Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
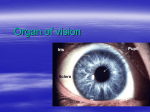
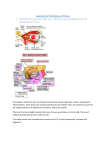
Vision The eyes are composed of an optical portion, which focuses the visual image on the receptor cells, and a neural component, which transforms the visual image into a pattern of neural discharges. The Optics of Vision Light waves are propagated in all directions from every point of a visible object. Before an accurate image of a point on the object is achieved, these divergent light waves must pass through an optical system that focuses them back into a point. In the eye, the image of the object being viewed is focused upon the retina, a thin layer of neural tissue lining the back of the eyeball. The retina contains the light-sensitive receptor cells, the rods and cones, as well as several types of neurons. The lens and cornea of the eye are the optical systems that focus impinging light rays into an image upon the retina. At a boundary between two substances of different densities, such as the cornea and the air, light rays are bent so that they travel in a new direction. The cornea plays a larger quantitative role than the lens in focusing light rays because the rays are bent more in passing from air into the cornea than they are when passing into and out of the lens or any other transparent structure of the eye. The surface of the cornea is curved so that light rays coming from a single point source hit the cornea at different angles and are bent different amounts, directing the light rays back to a point after emerging from the lens. The image is focused on a specialized area known as the fovea centralis, the area of the retina that gives rise to the greatest visual clarity. The image on the retina is upside down relative to the original light source, and it is also reversed right to left. Light rays from objects close to the eye strike the cornea at greater angles and must be bent more in order to reconverge on the retina. Although, as noted above, the cornea performs the greater part quantitatively of focusing the visual image on the retina, all adjustments for distance are made by changes in lens shape. Such changes are part of the process known as accommodation. The shape of the lens is controlled by the ciliary muscle and the tension it applies to the zonular fibers, which attach this smooth muscle to the lens. To focus on distant objects, the zonular fibers pull the lens into a flattened, oval shape. When their pull is removed for near vision, the natural elasticity of the lens causes it to become more spherical. This more spherical shape provides additional bending of the light rays, which is important to focus near objects on the retina. The ciliary muscle, which is stimulated by parasympathetic nerves, is circular, like a sphincter, so that it draws nearer to the lens as it contracts and therefore removes tension on the zonular fibers, resulting in accommodation for viewing near objects. Accommodation also includes other mechanisms that move the lens slightly toward the back of the eye, turn the eyes inward toward the nose (convergence), and constrict the pupil. The sequence of events for accommodation is reversed when distant objects are viewed. The cells that make up most of the lens lose their internal membranous organelles early in life and are thus transparent, but they lack the ability to replicate. The only lens cells that retain the capacity to divide are on the surface of the lens, and as new cells are formed, older cells come to lie deeper within the lens. With increasing age, the central part of the lens becomes denser and stiffer and acquires a coloration that progresses from yellow to black. Since the lens must be elastic to assume a more spherical shape during accommodation for near vision, the increasing stiffness of the lens that occurs with aging makes accommodation for near vision increasingly difficult. This condition, known as presbyopia, is a normal part of the aging process and is the reason that people around 45 years of age may have to begin wearing reading glasses or bifocals for close work. The changes in lens color that occur with aging are responsible for cataract, which is an opacity of the lens and one of the most common eye disorders. Early changes in lens color do not interfere with vision, but is impaired as the process slowly continues. The opaque lens can be removed surgically. With the aid of an implanted artificial lens or compensating eyeglasses, effective vision can be restored, although the ability to accommodate is lost. Cornea and lens shape and eyeball length determine the point where light rays reconverge. Defects in vision occur if the eyeball is too long in relation to the focusing power of the lens. In this case, the images of near objects fall on the retina, but the images of far objects focus at a point in front of the retina. This is a nearsighted, or myopic, eye, which is unable to see distant objects clearly. In contrast, if the eye is too short for the lens, images of distant objects are focused on the retina but those of near objects are focused behind it. This eye is farsighted, or hyperopic, and near vision is poor. Defects in vision also occur where the lens or cornea does not have a smoothly spherical surface, a condition known as astigmatism. These surface imperfections can usually be compensated for by eyeglasses. The lens separates two fluid-filled chambers in the eye, the anterior chamber, which contains aqueous humor, and the posterior chamber, which contains the more viscous vitreous humor. These two fluids are colorless and permit the transmission of light from the front of the eye to the retina. The aqueous humor is formed by special vascular tissue that overlies the ciliary muscle. In some instances, the aqueous humor is formed faster than it is removed, which results in increased pressure within the eye. Glaucoma, the leading cause of irreversible blindness, is