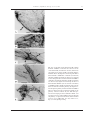
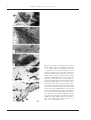
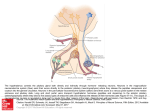
Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
Aqua-BioScience Monographs Vol. 3, No. 2, pp. 39–72 (2010) www.terrapub.co.jp/onlinemonographs/absm/ Reproductive Biology of Salmoniform and Pleuronectiform Fishes with Special Reference to Gonadotropin-Releasing Hormone (GnRH) Masafumi Amano School of Marine Biosciences, Kitasato University Ofunato, Iwate 022-0101, Japan e-mail: [email protected] Abstract A salmonid fish, masu salmon Oncorhynchus masou, has salmon gonadotropin-releasing hormone (sGnRH) and chicken GnRH-II (cGnRH-II), while a pleuronectiform fish, barfin flounder Verasper moseri, has sGnRH, cGnRH-II and seabream GnRH (sbGnRH). In masu salmon, sGnRH-immunoreactive (ir) cell bodies are scattered from the olfactory nerve through the ventral telencephalon (VT) and the preoptic area (POA). cGnRH-II-ir cell bodies are located in the midbrain tegmentum (MT). sGnRH but not cGnRH-II is detected in the pituitary. sGnRH peptide levels and sGnRH mRNA levels in the VT and the POA increased during gonadal maturation. sGnRH neurons are derived from the olfactory epithelium and migrate into the brain. In barfin flounder, sGnRH-ir, cGnRH-II-ir, and sbGnRH-ir cell bodies are located in the olfactory bulbs and the terminal nerve ganglion (TN), the MT, and the POA, respectively, and these neurons do not migrate in the brain. sbGnRH is detected in the pituitary. sbGnRH mRNA levels in the brain increased during gonadal maturation. Although three GnRH systems exist in the barfin flounder, anatomical distinction between the TN- and the POA-GnRH systems is not clear in masu salmon. Thus, it is suggested that sGnRH neurons in masu salmon play different roles according to the location in the brain. 1. General introduction Gonadotropin-releasing hormone (GnRH) is a decapeptide originally isolated from pig and sheep hypothalami as a physiologic regulator of luteinizing hormone (LH) release from the pituitary (Matsuo et al. 1971; Burgus et al. 1972). At present, it is generally accepted that GnRH regulates synthesis and release of pituitary gonadotropin (GTH) (see King and Millar 1992; Sherwood et al. 1993). It has been shown that two or three molecular forms of GnRH exist, even within the same species (Oka 1997; Okuzawa and Kobayashi 1999; Okubo and Nagahama 2008). In addition, GnRH can act as a neuromodulator and has also been implicated in reproductive behavior in many species including teleost fish such as dwarf gourami Colisa lalia (Yamamoto et al. 1997) and goldfish Carassius auratus (Volkoff and Peter 1999). © 2010 TERRAPUB, Tokyo. All rights reserved. doi:10.5047/absm.2010.00302.0039 Received on March 31, 2010 Accepted on June 24, 2010 Online published on August 31, 2010 Keywords • GnRH • brain • pituitary • gonad • radioimmunoassay • immunohistochemistry • in situ hybridization • quantitative PCR • masu salmon • sockeye salmon • barfin flounder To date, 15 forms of GnRHs have been identified based on their primary structure or complementary DNAs (cDNAs) in vertebrates, as shown in Fig. 1 (Okubo and Nagahama 2008; Kavanaugh et al. 2008). GnRH forms are traditionally named after the species from which they were first identified. In addition to vertebrate species, GnRH was isolated in invertebrates: e.g., in the protochordate Ciona intestinalis (Powell et al. 1996; Adams et al. 2003), octopus Octopus vulgaris (Iwakoshi et al. 2002) and the sea hare Aplysia californica (Zhang et al. 2008). In mammalian, avian, reptilian, and amphibian animals, GnRH is conveyed to the pituitary via the hypothalamo-hypophyseal portal vessels (Fig. 2A). In mammals, pulsatile release of mammalian GnRH (mGnRH) by hypothalamic neurons stimulates GTH secretion from the pituitary. However, teleost fishes lack the median eminence. Instead, GnRH neurons are found to directly innervate the pituitary (Fig. 2B). It is therefore interesting to examine GnRH systems in teleost fish in view of comparative endocrinology. 40 M. Amano / Aqua-BioSci. Monogr. 3: 39–72, 2010 Fig. 1. The primary structure of the 15 known molecular forms of GnRH in vertebrates. All GnRH forms are composed of 10 amino acids and contain an N-terminal pyroglutamate and C-terminal glycinamide. GnRH forms are traditionally named after the species from which they were first identified. Fig. 2. Schematic drawing of the brain–pituitary–gonadal system in (A) mammals and (B) teleost fishes. Understanding of reproductive mechanism of teleost fish is necessary for the establishment and development of fish aquaculture. GnRH is considered to play an important role in fish reproduction as in other vertebrate species. This monograph will focus on the reproductive biology of salmoniform and pleuronectiform fishes with special reference to GnRH, with emphasis on the author’s own work. Masu salmon, Oncorhynchus masou (Figs. 3A–C), and sockeye salmon, Oncorhynchus nerka, are used as a model of salmoniform. The masu salmon used were offspring of wild fish which had migrated to the Shiribetsu River (Hokkaido). Wild masu salmon migrate to the sea in the spring (1.5 years old), and return to the river in May after a one-year stay in the sea; they spawn in autumn and then die. The masu salmon used in this experiment also smoltified at 1.5 years old and matured at 3 years old in fresh water, although the growth rate was not very rapid. There is a landlocked form of masu salmon called “yamame.” This variety for the most part does not smoltify. Barfin flounder, Verasper moseri, is used as a model of pleuronectiform (Fig. 3D). This species is a large, multiple-spawning pleuronectiform fish inhabiting cold sea basins around east Hokkaido, Japan, and is promising for aquaculture and resource enhancement in northern Japan due to its high commercial value. As for the general information of GnRH in teleost fish, please consult recent reviews (Lethimonier et al. 2004; Guilgur et al. 2006; Okubo and Nagahama 2008; Oka 2009; Zohar et al. 2010). doi:10.5047/absm.2010.00302.0039 © 2010 TERRAPUB, Tokyo. All rights reserved. M. Amano / Aqua-BioSci. Monogr. 3: 39–72, 2010 41 Fig. 3. (A) Immature masu salmon, (B) precocious male masu salmon, (C) masu salmon smolt, and (D) barfin flounder. Bars indicate 5 cm. 2. Identification of GnRH forms in the brain 2-1. Identification of GnRH forms by HPLC and RIA in masu salmon Since two or three molecular forms of GnRH exist, even within the same species (see Oka 1997; Okuzawa and Kobayashi 1999; Okubo and Nagahama 2008), it is quite important to identify GnRH forms in the brain to understand the reproductive biology of fish. The presence of two forms of GnRH, salmon GnRH (sGnRH) and chicken GnRH-II (cGnRH-II), in the teleost brain was first reported in goldfish by employing reversed phase high performance liquid chromatography (rpHPLC) in doi:10.5047/absm.2010.00302.0039 © 2010 TERRAPUB, Tokyo. All rights reserved. 