 no cAMP used a mutant cell line cyc- that bound adrenalin but appeared to lack adenylate cyclase lucky experiment that led to the Nobel prize!! cAMP Expt 2 adrenalin cyc- + wild-type extract (AC inactivated) 把野生種萃取物中AC抑 制掉,但還是有反應?顯示 是cyc- 還是有自己的AC 活性所致, 所以是以前 cyc-對的認知有錯誤,是 少某種物質而非AC cAMP !? Explanation: cyc- cells didn’t lack adenylate cyclase, but lacked another factor (G proteins) that activates adenylate cyclase Cyc- mutant ? Not without adenylate cyclase →without some thing 5 The importance of G-proteins The Nobel Prize in Physiology and Medicine 1994 "for their discovery of G-proteins and the role of these proteins in signal transduction in cells" G-protein linked receptors most common type of receptor when receptor is activated by stimulus operates via an intermediary – G protein (guanine nucleotide binding protein) G protein in turn regulates enzyme or ion channel all G-protein coupled receptors have 7 transmembrane spanning regions A major target for drug e.g. beta blockers, antihistamines Most act via hetero-trimeric G-proteins with cAMP, cGMP and PLC often being used as downstream effectors Receptors downregulated following ligand activation to assist with shutoff of the switch Alfred G. Gilman USA Martin Rodbell USA 1941- 1925-1998 Largest class of cell surface Receptors All have a structure with seven transmembrane alpha helical Loops Orientation is always conserved with N terminus outside, C terminus inside the cell and cytosolic segments interacting with G proteins Sequences at C3, C4 and sometimes C2 determine which G protein is activated Genome sequencing has revealed more new members Half of all known drugs bind to G protein-linked receptors Nearly 2500 GPCRs have been identified Bovine rhodopsin was cloned in 1983 (Nathans and Hogness); adrenergic receptor in 1986 (Dixon et al.) General structure: N-terminal segment: glycosylation, ligand binding (outer) C-terminal segment: phosphorylation and palmitoylation (inner) Seven transmembrane domain (TMs): form six loops (three exoloops and three-four cytoloops) and a TM core that could provide ligand specificity and regulatory mechanism Membrane bound effector proteins Effector proteins are then capable of amplifying the signals that will then be further transduced to secondary targets. Examples: – Adenylyl cyclase (cAMP synthesis) – Phospholipase Cβ (inositol lipid release from membrane) – Phospholipase A2 (arachidonic acid release from membrane, precursor of prostaglandin and leukotriene synthesis) – Guanylyl cyclase (cGMP synthesis) – Cyclic nucleotide phosphodiesterase (breakdown of cGMP and cAMP) – Potassium and calcium ion channels Gsα stimulates adenylyl cyclase GTP bound Gsα interacts with adenylyl cyclase The structural changes that are induced are not known, but the result is active enzyme Many signals, through different receptors, can activate Gsα, resulting in a higher concentration of GTPGsα and the production of higher levels of cAMP Forskolin, applied to cells, will activate pathways mediated by cAMP adenylyl cyclase activator 6 GPCR encoded for >1000 genes, represent app.1% of human gemone GPCRs -Among membrane-bound receptors, the G protein-coupled receptors (GPCRs) are the most diverse. -In vertebrates, this family contains 1000 – 2000 members (>1% of the genome). -GPCRs have been very successful during evolution, being capable of transducing messages as different as photons, organic odorants, necleotides, peptides, lipids and proteins. -GPCRs have a common ”central core”, composed of 7 transmembrane helical domains. -The fine-tuning of coupling of the receptor to G proteins is regulated by splicing, RNA editing and phosphorylation. Illustration of the central core of rhodopsin. The core is viewed from the cytoplasm. Characteristics of G proteins 1. G protein is an trimeric protein which binds guanine nucleotides. 2. They function to couple integral membrane receptors to target membrane-bound enzymes. 