a disease in which the axons of the optic nerve die, but it is often associated with increased pressure within the eye. The amount of light entering the eye is controlled by muscles in the ring like, pigmented tissue known as the iris, the color being of no importance as long as the tissue is sufficiently opaque to prevent the passage of light. The hole in the center of the iris through which light enters the eye is the pupil. The iris is composed of smooth muscle, which is innervated by autonomic nerves. Stimulation of sympathetic nerves to the iris enlarges the pupil by causing the radially arranged muscle fibers to contract. Stimulation of parasympathetic fibers to the iris makes the pupil smaller by causing the sphincter muscle fibers, which circle around the pupil, to contract. These neurally induced changes occur in response to light-sensitive reflexes. Bright light causes a decrease in the diameter of the pupil, which reduces the amount of light entering the eye and restricts the light to the central part of the lens for more accurate vision. Conversely, the iris enlarges in dim light, when maximal illumination is needed. Changes also occur as a result of emotion or pain. Photoreceptor Cells The photoreceptor cells in the retina are called rods and cones because of the shapes of their light-sensitive tips. Note that the light-sensitive portion of the photoreceptor cells—the tips of the rods and cones—faces away from the incoming light, and the light must pass through all the cell layers of the retina before reaching the photoreceptors and stimulating them. A pigmented layer (the choroid), which lies behind the retina, absorbs light and prevents its reflection back to the rods and cones, which would cause the visual image to be blurred. The rods are extremely sensitive and respond to very low levels of illumination, whereas the cones are considerably less sensitive and respond only when the light is brighter than, for example, twilight. The photoreceptors contain molecules called photopigments, which absorb light. There are four different photopigments in the retina, one (rhodopsin) in the rods and one in each of the three cone types. Each photopigment contains an opsin and a chromophore. Opsin is a collective term for a group of integral membrane proteins, one of which surrounds and binds a chromophore molecule. The chromophore, which is the actual light-sensitive part of the photopigment, is the same in each of the four photopigments and is retinal, a derivative of vitamin A. The opsin differs in each of the four photopigments. Since each type of opsin binds to the chromophore in a different way and filters light differently, each of the four photopigments absorbs light most effectively at a different part of the visible spectrum. For example, one photopigment absorbs wavelengths in the range of red light best, whereas another absorbs green light best. Within the photoreceptor cells, the photopigments lie in specialized membranes that are arranged in highly ordered stacks, or discs, parallel to the surface of the retina. The repeated layers of membranes in each photoreceptor may contain over a billion molecules of photopigment, providing an effective trap for light. Light activates retinal, causing it to change shape. This change triggers a cascade of biochemical events lead to hyperpolarization of the photoreceptor cell’s plasma membrane and, thereby, decreased release of neurotransmitter (glutamate) from the cell. Note that in the case of photoreceptors the response of the cell a stimulus (light) is a hyperpolarizing receptor potential and a decrease in neurotransmitter release. The decrease in neurotransmitter then causes the bipolar cells, which synapse with the photoreceptor cell, to undergo a hyperpolarization in membrane potential. After its activation by light, retinal changes back to its resting shape by several mechanisms that do not depend on light but are enzyme mediated. Thus, in the dark, retinal has its resting shape, the photoreceptor cell is partially depolarized, and more neurotransmitter is being released. When one steps back from a place of bright sunlight into a darkened room, dark adaptation, a temporary “blindness,” takes place. In the low levels of illumination of the darkened room, vision can only be supplied by the rods, which have greater sensitivity than the cones. During the exposure to bright light, however, the rods’ rhodopsin has been completely activated. It cannot respond fully again until it is restored to its resting state, a process requiring some tens of minutes. Dark adaptation occurs, in part, as enzymes regenerate the initial form of rhodopsin, which can respond to light. Neural Pathways of Vision The neural pathways of vision begin with the rods and cones. These photoreceptors communicate by way of electrical synapses with each other and with second order neurons, the bipolar cell. The bipolar cells synapse (still within the retina) both upon neurons that pass information horizontally from one part of the retina to another and upon the ganglion cells. Via these latter synapses, the ganglion cells are caused to respond differentially to the various characteristics of visual images, such as color, intensity, form, and movement. Thus, a great deal of information processing takes place at this early stage of the sensory pathway. The distinct characteristics of the visual image are transmitted through the visual system along multiple, parallel pathways by two types of ganglion cells, each type concerned with different aspects of the visual stimulus. Parallel processing of information continues all the way to and within the cerebral cortex, to the highest stages of visual neural networks. Ganglion cells are the first cells in the visual system to respond to activation by producing action potentials, whereas the rods and cones and almost all other retinal neurons produce only graded potentials. The axons of the ganglion cells form the