42 M. Amano / Aqua-BioSci. Monogr. 3: 39–72, 2010 conjunction with radioimmunoassay (RIA) (Yu et al. 1988). Thus, GnRH molecules present in the brain of masu salmon were identified by rpHPLC in conjunction with specific RIAs according to Okuzawa et al. (1990). Major peaks were obtained in the fraction corresponding to the retention time of sGnRH and cGnRH-II in sGnRH RIA and cGnRH-II RIA, respectively. A minor peak was also obtained in the fraction corresponding to the retention time of cGnRH-II in sGnRH RIA (Fig. 4). These results indicate that masu salmon brain contained a peptide chromatographically and immunologically identical to sGnRH and cGnRH-II (Amano et al. 1992). Later, sGnRH cDNA was cloned from masu salmon brain (Suzuki et al. 1992). Fig. 4. Reverse-phase HPLC of masu salmon brain extract followed by (A) sGnRH RIA and (B) cGnRH-II RIA. Arrows indicate the elution time of synthetic sGnRH and cGnRH-II. The mobile phase was CH3CN (acetonitrile) containing 0.1% TFA. Reprinted with permission from Zoological Science, 9, Amano et al., Changes in salmon GnRH and chicken GnRH-II contents in the brain and pituitary, and GTH contents in the pituitary in female masu salmon, Oncorhynchus masou, from hatching through ovulation, 375–386, Fig. 2, © 1992, Zoological Society of Japan. 2-2. Identification of GnRH forms by cDNA cloning in barfin flounder The cDNA coding for mGnRH was initially isolated from human placenta (Seeburg and Adelman 1984) and, subsequently, mGnRH genes have been cloned from several species (Adelman et al. 1986; Mason et al. 1986). Chicken GnRH-I (cGnRH-I) gene has been cloned from chicken (Dunn et al. 1993). In teleost fish, the cDNA coding for sGnRH was first isolated from African cichlids Haplochromis burtoni (Bond et al. 1991). Then, the cDNA for cGnRH-II has been isolated from several teleost fish, e.g., African cichlids (White et al. 1994), African catfish Clarias gariepinus (Bogerd et al. 1994) and goldfish (Lin and Peter 1996), the cDNA for sGnRH from several teleost fish, e.g., masu salmon (Suzuki et al. 1992), Atlantic salmon Salmo salar (Klungland et al. 1992), red seabream Pagrus major (Okuzawa et al. 1994), plainfin midshipman Porichthys notatus (Grober et al. 1995), sockeye salmon (Ashihara et al. 1995), goldfish (Lin and Peter 1996), cDNA for catfish GnRH (cfGnRH) from African catfish (Bogerd et al. 1994), and cDNA for seabream GnRH (sbGnRH) from several teleost fish, e.g., African cichlid (White et al. 1995) and from red seabream (Okuzawa et al. 1997). In general, a GnRH precursor is composed of a signal peptide (SP), GnRH and a GnRHassociated peptide (GAP), which is connected to GnRH by a Gly–Lys–Arg sequence. Here, GnRHs in barfin flounder were identified by isolation of their cDNAs. Single-strand cDNA was reverse transcribed from barfin flounder brain poly (A) + RNA. A degenerate forward primer was synthesized based on highly conserved amino acid sequences of GnRH. The 3′ end of the GnRH cDNA was cloned by rapid amplification of cDNA ends (RACE) using the degenerated primer. The 5′ end of the GnRH cDNA was then cloned by 5′-RACE. PCR-amplified cDNA was inserted into pT7 Blue T (Novagen, Madison, WI) and sequenced using the Dye Terminator Cycle Sequencing Ready Reaction Kit (Applied Biosystems, Foster City, CA). The nucleotide sequence was determined using a 373 DNA sequencer (Applied Biosystems). Barfin flounder had three molecular forms of GnRH; sGnRH, cGnRH-II, and sbGnRH. Each GnRH cDNA encoded a SP, GnRH and a GAP. The sGnRH cDNA encoded a SP composed of 23 amino acids and a GAP composed of 54 amino acids (Fig. 5). The cGnRH-II cDNA encoded a SP of 23 amino acids and a GAP of 49 amino acids (Fig. 6). The sbGnRH cDNA encoded a SP of 26 amino acids and a GAP of 57 amino acids (Fig. 7) (Amano et al. 2002a). The existence of multiple forms of GnRH in the brain of masu salmon and barfin flounder suggests that each GnRH has a different distribution and function. Therefore, the physiological role of each GnRH form should be clarified. doi:10.5047/absm.2010.00302.0039 © 2010 TERRAPUB, Tokyo. All rights reserved. M. Amano / Aqua-BioSci. Monogr. 3: 39–72, 2010 43 Fig. 5. Nucleotide sequences of the cDNA encoding the sGnRH precursor of barfin flounder brain and the deduced amino acid sequence of the sGnRH precursor. Nucleotides are numbered from 5′ to 3′, beginning with the initiator codon (ATG) in the coding region for the opening reading frame. Amino acid residues are numbered with the first residue (Met) in the open reading frame. The asterisk indicates the stop codon. The nucleotides corresponding to the polyadenylation signal in the 3′-untranslated region (AATAAA) are underlined. Reprinted from General and Comparative Endocrinology, 126, Amano et al., Molecular cloning of three cDNAs encoding different GnRHs in the brain of barfin flounder, 325–333, © 2002, Elsevier Science (USA), with permission from Elsevier. 3. Distribution of GnRH in the brain and pituitary 3-1. Differential distribution of multiple forms of GnRH in discrete brain areas Differential distribution of multiple forms of GnRH in discrete brain areas has been examined by RIA in several fishes in order to clarify their functions in the brain. Okuzawa et al. (1990) first measured the sGnRH and cGnRH-II contents in the discrete brain regions of the rainbow trout, Oncorhynchus mykiss, using specific RIAs. These authors found that the contents of both forms of GnRHs varied in different brain regions. The levels of sGnRH were higher than those of cGnRH-II in the olfactory bulbs (OB), the telencephalon including preoptic area (POA), the hypothalamus, the optic tectumthalamus and the pituitary, whereas the cerebellum and the medulla oblongata contained much more cGnRH-II than sGnRH. Especially of note, cGnRH-II was undetectable in the pituitary. These results suggest that of the two GnRHs only sGnRH is involved in GTH secretion. In the goldfish, sGnRH was distributed in a larger amount in the OB, the telencephalon, the hypothalamus, and the pituitary than in the other regions, whereas cGnRH-II was distributed widely throughout the brain with highest concentrations in the medulla oblongata. The major difference between salmonid fishes and goldfish is that goldfish pituitary contains cGnRH-II (Kobayashi et al. 1992, 1994). In the European eel, Anguilla anguilla, mGnRH levels were higher than cGnRH-II levels in the pituitary, the olfactory lobes together with the telencephalon, and the diencephalon together with the mesencephalon, while the opposite results were obtained for the posterior part of the brain. Of interest, cGnRH-II levels in the pituitary were slightly above the detectable limit (Dufour et al. 1993). These studies indicated that sGnRH (or mGnRH) and cGnRH-II are differently distributed in the brain and also necessitated the investigation of the localization of GnRH neurons and the changes in GnRH levels during gonadal maturation. Existence of multiple forms of GnRH was demonstrated in the brain of masu salmon and barfin flounder, as shown in the previous section. Thus, differential distribution of multiple forms of GnRH in discrete brain areas of masu salmon and barfin flounder was examined by RIA. Extraction of GnRH from the discrete brain tissue (Figs. 8A, 9A) was done according to Okuzawa et al. (1990). sGnRH, cGnRH-II, and sbGnRH contents were measured by respective RIAs established by Okuzawa et al. (1990) and Senthilkumaran et al. (1999). Concentrations of sGnRH and cGnRH-II (pg/mg tissue) in each region of the brain of ovulated female masu salmon are shown in Fig. 8B. The concentrations of sGnRH and cGnRH-II varied in different brain regions. The levels of sGnRH were higher than those of cGnRH-II doi:10.5047/absm.2010.00302.0039 © 2010 TERRAPUB, Tokyo. All rights reserved. 