3. They can be considered molecular switches wherein… GDP (inactive) GTP (active) + 4. The dissociated subunit expresses GTPase activity. (A) GPCRs have a central common core made of seven transmembrane helices (TM-I to -VII) connected by three intracellular (i1, i2, i3) and three extracellular (e1, e2, e3) loops. The diversity of messages which activate those receptors is an illustration of their evolutionary success. (B) Illustration of the central core of rhodopsin. The core is viewed from the cytoplasm. cytoplasm The length and orientation of the TMs are deduced from the two-dimensional crystal of bovine and frog rhodopsin (Unger et al., 1997). The N- and C-terminal of i2 (including the DRY sequence) and i3 are included in TM-III and -VI. The core is represented under its 'active conformation'. The TM-VI and -VII lean out of the structure, the TM-VI turn by 30% on its axis (clockwise as viewed from the cytoplasm) (Bourne, 1997). This opens a cleft in the central core in which G proteins can find their way. i2 and i3 loops are the two main loops engaged in G protein recognition and activation. EMBO J. 18: 1723-1729 (1999) GPCRs 5. GTPS blocks GTPase activity of GTP. 7 Receptor activation… GPCRs activate different sub-classes of heterotrimeric G-proteins and effector systems (cont’d) Heterotrimeric G-proteins Highly conserved mechanism linking to most GPCR’s Following activation of GPCR the GDP on G( heterotrimeric Gprotein is switched for GTP causing dissociation to G and G after stimulation. Both can have signalling effects G Over 20 forms have now been identified in mammals. Divided into Gs which generally activate effector, Gi which generally inhibitory, Gq which generally act via phospholipases. The GTPase activity acts as a time dependent switch by converting GTP to GDP G About 6 forms of and 12 forms of have now been identified. The G stay as dimer and can regulate molecules including K+ channels and PI 3-kinases Nature Reviews Molecular Cell Biology 3; 639-650 Receptors cycle between resting and active states G proteins are activated in response to binding by an activated receptor General Themes in Heterotrimeric G protein Pathways May be > 20 GTP displaces GDP and subunits dissociate. Gα is activated Rockman H.A. et al (2002) Nature 415:206 8 The tools for G-protein receptor signal transduction research So, inhibitory signal ↓ So, stimulatory signal ↑ Basic structure/function of G-protein subunits: heterotrimers consist of one copy of alpha (39-45kDa), beta (35-36kDa) and gamma (5-7kDa) subunits G-protein ADP ribosylation + Nicotinamide CTX: for Gs PTX: for Gi Gs Bacterial toxin (PTX, CTX) ADP-ribose G GTPase 9 Family 1 contains most GPCRs including receptors for odorants (氣 味), small ligands, peptide hormone and glycoprotein. Disulphide bridge connects e1 and e2 and palmitoylated cysteine in C-terminal. Family 2 GPCRs have relative long Nterminal that contains several cysteines (network of disulphide bridge). Examples include glucagon, GnRH and PTH receptors. Family 3 GPCRs have very large span N-terminal sequences and C-terminal tail. The ligand binding domain is located in the N-treminus. The i3 loop is short and highly conserved. Representative samples are mGluR, Ca2+-sensing and GABA-B receptor. 85 kDa Functional regions within GPCRs: G-protein interacting domains & ligand binding domain The three subfamilies of 一種味覺 GPCRs are depicted with examples of their endogenous agonists. The binding modes of the orthosteric (直立) ligands for each receptor type are depicted by a green rectangle. The GPCR signals either by coupling to heterotrimeric Gproteins consisting of and subunits (which trigger a wide range of metabolic cascades and ion channel activities) or by direct association with effector molecules. AC, adenylyl cyclase; ATP, adenosine triphosphate; cAMP, cyclic adenosine monophosphate; PLC, phospholipase C; IP3, inositol-3,4,5-trisphosphate; DAG, diacylglycerol. Schematic presentation of the general structure of GPCRs and receptorreceptor-ligand interactions 1. Proteinase-activated receptor ( PAR ) Family PAR1~4 2. PAR2’s activation induces acute inflammation Platelet aggregation