output from the retina—the optic nerve, cranial nerve II. The two optic nerves meet at the base of the brain to form the optic chiasm, where some of the fibers cross to the opposite side of the brain, providing both cerebral hemispheres with input from each eye. Optic nerve fibers project to several structures in the brain, the largest number passing to the thalamus, where the information from the different ganglion cell types is kept distinct. In addition to the input from the retina, many neurons of the lateral geniculate nucleus also receive input from the brainstem reticular formation and input relayed back from the visual cortex. These nonretinal inputs can control the transmission of information from the retina to the visual cortex and may be involved in the ability to shift attention between vision and the other sensory modalities. The lateral geniculate nucleus sends action potentials to the visual cortex, the primary visual area of the cerebral cortex. Different aspects of visual information are carried in parallel pathways and are processed simultaneously in a number of independent ways in different parts of the cerebral cortex before they are reintegrated to produce the conscious sensation of sight and the perceptions associated with it. The cells of the visual pathways are organized to handle information about line, contrast, movement, and color. They do not, however, form a picture in the brain. Rather, they form a spatial and temporal pattern of electrical activity. We mentioned that a substantial number of fibers of the visual pathway project to regions of the brain other than the visual cortex. For example, visual information is transmitted to the suprachiasmatic nucleus, which lies just above the optic chiasm and functions as a “biological clock.” Information about diurnal cycles of light intensity is used to entrain this neuronal clock. Other visual information is passed to the brainstem and cerebellum, where it is used in the coordination of eye and head movements, fixation of gaze, and change in pupil size. Color Vision The colors we perceive are related to the wavelengths of light that are reflected, absorbed, or transmitted by the pigments in the objects of our visual world. For example, an object appears red because shorter wavelengths, which would be perceived as blue, are absorbed by the object, while the longer wavelengths, perceived as red, are reflected from the object to excite the photopigment of the retina most sensitive to red. Light perceived as white is a mixture of all wavelengths, and black is the absence of all light. Color vision begins with activation of the photopigments in the cone receptor cells. Human retinas have three kinds of cones, which contain red-, green-, or bluesensitive photopigments. As their names imply, these pigments absorb and hence respond optimally to light of different wavelengths. Because the red pigment is actually more sensitive to the wavelengths that correspond to yellow, this pigment is sometimes called the yellow photopigment. Although each type of cone is excited most effectively by light of one particular wavelength, it responds to other wavelengths as well. Thus, for any given wavelength, the three cone types are excited to different degrees. For example, in response to light of 531-nm wavelengths, the green cones respond maximally, the red cones less, and the blue cones not at all. Our sensation of color depends upon the relative outputs of these three types of cone cells and their comparison by higher-order cells in the visual system. The pathways for color vision follow those described in. Ganglion cells of one type respond to a broad band of wavelengths. In other words, they receive input from all three types of cones, and they signal not specific color but general brightness. Ganglion cells of a second type code specific colors. These latter cells are also called opponent color cells because they have an excitatory input from one type of cone receptor and an inhibitory input from another. The cell gives a weak response when stimulated with a white light because the light contains both blue and red wavelengths. Other more complicated patterns also exist. The output from these cells is recorded by multiple—and as yet unclear—strategies in visual centers of the brain. At high light intensities, as in daylight vision, most people—92 percent of the male population and over 99 percent of the female population—have normal color vision. People with the most common kind of color blindness—a better term is color deficiency—either lack the red or green cone pigments entirely or have them in an abnormal form; as a result, they have trouble perceiving red versus green. Eye Movement The cones are most concentrated in the fovea centralis, and images focused there are seen with the greatest acuity. In order to focus the most important point in the visual image (the fixation point) on the fovea and keep it there, the eyeball must be able to move. Six skeletal muscles attached to the outside of each eyeball control its movement. These muscles perform two basic movements, fast and slow. The fast movements, called saccades, are small, jerking movements that rapidly bring the eye from one fixation point to another to allow search of the visual field. In addition, saccades move the visual image over the receptors, thereby preventing adaptation. Saccades also occur during certain periods of sleep when the eyes are closed, and may be associated with “watching” the visual imagery of dreams. Slow eye movements are involved both in tracking visual objects as they move through the visual field and during compensation for movements of the head. The control centers for these compensating movements obtain their information about head movement from the vestibular system, which will be described shortly. Control systems for the other slow movements of the eyes require the continuous feedback of visual information about the moving object.