44 M. Amano / Aqua-BioSci. Monogr. 3: 39–72, 2010 Fig. 6. Nucleotide sequences of the cDNA encoding the cGnRH-II precursor of barfin flounder brain and the deduced amino acid sequence of the cGnRH-II precursor. For details, see the legend to Fig. 5. Reprinted from General and Comparative Endocrinology, 126, Amano et al., Molecular cloning of three cDNAs encoding different GnRHs in the brain of barfin flounder, 325–333, © 2002, Elsevier Science (USA), with permission from Elsevier. Fig. 7. Nucleotide sequences of the cDNA encoding sbGnRH precursor of barfin flounder brain and the deduced amino acid sequence of the sbGnRH precursor. For details, see the legend to Fig. 5. Reprinted from General and Comparative Endocrinology, 126, Amano et al., Molecular cloning of three cDNAs encoding different GnRHs in the brain of barfin flounder, 325–333, © 2002, Elsevier Science (USA), with permission from Elsevier. doi:10.5047/absm.2010.00302.0039 © 2010 TERRAPUB, Tokyo. All rights reserved. M. Amano / Aqua-BioSci. Monogr. 3: 39–72, 2010 Fig. 8. (A) Schematic diagram of a sagittal section of masu salmon brain. The letters a–g represent the following brain areas: a, olfactory bulb; b, telencephalon including preoptic area; c, hypothalamus; d, optic tectum-thalamus including midbrain; e, cerebellum; f, medulla oblongata; g, pituitary. Reprinted with permission from Zoological Science, 14, Amano et al., Distribution and function of gonadotropin-releasing hormone (GnRH) in the teleost brain, 1–11, Fig. 2, © 1997, Zoological Society of Japan. (B) The concentration (pg/mg tissue) of sGnRH and cGnRH-II in discrete areas of the brain and pituitary of ovulated masu salmon (mean ± SEM). The letters a–g represent the same areas as indicated in (A). in the OB, the telencephalon including POA, the hypothalamus, the optic tectum-thalamus and the pituitary, whereas the medulla oblongata contained much more cGnRH-II than sGnRH. Especially of note, cGnRH-II was undetectable in the pituitary. In barfin flounder, the dominant form of GnRH in the pituitary was sbGnRH; sbGnRH levels were much higher than the sGnRH and cGnRH-II levels (Fig. 9B). sGnRH levels were high in the anterior part of the brain, especially in the OB. cGnRH-II levels were high in the posterior part of the brain, especially in the medulla oblongata. Levels of sbGnRH were extremely low in all regions of the brain compared to those of sGnRH and cGnRH-II; sbGnRH was below the detectable limit in the cerebellum and the medulla oblongata (Amano et al. 2002b). These results suggest that sGnRH and sbGnRH are involved in GTH secretion in masu salmon and barfin flounder, respectively. Judging from the wide distribution of sGnRH and cGnRH-II in the brain of masu salmon and barfin flounder, it is also suggested that sGnRH and cGnRH-II function as a neuromodulator in the brain. 45 Fig. 9. (A) Schematic diagram of a sagittal section of barfin flounder brain. The letters a–g represent the following brain areas: a, olfactory bulb; b, telencephalon including preoptic area; c, hypothalamus; d, optic tectum-thalamus including midbrain; e, cerebellum; f, medulla oblongata; g, pituitary. (B) The concentration (pg/mg tissue) of sGnRH, cGnRH-II and sbGnRH in discrete areas of the brain and pituitary of barfin flounder (mean ± SEM). The letters a–g represent the same areas as indicated in (A). With kind permission from Springer Science+Business Media: Cell and Tissue Research, Three GnRH systems in the brain and pituitary of a pleuronectiform fish, barfin flounder Verasper moseri, 309, 2002, 323–329, Amano et al., Figs. 1 and 2, © 2002, Springer-Verlag. 3-2. Localization of GnRH neurons in the brain An approach requisite for clarification of the function of each GnRH-immunoreactive (ir) neuronal group is to examine its projection area in the brain. Immunohistochemistry (IHC) has been used for this purpose. Unfortunately, most of these studies used antiserum against mGnRH. Furthermore, it was clarified that more than one form of GnRH molecule exists, even within the same species. Thus, the distribution of sGnRH- and cGnRH-IIir cell bodies and fibers in the brain of masu salmon and that of sGnRH-, cGnRH-II-, and sbGnRH-ir cell bodies and fibers in the brain of barfin flounder were examined by IHC using specific antibodies for sGnRH, cGnRH-II, and sbGnRH. The distribution of sGnRH-ir cell bodies and fibers, and that of cGnRH-II-ir cell bodies and fibers in the masu salmon are summarized in Fig. 10. sGnRH-ir cell bodies were scattered in the olfactory nerve (ON) (Fig. 11A), the OB (Fig. 11B), between the OB and the telencephalon which corresponds to the terminal nerveganglion (TN) doi:10.5047/absm.2010.00302.0039 © 2010 TERRAPUB, Tokyo. All rights reserved. 46 M. Amano / Aqua-BioSci. Monogr. 3: 39–72, 2010 Fig. 10. (A) Schematic drawing of the distribution of sGnRH-ir cell bodies (closed circles) and fibers (lines) in a sagittal section of masu salmon. (B) Schematic drawing of the distribution of cGnRH-II-ir cell bodies (closed circles) and fibers (lines) in a sagittal section of masu salmon. C, cerebellum; M, medulla oblongata; MT, midbrain tegmentum; OB, olfactory bulb; ON, olfactory nerve; OpN, optic nerve; OT, optic tectum; PIT, pituitary; SV, saccus vasculocus; T, telencephalon. Reprinted with permission of John Wiley & Sons, Inc. from Journal of Comparative Neurology, 314, Amano et al., Immunocytochemical demonstration of salmon GnRH and chicken GnRH-II in the brain of masu salmon, Oncorhynchus masou, 587–597, © 1991, Wiley-Liss, Inc., a Wiley Company. (Figs. 11C, D), the ventral telencephalon (VT) (Figs. 11E, F), and the POA (Figs. 11G, H). sGnRH-ir fibers were distributed in various brain regions from the OB to the spinal cord. sGnRH-ir fibers directly innervated the pituitary (Fig. 11I). cGnRH-II-ir cell bodies were found in the midbrain tegmentum (MT) located rostral to the motoneurons of the oculomotor nerve (Figs. 11J, K). The