J Biol Chem 273:17299-17302,1998 10 Ligand Binding and GPCR Activation The G subunit of G protein cycles between active and inactive forms • Modes of ligand binding: Exclusive in TM core: photon, biogenic amines, nucleotides and lipid substrates (B) core, exoloops and N-terminal segment: peptides of ≤40 amino acids (C) N-terminal segment cleavage: protease( thrombin) (D) N-terminal segment and exoloops: glycoprotein hormone ( approximately 350 amino acids) (E) N-terminal segment (~600 amino acids): calcium channel, GABA and metabotropic receptor (F) FRET Fluorescence resonance energy transfer Applications for monitoring molecular interactions in Living cells by FRET Monitor protein-protein interaction Calcium sensor Monitor intramolecular conformational change Kinase activation sensor Receptor mediated activation of coupled G-proteins occurs within a few seconds of ligand in living cells M13, a peptide binds to calmodulin in calcium-dependent manner 11 Diversity (of physiological responses to GPCR stimulation) Receptor activation… GPCRs activate different sub-classes of heterotrimeric G-proteins and effector systems GRK: G-protein coupled receptor kinase GPCR ↓ Second messenger ↓ effector ↓ response TARGET TISSUE HORMONE MAJOR RESPONSE Thyroid gland thyroid-stimulating hormone (TSH) thyroid hormone synthesis and secretion Adrenal cortex adrenocorticotrophic hormone (ACTH) cortisol secretion Ovary luteinizing hormone (LH) progesterone secretion Muscle adrenaline glycogen breakdown Bone parathormone bone resorption Heart adrenaline increase in heart rate and force of contraction Liver glucagon glycogen breakdown Kidney vasopressin water resorption Fat adrenaline, ACTH, glucagon, TSH triglyceride breakdown TARGET TISSUE SIGNALING MOLECULE MAJOR RESPONSE Liver vasopressin glycogen breakdown Pancreas acetylcholine amylase secretion Smooth muscle acetylcholine contraction Blood platelets thrombin aggregation GPCR vs. RTK Complexity of GPCR signalling Cascades GPCRs cross talk with Receptor Tyrosine Kinases (RTK) Given such a diversity in responses, how does GPCR signaling specificity occur??? AC: adenylyl cyclase PDE: phosphodiesterase PLC: phospholipase C Multiple physiological responses 12 The mechanisms regulate (terminate) signaling form GPCR 1.GTP → GDP (exchange) 2. Degradation of second message, cAMP phosphodiesterase (cAMP →5’AMP) or cGMP phosphodiesterase… 3.Receptor phophorylation by down stream signal (cAMP → PKA → phosphorylation of receptor); feedback regulation (Desentization; heterologus or homologous) 4. Protein Phosphatase catalyzes removal by hydrolysis of phosphates that were attached to proteins via Protein Kinase A Turn off of the signal from GPCR Agonist or antagonist → receptor → activation of receptor specific enzyme (receptor kinase) → phosphorylation; directly from receptor action is called homologus desentization Anchoring proteins localize effects of cAMP to specific subcellular regions (new model for turn off signal from GPCR) Phosphodiesterase enzymes catalyze: cAMP + H2O AMP N N The phosphodiesterase that cleaves cAMP is activated by phosphorylation catalyzed by Protein Kinase A. N N H2 5' C O Thus cAMP stimulates its own degradation, leading to rapid turnoff of a cAMP signal. NH2 cAMP O 4' O H H 3' P O O- H 1' 2' H OH In heart muscle: adrenergic receptor → cAMP ↑→ activate PKA → C, catalytic region → phosphorylate → PDE → activation → degradation of cAMP PDE:phosphodiesterase, cAMP →5’AMP AKPA: ANCHORING PROTEIN 13 AKAPs (A-Kinase Anchoring Proteins) are scaffold proteins with multiple domains that bind to regulatory subunits of Protein Kinase A phosphorylated derivatives of phosphatidylinositol various other signal proteins, such as: • G-protein-coupled receptors (GPCRs) • Other kinases such as Protein Kinase C • Protein phosphatases • Phosphodiesterases AKAPs localize hormone-initiated signal cascades within a cell, and coordinate activation of protein kinases as well as rapid turn-off of such signals. Receptor desensitization occurs. This process varies