distribution of cGnRH-II-ir fibers was basically similar to that of sGnRH-ir fibers except for the absence of cGnRH-II-ir fibers in the pituitary (Fig. 11L). The number of cGnRH-II-ir fibers in the brain was much fewer than those of sGnRH. The distribution of sGnRH-ir cell bodies in chum salmon, Oncorhynchus keta, was later reported and was consistent with that in masu salmon (Kudo et al. 1996). These results suggest that, in salmonid, sGnRH not only regulates GTH secretion in the pituitary but also functions as a neuromodulator in the brain, whereas cGnRH-II functions only as a neuromodulator (Amano et al. 1991). The distribution of sGnRH-, cGnRH-II-, and sbGnRH-ir cell bodies and fibers in the barfin flounder is summarized in Fig. 12. sGnRH-ir cell bodies were located in the ventromedial part of the rostral OB and in the TN, and distinct bundles of axons connecting these two regions were observed (Fig. 13A). sGnRH-ir fibers were observed throughout the brain excluding the pituitary. cGnRH-II-ir cell bodies were located in the MT (Fig. 13B) almost the same rostrocaudal levels as nucleus of the medial longitudinal fasciculus (nMLF) neurons, but they seem to constitute a separate cell group as in dwarf gourami (Yamamoto et al. 1998). cGnRH-II-ir fibers were observed throughout the brain excluding the pituitary (Fig. 13B). sbGnRH-ir cell bodies were located in the POA (Fig. 13C). sbGnRH-ir fibers were localized mainly in the POA–hypothalamus–pituitary which formed a distinctive bundle of axons projecting to the pituitary (Fig. 13D), and were not distributed in the other areas. In the pituitary, sbGnRH-ir fibers were observed in the proximal pars distalis (PPD) (Fig. 13E). Thus, it is strongly suggested that sbGnRH is by far the best candidate GnRH that is involved in gonadal maturation by stimulating GTH secretion in barfin flounder. Indeed, some sbGnRH-ir fibers were in close apposition with GTH cells mainly in the PPD of the pituitary in doi:10.5047/absm.2010.00302.0039 © 2010 TERRAPUB, Tokyo. All rights reserved. M. Amano / Aqua-BioSci. Monogr. 3: 39–72, 2010 47 Fig. 11. (A) Sagittal section through the transitional area between the ON and the OB of masu salmon. sGnRH-ir cell bodies (arrowheads) are observed. (B) Sagittal section through the OB. sGnRH-ir cell body (arrowhead) is observed in the ventral part of the OB of masu salmon. A bundle of sGnRH-ir fibers (double arrowhead) arises from the sGnRH-ir cell bodies in the OB. (C) Frontal section through the ventral OB of masu salmon. sGnRH-ir cell bodies (arrowheads) and bundles of sGnRH-ir fibers (double arrowheads) are observed. (D) Sagittal section through the transitional area between the OB and the telencephalon of masu salmon. sGnRH-ir cell body (arrowhead) is observed in the most ventral part. (E) Sagittal section through the VT of masu salmon. sGnRH-ir cell bodies (arrowheads) are observed. (F) Frontal section through the VT of masu salmon. sGnRH-ir cell bodies (arrowheads) are observed. (G) Sagittal section through the POA of masu salmon. sGnRH-ir cell bodies (arrowheads) are observed in the nucleus preopticus parvicellularis anterioris (PPa). (H) Frontal section through the POA of masu salmon. sGnRH-ir cell bodies (arrowheads) are observed in the nucleus preopticus magnocellularis, pars magnocellularis (PMm). (I) Sagittal section through the pituitary of masu salmon. sGnRH-ir fibers are observed in the vicinity of GTH cells of the proximal pars distalis of the pituitary. (J) Sagittal section through the MT of masu salmon. cGnRH-II-ir cell bodies (arrowheads) are observed. (K) Frontal section through the MT of masu salmon. cGnRHII-ir cell body (arrowhead) is observed in the nMLF. (L) Sagittal section through the pituitary of masu salmon. No cGnRH-II-ir fibers are observed. Bars indicate 100 µm. Reprinted with permission of John Wiley & Sons, Inc. from Journal of Comparative Neurology, 314, Amano et al., Immunocytochemical demonstration of salmon GnRH and chicken GnRH-II in the brain of masu salmon, Oncorhynchus masou, 587–597, © 1991, Wiley-Liss, Inc., a Wiley Company. doi:10.5047/absm.2010.00302.0039 © 2010 TERRAPUB, Tokyo. All rights reserved. 48 M. Amano / Aqua-BioSci. Monogr. 3: 39–72, 2010 Fig. 12. (A) Schematic drawing of the distribution of sGnRH-ir cell bodies (closed circles) and fibers (lines) in a sagittal section of barfin flounder. (B) Schematic drawing of the distribution of cGnRH-II-ir cell bodies (closed circle) and fibers (lines) in a sagittal section of barfin flounder. (C) Schematic drawing of the distribution of sbGnRH-ir cell bodies (closed circle) and fibers (lines) in a sagittal section of barfin flounder. C, cerebellum; M, medulla oblongata; MT, midbrain tegmentum; OB, olfactory bulb; ON, olfactory nerve; OpN, optic nerve; OT, optic tectum; PIT, pituitary; POA, preoptic area; T, telencephalon. With kind permission from Springer Science+Business Media: Cell and Tissue Research, Three GnRH systems in the brain and pituitary of a pleuronectiform fish, barfin flounder Verasper moseri, 309, 2002, 323–329, Amano et al., Fig. 3, © 2002, Springer-Verlag. a pleuronectiform fish Japanese flounder Paralichthys olivaceus (Pham et al. 2007). It is also suggested that sGnRH and cGnRH-II function as neuromodulators in the brain, because both sGnRH-ir fibers and cGnRH-II-ir fibers were distributed widely in the brain but not in the pituitary (Amano et al. 2002b). 4. Effects of GnRH on GTH secretion 4-1. Effec ts of GnRH on GTH release in sockeye salmon In masu salmon, it is suggested that sGnRH not only regulates GTH synthesis and release from the pituitary, but also functions as a neuromodulator in the brain, whereas cGnRH-II functions only as a neuromodulator, because cGnRH-II was not detected in the pituitary (Amano et al. 1991). sGnRH and cGnRH-II have been detected also in sockeye salmon brain by rpHPLC and RIA, and their distributions in the brain and pituitary were similar to those of masu salmon (data not shown). The absence of cGnRH-II in the pituitary in salmonid species suggests that cGnRH-II would not be involved in the stimulation of the pituitary GTH in physiological conditions. Two forms of teleostean GTHs, GTH-I and GTHII, was first isolated from chum salmon (Suzuki et al. 1988a, b, c; Kawauchi et al. 1989). Then, they have been isolated from several fish species including salmonids such as coho salmon Oncorhynchus kisutch (Swanson et al. 1989) and masu salmon (Gen et al. 1993; Kato et al. 1993). GTH-I and GTH-II are now commonly referred to as the follicle stimulating hormone (FSH) and luteinizing hormone (LH), respectively, as in mammals (Kawauchi and Sower 2006). In goldfish, both sGnRH and cGnRH-II have the potency to stimulate GTH (possibly LH) release in a dose-dependent manner (Chang et al. 1990, 1991; Habibi 1991). In contrast to salmonids, goldfish possess both forms of GnRH in the pituitary (Yu et al. 1987, 1988; Kim et al. 1995). Therefore, it is interesting to examine whether both GnRHs have the potency to stimulate FSH and LH secretion in salmonid species. Thus, the effects of sGnRH and cGnRH-II on GTH subunits, FSH β and LHβ, release from the superfused pituitary of mature female sockeye salmon were examined by newly developed sensitive and specific RIAs for FSH β and LH β. Pituitaries were dissected into two parts for culture with either sGnRH or cGnRH-II. Half of the pituitaries were pooled and dissected into fragments, and were packed in the superfusion medium. Fractions were collected every 10 minutes in an automatic fraction collector set at 13°C. Fragments were exposed sequentially to 10 minute pulses of six concentrations (0.1, 1, 10, 100, 1000, 10000 nM) of GnRH administered at 80 minute intervals. Exposure of 10 minute pulses of various concentrations of sGnRH and cGnRH-II resulted in a rapid increase of FSHβ and LHβ from perfused fragments of the pituitary (Fig. 14). Thus, it is ascertained that both GnRHs have the potency to stimulate the release of both GTHs from sockeye salmon pituitary. However, cGnRHII was not detected in the pituitary of sockeye salmon by RIA and IHC (data not shown). Therefore, it is possible that cGnRH-II has no involvement in GTH release in this species. It may be a pharmacological effect according to the similarity of amino acid sequences. It is reported in masu salmon that sGnRH stimulates release of LH but not FSH in spawning fish, whereas sGnRH stimulates release of FSH and LH in pre-spawning fish in vitro (Ando et al. 2004). Moreover, sGnRH increases the amount of FSH β mRNAs but not LHβ mRNAs in pre-spawning fish in vitro (Ando doi:10.5047/absm.2010.00302.0039 © 2010 TERRAPUB, Tokyo. All rights reserved. M. Amano / Aqua-BioSci. Monogr. 3: 39–72, 2010 Fig. 13. (A) Sagittal section through the OB of barfin flounder. sGnRH-ir cell bodies are observed in the ventromedial OB (arrowhead on the left) and the TN (arrowhead on the right). Double arrowhead indicates sGnRH-ir fibers. (B) Sagittal section through the MT of barfin flounder. cGnRH-II-ir cell bodies (arrowheads) and fibers (double arrowhead) are observed. (C) Sagittal section through the POA of barfin flounder. sbGnRH-ir cell bodies (arrowheads) and fibers (double arrowhead) are observed. (D) Sagittal section through the POA of barfin flounder. Bundles (one on each side of the brain) of sbGnRH-ir fibers (double arrowheads) arise from the POA area and run toward the pituitary. (E) Sagittal section through the pituitary of barfin flounder. sbGnRH-ir fibers are observed in the proximal pars distalis of the pituitary. The bars indicate 50 µm. With kind permission from Springer Science+Business Media: Cell and Tissue Research, Three GnRH systems in the brain and pituitary of a pleuronectiform fish, barfin flounder Verasper moseri, 309, 2002, 323–329, Amano et al., Fig. 5, © 2002, Springer-Verlag. doi:10.5047/absm.2010.00302.0039 © 2010 TERRAPUB, Tokyo. All rights reserved. 49 50 M. Amano / Aqua-BioSci. Monogr. 3: 39–72, 2010 4-2. Effects of GnRH antagonist on GTH levels in masu salmon and sockeye salmon Fig. 14. Changes in released (A) FSHβ (ng/ml) and (B) LHβ (ng/ml) stimulated by various concentrations of sGnRH, and (C) FSHβ (ng/ml) and (D) LHβ (ng/ml) stimulated by various concentrations of cGnRH-II in sockeye salmon. et al. 2004). Thus, it is suggested that the effects of GnRH on GTH secretion depends on reproductive stage of fish. Unlike tetrapods, teleosts lack a hypothalamohypophysial portal vascular system (Fig. 2). The GnRH neurons directly innervate the pituitary, and GnRH is considered to be released either from nerve terminals that are located near or in direct synaptic contact with pituitary cells or from terminals separated from the pituitary cells by a basement membrane (Peter et al. 1990). This renders it impossible to measure GnRH levels in the hypothalamo-hypophysial portal blood, or to examine GnRH release in vivo (Okuzawa and Kobayashi 1999). Based on the changes of GTH in the pituitary and the plasma in salmonid fishes, it is speculated that FSH stimulates early gonadal development and LH modulates the later stages (Suzuki et al. 1988c; Naito et al. 1991; Tyler et al. 1991; Prat et al. 1996). However, there is no direct evidence for endogenous sGnRH-stimulated GTH secretion in salmonid fishes. The two classical approaches used to study the role of neuroendocrine factors in the regulation of pituitary function are immunoneutralization (a method used to examine the role of a hormone by diminishing its potency with the injection of its antibody) and antagonistic inhibition. However, it is not practical to use immunoneutralization in teleosts due to the direct innervation of gonadotrophs by neurosecretory fibers (Peter et al. 1990; Kah et al. 1993). Thus, GnRH antagonist treatment is considered to be effective for examining the neuroendocrinological function of endogenous GnRH. GnRH antagonist, [Ac∆3-Pro1, 4FD-Phe2, D-Trp3,6]-mGnRH was used. In late September, underyearling precocious male masu salmon that exhibited spermatogenesis were intraperitoneally injected with saline, sGnRH (0.1 µ g/g body weight (BW)), sGnRH (0.1 µ g/g BW) + GnRH antagonist (0.5 µ g/g BW), and GnRH antagonist (0.5 µ g/g BW), respectively. To measure plasma LH levels by RIA (Kobayashi et al. 1987), blood samples were collected at 3 h postinjection. GnRH antagonist treatment for 3 h significantly inhibited an increase in plasma LH levels that was artificially induced by exogenous sGnRH administration in underyearling precocious male masu salmon, indicating the stimulatory effects of sGnRH on LH secretion (Fig. 15). Considering the fact that sGnRH and not cGnRH-II has been detected in the pituitary of the salmonid fish (Amano et al. 1991), the effects of the GnRH antagonist on the LH levels are considered to be the result of the inhibition of sGnRH function. Subsequently, the effect of the GnRH antagonist on LH synthesis was examined in underyearling immature sockeye salmon that were pretreated with exogenous testosterone to increase the pituitary LH contents as a result of positive feedback (Okuzawa 2002). The GnRH antagonist (50 µ l/20 g BW, i.e., 1.0 µ g/g BW) was intraperitoneally injected once a week for 6 weeks from early doi:10.5047/absm.2010.00302.0039 © 2010 TERRAPUB, Tokyo. All rights reserved. M. Amano / Aqua-BioSci. Monogr. 