with the hormone. Some receptors are phosphorylated via specific receptor kinases. The phosphorylated receptor may then bind to a protein arrestin, that promotes removal of the receptor from the membrane by clathrin-mediated endocytosis. First discovery: -adrenergic receptor Heterologous desensitization: – Four residues in the cytosolic domain of the adrenergic receptor can be phosphorylated by PKA – Activity of all Gs protein – coupled receptors, not just the -adrenergic receptor, is reduced Homologous desensitization: – Other residues in the cytosolic domain of the adrenergic receptor are phosphorylated by the receptorspecific -adrenergic receptor kinase (BARK) – BARK only phosphorylates the -adrenergic receptor which facilitates -arrestin binding to the phosphorylated receptor – Related with GRK (G-protein receptor kinase Hormonally induced negative regulation of receptors is referred to as homologousdesensitization This homeostatic mechanism protects from toxic effects of hormone excess. Heterologous desensitization occurs when exposure of the cell to one agonist reduces the responsiveness of the cell any other agonist that acts through a different receptor. This most commonly occurs through receptors that act through the adenylyl cyclase system. Heterologous desensitization results in a broad pattern of refractoriness with slower onset than homologous desensitization 14 G protein-coupled receptor kinases (GRKs) Receptor phosphorylation by second messenger kinases A family of serine-threonine kinases that recognizes and phosphorylates receptors in their agonist-stimulated form Consensus sequence: serine/threonine residues surrounded by acidic residues DLEESSSSD Members of the GRK family and their regulation GRK Tissue expression Regulation GRK1 (rhodopsinkinase) Retinal rods and cones Farnesylation of C terminus GRK2 (ARK1) Ubiquitous, brain PIP2 and G binding GRK3 (ARK2) Ubiquitous, in the brain PIP2 and G binding is lower than GRK2 Heterologous desensitization GRK4 Testes; low in brain GRK5 Ubiquitous, brain Modulated by CAM and calcium sen sor proteins GRK6 Ubiquitous, brain palmitoylated cysteine residues GRK7 Retinal cones Geranygeranylated Receptor phosphorylation by GRKs 到處存在 GRK: G-protein receptor kinase homologous-desensitization Regulation of GRK function PIP2-binding through polybasic C ter minus domains and palmitoylated cysteine residues GRK: G-protein coupled receptor kinase Desensitization or Endocytosis of GPCR’s Effected by Phosphorylation 1. The ligand activated receptor can be phosphorylated on select Ser/Thr residues by GRK (e.g. BARK - adrenergic receptor kinase). These phosphorylated residues provide a docking site for arrestin resulting in inactivation/desensitization. 2. In some instances, arrestin binding targets the receptor for clathrin-dependent endocytosis. 3. In addition, if the occupied GPCR leads to cAMP, the receptor can also be phosphorylated by PKA leading to its inactivation/densensitization. Penela P, Ribas C, Mayor F. Jr. Mechanisms of regulation of the expression and function of G protein-coupled receptor kinases. Cell Signal. 2003 Nov;15(11):973-981. 15 Clathrin-dependent Receptor-mediated endocytosis -Arrestins : intracellular protein - Interaction with phosphorylated GPCRs uncouples the receptors from heterotrimeric G proteins, producing a nonsignaling, desensitized receptor. (desensitization ) - Target the GPCRs to clathrin-coated pits for endcytosis to function as docking proteins that link receptors to components of the endocytic machinery such as AP-2 and clathrin (internalization ) - Regulate the dephosphorylation of Receptors (resensitization). GRK-phosphorylation/ arrestin binding : uncouling, desensitization Clathrin coated pit : pinch off –dynamin - sequestration Agonist binding ? The ability of -Arrestins to remain associated with some receptors but not others suggests that -Arrestins may regulate the cellular trafficking and dephosphorylation of receptor and ultimately their kinetics of resensitization. Ref. Seminars in Cell & Developmental biology 9,1998 Receptor Down-Regulation NO-self enzymatic activity Slower onset (hours to days), more prolonged effect Decreased synthesis of receptor proteins Increase in receptor internalization and degradation Internalization involves endocytosis of receptor: the endocytic vesicle may ultimately return the receptor to the cell surface, or alternatively may deliver the receptor to a lysosome for destruction. Endocytic vesicles are associated with phosphatases which can clear phosphate from a receptor and ready it for reuse before returning it to the plasma membrane. -lack intrinsic catalytic activity -binding of the ligand results in the formation of a receptor dimer (2 receptors) -this dimer than activates a class of protein called tyrosine kinases -this activation results in the phosphorylation of downstream targets by these tyrosine kinases (stick phosphate groups onto tyrosines within the target protein) -receptors for cytokines such as, interferons 16 G protein-coupled receptors transmit signals to MAP kinase Two types of intracellular signaling complexes Activated G of Gprotein May also InduceMAPK Cascade Scaffold protein All MAP kinase are serine/threonine kinase Adapter protein: directly contact with receptor Cytokine receptor Two types of intracellular signaling complexes All molecule attached at scaffold protein Cytokine receptors signal to the nucleus in a direct pathway Tyrosine kinase receptors One protein → phosphorylation → induced another protein bind → induced another protein 17 Signaling pathway using modular binding domains Signal protein complexes (more efficiency and specific) Signal cascades are often mediated by large "solid state" assemblies that may include receptors, effectors, and regulatory proteins, linked together in part by interactions with specialized scaffold proteins. Scaffold proteins often interact also with membrane constituents, elements of the cytoskeleton, and adaptors mediating recruitment into clathrin-coated vesicles. They improve efficiency of signal transfer, facilitate interactions among different signal pathways, and control localization of signal proteins within a cell. One strategy the cell uses to achieve specificity involves scaffolding proteins They organize groups of interacting signaling proteins into signaling complexes Because the scaffold guides the interactions between the successive components in such a complex, the signal is relayed with speed Adapter protein: directly contact with receptor; It also signal players In addition, cross-talk between signaling pathways is avoided Scaffolding proteins are large relay proteins to which other relay proteins are attached Scaffolding proteins can increase the signal transduction efficiency Signal molecule Plasma membrane Some receptors and signal transduction protein are localized Clustering of membrane proteins mediated by adapter domains Synaptic junction: chemical signal → presynaptic cell → clustering receptor → raid and efficient signal transmission PDZ: 90 a.a.; target protein Ser-Thr-X-Φ; X: any, Φ: hydorphobic PDZ interact with subunit receptor formed complex Receptor Three different protein kinases Scaffolding protein Protein motifes (1) Mediate protein-protein interactions (2) Determine the location of signaling proteins 18 Different kinds of intracellular signaling proteins along a signaling pathway from the cell surface to the nucleus Signal Transduction Domains Src tyrosine kinase contains 4 functional regions (known function and by homologies with domains in other proteins). The Src Homology (SH) domains have subsequently been defined as -SH1 being the Tyrosine Kinase -SH2 being a domain that binds phosphorylated tyrosines (or PTB) -SH3 being a domain that binds proline rich regions -SH4 being a domain that regulated addition of lipids Thus the domains contain not just enzyme activities but control the formation of protein complexes and anchoring to membranes signal receptor Lipid eg PIP3 PH PP Tyr K YP SH2 P SH3 Relay(接替) proteins: pass the message to the next