3: 39–72, 2010 51 Fig. 15. Changes in plasma LH concentrations (ng/ml) in underyearling precocious male masu salmon (mean ± SEM). In each group (n = 8), means with different letters indicate significant difference (P < 0.05). Reprinted with permission of John Wiley & Sons, Inc. from Journal of Experimental Zoology Part A, 307A, Amano et al., Effects of a gonadotropin-releasing hormone antagonist on gonadotropin levels in masu salmon and sockeye salmon, 535–541, © 2007, Wiley-Liss, Inc., a Wiley Company. August. The fish were fed testosterone 25 µ g/g-diet (1.5% of BW) throughout the experiment. The LH contents in the pituitary and plasma were measured by RIA (Kobayashi et al. 1987). GnRH antagonist treatment slightly but significantly inhibited an increase in the testosterone-stimulated pituitary LH content levels (Fig. 16A). However, it had no effect on the plasma LH levels (Fig. 16B). Assuming that the pituitary LH content reflects the mRNA levels, as observed in striped bass Morone saxatilis (Hassin et al. 1999), these results indicate that the decrease in LH contents following GnRH antagonist treatment was due to a decrease in LH synthesis and not due to an increase in LH release. Thus, it is suggested that endogenous sGnRH stimulates LH synthesis in sockeye salmon. However, considering that the GnRH antagonist decreased the pituitary LH contents only slightly, the GnRH antagonist partially inhibited an increase in testosterone-stimulated LH synthesis (Amano et al. 2007). Moreover, GTH secretion is controlled not only by GnRH but also by other factors such as dopamine and sex steroids (Zohar et al. 2010). Thus, it is necessary to take these factors into consideration for more precise analysis. 5. Changes in GnRH levels in the brain during gonadal maturation 5-1. Changes in GnRH peptide levels during gonadal maturation in female masu salmon If GnRH is involved in gonadal maturation, GnRH contents in the brain would be expected to change during Fig. 16. Changes in (A) pituitary LH contents (ng) and (B) plasma LH concentrations (ng/ml) in underyearling immature sockeye salmon (mean ± SEM). **(P < 0.01) indicates statistical significance between the groups. In each group, means with different letters indicate significant difference (P < 0.05). Reprinted with permission of John Wiley & Sons, Inc. from Journal of Experimental Zoology Part A, 307A, Amano et al., Effects of a gonadotropin-releasing hormone antagonist on gonadotropin levels in masu salmon and sockeye salmon, 535–541, © 2007, Wiley-Liss, Inc., a Wiley Company. gonadal maturation. Several studies were conducted to measure brain GnRH contents by RIA in relation to gonadal maturation of teleost fish, e.g., in European eel (Dufour et al. 1982), platyfish Xiphophorus maculatus (Schreibman et al. 1983), brown trout Salmo trutta (Breton et al. 1986), caribe colorado Pygocentrus notatus (Gentile et al. 1986), goldfish (Yu et al. 1987), and chinook salmon Oncorhynchus tschawytscha (Lewis et al. 1992). However, the results were discordant. A clear correlation between brain GnRH contents and gonadal maturity was observed only in caribe colorado (Gentile et al. 1986). Such discrepancies may be, in part, due to the use of RIAs employing nonspecific antibodies. Thus, changes in sGnRH and cGnRH-II contents in the brain and pituitary of female masu salmon from hatching through gonadal maturation for three years were examined by using specific RIAs for sGnRH and cGnRH-II. For purposes of this study, masu salmon eggs were artificially fertilized in October. The eggs hatched in December, and the fish were reared under natural photoperiod in spring water of constant temperature (9–10°C) throughout the experiment. Brain and pituitary doi:10.5047/absm.2010.00302.0039 © 2010 TERRAPUB, Tokyo. All rights reserved. 52 M. Amano / Aqua-BioSci. Monogr. 3: 39–72, 2010 were sampled for the measurement of GnRHs and GTHs. Ovarian weights were measured to calculate gonadosomatic index (GSI) as follows: gonad weight/BW × 100. Extraction of GnRH from the brain tissue and measurements of sGnRH and cGnRH-II by respective RIAs were done according to Okuzawa et al. (1990). Changes in BW and GSI are shown in Figs. 17A and 17B. GSI rapidly increased from July through October of 2 year olds, in accordance with the advancement of vitellogenesis and ovulation. Ovulation was observed in October. sGnRH concentrations (pg/mg tissue) in the OB and the telencephalon including POA increased significantly during vitellogenesis and ovulation (Figs. 18A, B). sGnRH concentrations in the hypothalamus also showed a similar tendency (Fig. 18C). To the contrary, no significant changes were seen in the optic tectum-thalamus and cerebellum-medulla oblongata during vitellogenesis and ovulation (Figs. 18D, E). Pituitary sGnRH contents (pg) showed a stepwise increase every summer for three years, and significantly increased prior to ovulation (Fig. 19A). During gonadal maturation, LH levels in the pituitary and those in the plasma correlatively increased with the elevation in sGnRH levels in the brain and pituitary (Figs. 19B, C). cGnRH-II was undetectable in the pituitary throughout the experiment (data not shown). Further, no significant changes in the concentration of cGnRH-II were found in discrete brain areas during vitellogenesis and ovulation (data not shown). These results, together with those of immunohistochemical studies, suggest that sGnRH in the telencephalon including POA and the hypothalamus is involved in gonadal maturation in masu salmon (Amano et al. 1992). It has been demonstrated that GnRH fibers originating from the TN is not involved in GTH secretion and further in gonadal maturation in goldfish. The sGnRH contents in the brain except the OB markedly decreased by olfactory tract sectioning (OTX), whereas the cGnRH-II contents in the brain showed no clear changes. Despite large decreases in the brain sGnRH contents, gonadal maturation was not inhibited (Kobayashi et al. 1992, 1994; Kim et al. 2001). In dwarf gourami, sGnRH neurons in the TN may be the most extensively projecting GnRH neurons in the brain except in the pituitary (Oka and Matsushima 1993). These results indicate that most of the sGnRH fibers in the brain originates from the TN and that sGnRH neurons in the TN do not project to the pituitary. Thus, it is possible that changes in the sGnRH contents measured by RIA reflect the activities of GnRH neurons in the TN. It should be noted again that the levels of GnRH measured by RIA must be considered as a summation of synthesis, release, and degradation of GnRH at any point in development. Therefore, it is necessary to examine the expression of GnRH gene in order to clarify function of GnRH in reproduction. 