signaling component Adaptor proteins: link one signaling protein to another without themselves participating in the signaling event Bifurcation (分枝)proteins: spread the signal from one signaling pathway to another Amplifier proteins: usually either enzymes or ion channels that enhance the signal they receive PPP effects Ligand binding to the receptor activates an intrinsic enzymatic activity Receptors with intrinsic enzymatic activity (e.g. guanylate cyclase activity, protein phosphatase, serine/threonine kinase or tyrosine kinase activity) Transducer proteins: convert the signal to a different form e.g. adenyl cyclase Tyrosine kinase receptors these receptors traverse the membrane only once receptor has intrinsic enzyme activity – i.e. the receptor itself is an enzyme respond exclusively to protein stimuli – cytokines – mitogenic growth factors: • platelet derived growth factor • epidermal growth factor These usually receive signals that regulate: cell proliferation (growth factors) or cell differentiation (inducers) Tyrosine kinase activation is key event in intracellular signal transduction. Most Tyrosine Kinase Receptors exist as inactive monomers in membrane. Receptor autophosphorylation occurs via a ligand-induced receptor dimerization Phosphotyrosine residues in cytoplasmic domain of receptor act as docking sites that bind cytoplasmic signaling molecules. 19 There are six major families of receptor tyrosine kinases. All have a TK domain on the cytosolic, COOH-terminal end, A single-pass transmembrane domain, and One or more cysteine-rich or Immunoglobuliu-like ligand-binding domains. Tyrosine kinase: (phosphorylation & dephosphorylation) kinase enzymes add a phosphate (Pi) group tyrosine kinase speicfically to tyrosine residue phosphatases remove Pi phosphorylation state alters shape (conformation) of protein and changes its function – enzyme activity; solute transport; gene expression covalent modification by phosphorylation is extremely important in regulation biological responses • An individual tyrosine-kinase receptor consists of several parts: – an extracellular signal-binding sites, – a single alpha helix spanning the membrane, and – an intracellular tail with several tyrosines. • • • When ligands bind to two receptor polypeptides, the polypeptides aggregate, forming a dimer. This activates the tyrosine-kinase section of both. These add phosphates to the tyrosine tails of the other polypeptide. • • The fully-activated receptor proteins activate a variety of specific relay proteins that bind to specific phosphorylated tyrosine molecules. – One tyrosine-kinase receptor dimer may activate ten or more different intracellular proteins simultaneously. These activated relay proteins trigger many different transduction pathways and responses. 20 Tyrosine Kinase Receptors Tyrosine Kinase Receptors Note steps involved: 1. Ligand Reception 2. Receptor Dimerization 3. Catalysis (Phosphorylization) 4. Subsequent Protein Activation 5. Further Transduction 6. Response The TGFβ superfamily consists of many members. Bone morphogenetic proteins (BMP) is the largest family. TGFβ is formed by cleavage of a secreted inactive precursor During development, TGFβ signaling is involved in pattern formation, cell proliferation, differentiation, ECM production, and cell death. In adults, TGFβ is involved in tissue repair and immune regulation. Maturation of TGFβ is dependent on release from LTBP (latent TGF- – binding protein) by proteolysis. TGF- receptor and the direct activation of Smads 潛在 RII receptor has a constitutive ser/thr kinase activity TGFβ binding induces complex formation between RII and RI; phosphorylation of RI by RII activates RI kinase activity RI did not bind to TGF RI kinases Smad transcription factors. Phosphorylation results in a conformational change in Smad phosphorylation Complexes with other transcriptional factor = Smad4 Complex moves in nucleus Activates gene Gene = plasminogen activator inhibitor no cell growth TGFR: receptor serine/ threonine kinase Smad: transcription factor TFE: transcription factor 21 Membrane Form of Guanylyl Cyclase Atrial natriuretic factor Small hydrophobic signaling molecules, such as steroids, can cross the cell membrane (e.g. estrogen, vitamin D, thyroid hormone, retinoic acid) and bind to intracellular receptors 1. Receptor guanylyl cyclases generate cGMPdirectly as an intracellular mediator 2. Atrial natriuretic peptides (ANPs) are family of related peptide hormones 3. Single pass transmembrane protein that has extracellular binding site for ANPs and an intracellular guanylyl cyclase catalytic unit. 4. Binding of ANP activates cylase to produce cGMP which in turn activates cGMPdependent protein kinase (GKinase) kidney Intracellular receptors (nucleus receptor) intestinal epithelial cells The hormone-receptor complex has an exposed DNA binding site and can activate transcription directly (or, more typically as a homo- or hetero-dimer) This usually initiates a cascade of transcription events GTP cGMP Nuclear Receptors Some signaling molecules that bind to intracellular receptors Lipid soluble ligands that penetrate cell membrane (corticosteroids, mineralocorticoids, sex steroids, Vitamin D, thyroid hormone) Receptors contain DNA-binding domains and act as ligand-regulated transcriptional activators or suppressors(=> characteristic lag period of 30 minutes to several hours): cortisol response Ligand binding of the receptors triggers the formation of a dimeric complex that can interact with specific DNA sequences (=“Response Elements”) to induce transcription. The resulting protein products possess half-lifes that are significantly longer than those of other signaling intermediates => Effects of nuclear receptor agonists can persist for hours or days after plasma concentration is zero. 雌二酮 estradiol 睪固酮 testosterone Vitamin D3 一種腎上腺 皮質內泌素 thyroxine 甲狀腺素 Retinoic acid 維他命A酸 22 Nuclear Receptors • The nuclear receptor superfamily Examples: – Glucocorticoids: Inhibit transcription of COX-2; induce transcription of Lipocortin – Mineralcorticoids: Regulate expression of proteins involved in renal function – Retinoids (Vit A derivatives): Control embryonic development of limbs and organs; affect epidermal differentiation => dermatological use (Acne)痤瘡 粉刺 – PPARs (Peroxisome Proliferation-Activated Receptors): control metabolic processes: • PPAR: Target of Fibrates (cholesterol lowering drugs: stimulate -oxidation of fatty acids) • PPAR: Target of Glitazones (anti-diabetic drugs: induce expression of proteins involved in insulin signaling => improved glucose uptake) Hinge 鉸縺 23 Nuclear Receptor Family is Large but not ubiquitous: mammals have ~50-60 genes flies 21 worms 270 (!!!) plants 0 yeast 0 Only a handful of physiological ligands have been identified, (despite many genes, worms lack any known lipid based endocrine system) Early primary response (A) and delayed secondary response (B) that result from the activation of an intracellular receptor protein Nuclear receptor family (steroid) Steroid hormone receptors are part of the superfamily of nuclear receptors that contains over 30 members. All members have conserved regions of high homology Hormone binding domain 90% homologous 10% difference accounts for specificity DNA binding domain which contains zinc fingers Receptors are found complexed with heat shock proteins (HSP) Unoccupied receptor held in inactive conformation by HSP Ligand binding releases HSP and exposes DNA binding domain Hormone receptor complex then binds to response elements on gene and allows transcription to occur Ligand gate Ion-channel receptors Ligand binding changes the conformation of the receptor so that specific ions flow through it 24 Nongated ion channels and the resting membrane potential Types of Membrane Ionic Channels Non-gated channels: leakage channels open at rest Gated: need ligand to activation; Non-gated: do not need ligand Ion Channel (non-gate) Generation of electrochemical gradient across plasma membrane Gated Channels: – Voltage-gated channels – Mechanically-gated channels – Chemically-gated