5-2. Changes in sGnRH mRNA expression during gonadal maturation in female masu salmon Fig. 17. Changes in (A) BW (g) and (B) GSI (%) during gonadal maturation of female masu salmon (mean ± SEM). ***(P < 0.001) and *(P < 0.05) indicate statistical significance. Reprinted with permission from Zoological Science, 9, Amano et al., Changes in salmon GnRH and chicken GnRH-II contents in the brain and pituitary, and GTH contents in the pituitary in female masu salmon, 375–386, Fig. 4, © 1992, Zoological Society of Japan. In female masu salmon, sGnRH peptide levels in the telencephalon, including POA, and the hypothalamus increased during gonadal maturation as mentioned above (Amano et al. 1992). The widespread distribution of sGnRH-ir cell bodies in the ventral part of the brain suggests that sGnRH has several functions in the brain in addition to the stimulation of GTH secretion (Amano et al. 1991); however, it still remains obscure which sGnRH neurons are involved in gonadal maturation via GTH secretion. In situ hybridization (ISH) is useful to detect GnRH mRNA levels when certain GnRH neurons, such as sGnRH neurons in masu salmon, are scattered in the brain. Thus, the changes of sGnRH mRNA expression during ovulation in 2-year-old female masu salmon were examined by ISH. Sampling was carried out at 2-monthly intervals from April through October (month of ovulation). Brain tissue was fixed for ISH. Serial sagittal sections were cut at 8 µm. Procedure of ISH is illustrated in Fig. 20. The number of neurons expressing sGnRH mRNA in the OB, the TN, the VT, and the POA were measured (Amano et al. 1994). doi:10.5047/absm.2010.00302.0039 © 2010 TERRAPUB, Tokyo. All rights reserved. M. Amano / Aqua-BioSci. Monogr. 3: 39–72, 2010 53 Fig. 19. Changes in (A) pituitary sGnRH contents (pg), (B) pituitary LH contents (ng), and (C) plasma LH concentration (ng/ml) during gonadal maturation of female masu salmon (mean ± SEM). ***(P < 0.001) and *(P < 0.05) indicate statistical significance. (A) and (B) are reprinted with permission from Zoological Science, 9, Amano et al., Changes in salmon GnRH and chicken GnRH-II contents in the brain and pituitary, and GTH contents in the pituitary in female masu salmon, 375–386, Figs. 8 and 5A, respectively, © 1992, Zoological Society of Japan. Fig. 18. Changes in sGnRH concentrations (pg/mg tissue) in the (A) OB, (B) telencephalon including POA, (C) hypothalamus, (D) optic tectum-thalamus, and (E) cerebellum and medulla oblongata during gonadal maturation of female masu salmon (mean ± SEM). ***(P < 0.001), **(P < 0.01) and *(P < 0.05) indicate statistical significance. Reprinted with permission from Zoological Science, 9, Amano et al., Changes in salmon GnRH and chicken GnRH-II contents in the brain and pituitary, and GTH contents in the pituitary in female masu salmon, 375–386, Fig. 7, © 1992, Zoological Society of Japan. Changes in BW and GSI are shown in Figs. 21A and 21B, respectively. GSI rapidly increased from April through October, in accordance with vitellogenesis and ovulation. The number of neurons expressing sGnRH mRNA in the VT and the POA increased with gonadal maturation (Fig. 21C). Plasma LH levels remained low until August, but rapidly increased in October (Fig. 22A). These results indicate that sGnRH neurons in the VT and the POA are involved in the regulation doi:10.5047/absm.2010.00302.0039 © 2010 TERRAPUB, Tokyo. All rights reserved. 54 M. Amano / Aqua-BioSci. Monogr. 3: 39–72, 2010 Fig. 20. Procedure of in situ hybridization using 35S labeled oligonucleotide probe. of gonadal maturation possibly through GTH secretion. The plasma levels of estradiol-17β (E2) (Fig. 22B) and testosterone (Fig. 22C) increased in August, whereas 17α20β-dihydroxy-4-pregnen-3-one (DHP) (Fig. 22D) and LH levels suddenly increased in October. This suggests that increased E2 and/or testosterone stimulated sGnRH synthesis in the POA and VT as a result of positive feedback (Amano et al. 1995a). This is supported by the fact that sGnRH gene expression in the POA was activated by aromatizable androgen, 17 α-methyltestosterone (17α -MT) in future precocious male (Amano et al. 1994) and 2-year-old female masu salmon which are just before the initiation of gonadal maturation (Amano et al. 1997a). No significant changes of the sGnRH synthetic activity were observed in the OB and the TN (Fig. 21C), suggesting that sGnRH neurons in the OB and the TN are not involved in the regulation of gonadal maturation. In the dwarf gourami, GnRH originating from the TN does not affect GTH secretion but rather has a function as a neuromodulator in the brain (Oka and Ichikawa 1990; Oka 1992; Yamamoto et al. 1995). sGnRH originating from the TN is also not considered to be essential for gonadal development in goldfish (Kobayashi et al. 1992, 1994; Kim et al. 2001). It is also possible that sGnRH neurons in the TN function to modulate neuronal activity in masu salmon. sGnRH neurons in the ON and the OB are suggested to have some important roles in seaward migration of chum salmon (Kudo et al. 1994; Parhar et al. 1994). Wild masu salmon migrate to the sea in the spring after smoltification. The fish used were offspring of wild fish and they smoltified in the spring at 1.5 years old. Thus, it may be that sGnRH neurons in the OB are also involved in seaward migration in masu salmon. 5-3. Changes in brain GnRH mRNA and pituitary GnRH peptide levels during gonadal maturation in barfin flounder Barfin flounder first mature in early spring at 2 (male) and 3 years (female), respectively. The barfin flounder has sGnRH, cGnRH-II, and sbGnRH (Amano et al. 2002a). To clarify the possible functions of GnRH in the brain of barfin flounder, we have previously examined the distribution of the three forms of GnRH in various areas of the brain by RIA, and the localization of GnRH-ir cell bodies and fibers in the brain and pituitary by IHC. As a result, it appears that sbGnRH is involved in gonadal maturation through stimulating GTH secretion; sbGnRH-ir cell bodies located in the POA send fibers into the pituitary, and levels of sbGnRH in the pituitary were much higher than those of sGnRH and cGnRH-II (Amano et al. 2002b). To confirm that sbGnRH is involved in gonadal maturation in barfin flounder, it is necessary to show that sbGnRH levels in the brain and pituitary change during gonadal maturation. ISH is useful to detect GnRH mRNA levels when certain GnRH neurons, such as sGnRH neurons in masu salmon, are scattered in the brain. In the barfin flounder, GnRH neurons were localized in certain brain regions; sbGnRH, sGnRH, and cGnRH-II neurons were located in the POA, in the ventromedial part of the rostral OB and the TN, and in the MT, respectively (Amano et al. 2002a, b). In this case, it is useful to measure GnRH mRNA levels by real-time quantitative PCR rather than by ISH. Therefore, to clarify the physiological roles of the respective GnRH forms during gonadal maturation of the barfin flounder, changes in brain GnRH mRNA levels were examined by real-time quantitative PCR. We also measured GnRH peptide levels in the pituitary by time-resolved fluoroimmunoassay (TR-FIA) for sGnRH, cGnRH-II, and sbGnRH. We also examined the changes in plasma levels of testosterone, E2, and DHP. In male barfin flounder, GSI remained low until autumn of 1-year-olds and then rapidly increased in January and fish spermiated in March (Fig. 23A). The amount of sbGnRH mRNA per brain significantly increased in January and remained at high levels in March (Fig. 23B). The amounts of sGnRH and cGnRH-II mRNA per brain did not show significant changes during the experimental periods (Figs. 23C, D). Pituitary sbGnRH peptide doi:10.5047/absm.2010.00302.0039 © 2010 TERRAPUB, Tokyo. All rights reserved. M. Amano / Aqua-BioSci. Monogr. 