channels (from outside or inside of the membrane) • Neurotransmitter-activated • Calcium-gated • ATP-gated • Cyclic nucleotide-gated • About 100 different kinds of channels Ligand gate Ion channel characterizations multi-subunit, transmembrane protein complexes complex is both the receptor and ion channel stimuli: chemical, stretch, voltage or light stimulus induces conformational change to open or close ion channel i.e. Ca+ gradient regulation of signal transduction , muscle contraction and triggers secretion of digestive enzyme in to exocrine pancreastic cells i.e. Na+ gradient uptake of a.a , symport, antiport; formed membrane potential i.e. K+ gradient formed membrane potential Q: how does the electrochemical gradient formed? Selective movement of Ions Create a transmembrane electric potential difference Ligand-gate ion channels chemical stimuli bind to receptor and open or close ion channel stimuli can be extracellular or intracellular EXTRACELLULAR STIMULI: (neurotransmitters) – e.g. acetylcholine, dopamine, GABA, glutamate INTRACELLULAR STIMULI: (second messengers) – e.g. IP3, cAMP, cGMP, Ca2+ Light gated channels respond to light; in the eyes Mechanically gated channels respond to vibration or pressure - in the ear, touch 25 NMDA receptor-Ligand gated channel Ion-channel-linked receptors Convert chemical signals ==> electrical signals Extracellular ligand-gated nicotinic ACh (muscle): 2 (embryonic), 2 (adult) nicotinic ACh (neuronal): (2-10), (2-4) glutamate: NMDA, kainate, AMPA P2X (ATP) Many other types of transmembrane ion channels ==> Ion channels are common drug targets! • Voltage-gated channels: • Gating: controlled by membrane polarization/depolarization • Selectivity: Na+, K+ or Ca+ ions 5-HT3 GABAA: (1-6), (1-4), (1-4), , , (1-3) Glycine • Intracellular ligand-gated channels: • Ca+ controlled K+ channel • ATP-sensitive K+ channel • IP3-operated Ca+ channel (in the ER membrane) 26 Voltage gate ion channels ion channel undergoes conformational change folllowing electrical stimulus this “depolarization” opens the channel – leads to flow of Na+ into cell – constitutes an “action potential” channel re-closes Pseudo-subunit vs. true subunit structure Passive-Mediated Transport • Gated vs non-gated • Gated Na+-channel Non-Gated - voltage Na+-channel Na+-channel - chemical K+-channel - voltage K+-channel K+-channel - chemical Cl--channel - chemical Cl--channel 27 Regulation of Ion Channels A wide range of plasma membrane ligand and voltage sensitive ion channels exist controlling cytoplasmic levels of Na+, K+, and Ca2+. For example, acetyl choline receptor allows influx of Na+ and K+ triggering action potential in nerve/muscle Cytoplasmic Ca2+ levels regulated via outer membrane- and ER receptors (ryanodine and IP3) Changes in level of Ca2+ are particularly dramatic (1000 fold increase) A major effector of Ca2+ is calmodulin which activates myosin light chain kinase (hence promotes contraction) and calmodulin dependent protein kinase (metabolism, transcription etc) ATP dependent pumps rapidly transport Ca2+ back to where it came from meaning the signal can be very rapidly shut off then switched on again – eg muscle contraction Intracellular ligand-gated leukotriene C4-gated Ca2+ ryanodine receptor Ca2+ IP3-gated Ca2+ IP4-gated Ca2+ Ca2+-gated K+ Ca2+-gated nonselective cation Ca2+-gated Cl– cAMP cation cGMP cation cAMP chloride ATP Cl– volume-regulated Cl– arachidonic acidactivated K+ Na+-gated K+ G-protein linked receptors coupled to ion channels • • • • • • • • • • • • • • • Acetylcholine (muscarinic) • Adenosine & adenine nucleotides • Adrenaline & noradrenaline • Angiotensin • Bombesin • Bradykinin • Calcitonin • Cannabinoid • Chemokine • Cholecystokinin & gastrin • Dopamine • Endothelin • Galinin • GABA (GABAB) • Glutamate (quisqualate) • Histamine 5-Hydroxytryptamine (1,2) Leukotriene Melatonin Neuropeptide Y Neurotensin Odorant peptides Opioid peptides Platelet-activating factor Prostanoid Protease-activated Tachykinins Taste receptors VIP Vasopressin and oxytocin 28