3: 39–72, 2010 Fig. 21. Changes in (A) BW (g), (B) GSI (%), and (C) the number of neurons expressing sGnRH mRNA in the OB, TN, VT and POA during gonadal maturation of 2-year-old female masu salmon (mean ± SEM). In each group, means with different letters indicate significant difference (P < 0.05). (B) and (C) are reprinted from General and Comparative Endocrinology, 99, Amano et al., Salmon GnRH synthesis in the preoptic area and the ventral telencephalon is activated during gonadal maturation in female masu salmon, 13–21, © 1995, Academic Press, Inc., with permission from Elsevier. content significantly increased in March (Fig. 23E). Pituitary sGnRH peptide and cGnRH-II peptide contents were extremely low compared to sbGnRH peptide levels and showed no significant changes during the experiment (data not shown). In female barfin flounder, GSI remained low until August of 2-year-olds and increased thereafter until April when the fish began to ovulate (Fig. 24A). The sbGnRH mRNA levels per brain increased significantly from August to April (Fig. 24B). Pituitary sbGnRH peptide levels also increased significantly during this period (Fig. 24C). These results in 55 Fig. 22. Changes in plasma concentrations (ng/ml) of (A) LH, (B) E2, (C) testosterone, and (D) DHP during gonadal maturation of 2-year-old female masu salmon (mean ± SEM). In each group, means with different letters indicate significant difference (P < 0.05). Reprinted from General and Comparative Endocrinology, 99, Amano et al., Salmon GnRH synthesis in the preoptic area and the ventral telencephalon is activated during gonadal maturation in female masu salmon, 13–21, © 1995, Academic Press, Inc., with permission from Elsevier. both sexes indicate that sbGnRH is involved in gonadal maturation in barfin flounder via the synthesis of steroid hormones (Amano et al. 2004a, 2008). doi:10.5047/absm.2010.00302.0039 © 2010 TERRAPUB, Tokyo. All rights reserved. 56 M. Amano / Aqua-BioSci. Monogr. 3: 39–72, 2010 Fig. 24. Changes in (A) GSI (%), (B) sbGnRH mRNA levels (× 106 copies/brain), and (C) pituitary sbGnRH contents (ng/pituitary) during gonadal maturation of female barfin flounder (mean ± SEM). Means with differing letters differ significantly (P < 0.05). Reprinted from General and Comparative Endocrinology, 158, Amano et al., Changes in brain seabream GnRH mRNA and pituitary seabream GnRH peptide levels during ovarian maturation in female barfin flounder, 168–172, © 2008, Elsevier Inc., with permission from Elsevier. Fig. 23. Changes in (A) GSI (%), (B) sbGnRH mRNA levels (× 107 copies/brain), (C) sGnRH mRNA levels (× 107 copies/brain), (D) cGnRH-II mRNA levels (× 107 copies/brain), and (E) pituitary sbGnRH contents (ng) during gonadal maturation of male barfin flounder (mean ± SEM). Means with differing letters differ significantly (P < 0.05). Reprinted from Comparative Biochemistry and Physiology, Part B, 138, Amano et al., Changes in brain GnRH mRNA and pituitary GnRH peptide during testicular maturation in barfin flounder, 435–443, © 2004, Elsevier Inc., with permission from Elsevier. 6. Effects of photoperiod on the brain–pituitary– gonadal axis in masu salmon and sockeye salmon 6-1. Stimulation of precocious maturation in male masu salmon by short photoperiod treatment It is well established that photoperiod is of primary importance in the induction of the initiation and modulation of reproductive development in salmonid fish. For example, in Atlantic salmon parr, precocious male reared under short photoperiod mature earlier than those reared under long photoperiod (Lundqvist 1980). In landlocked masu salmon “yamame”, rapid vitellogenesis is induced by a short photoperiod, although maturation is initiated doi:10.5047/absm.2010.00302.0039 © 2010 TERRAPUB, Tokyo. All rights reserved. M. Amano / Aqua-BioSci. Monogr. 3: 39–72, 2010 under a long photoperiod (Takashima and Yamada 1984). However, the endocrinological mechanisms of such phenomena were not fully understood. In masu salmon reared under constant water temperature, sGnRH concentrations in the discrete brain areas showed seasonal changes: high during autumn–winter and low in summer. In addition, pituitary sGnRH contents showed a stepwise increase every summer for three years (Amano et al. 1992). These results suggest that synthesis of sGnRH in the brain increases under a short photoperiod and sGnRH produced is transported from cell bodies to the pituitary to induce gonadal maturation. However, direct evidence is lacking. Therefore, the effects of the photoperiod on sGnRH mRNA levels in the brain, GTH levels in the pituitary, and testicular maturation of 1-year-old male masu salmon were examined in order to ascertain that short photoperiod induces increases in sGnRH and GTH. Underyearling male masu salmon were reared under a short photoperiod (8L16D; lights on 0900–1700 h) and a long photoperiod (16L8D; lights on 0400–2000 h) from June through September. Fish were sampled in July, August, and October. Changes in GSI are shown in Fig. 25A. Spermiation was observed in August in the 8L16D group and in September in the 16L8D group, respectively. sGnRH mRNA levels in the VT and the POA increased when the fish spermiated; the activity increased in August in the 8L16D group, and in September in the 16L8D group, respectively (Figs. 25B, C). Moreover, the increase of sGnRH mRNA levels was in accordance with the increase of pituitary FSHβ (Fig. 25D) and LHβ contents (Fig. 25E). No significant changes in sGnRH mRNA levels in relation to gonadal maturation were observed in the OB and the TN (data not shown). These results indicate that sGnRH neurons in the VT and the POA are influenced by photoperiod, and are involved in the regulation of gonadal maturation through GTH secretion (Amano et al. 1995b). It has been shown that sGnRH synthetic activity is also considered to be influenced by gonadal steroids; sGnRH mRNA levels in the POA were activated by oral 17α -MT treatment in 1-year-old future precocious males (Amano et al. 1994) and in 2-year-old female masu salmon, which initiated ovarian development (Amano et al. 1997a). These results suggest that sGnRH neurons in the VT and the POA also respond to changes in steroid hormone levels. From these results, it is suggested that sGnRH synthetic activity is influenced by both photoperiod and steroid hormones. To further assess the effects of photoperiod on sGnRH synthetic activity, underyearling precocious male masu salmon were castrated to remove the effects of steroid hormones, and the changes in sGnRH mRNA levels in the brain were examined by manipulating the photoperiod. In early August, underyearling male masu salmon (mean BW 9.4 g; mean GSI 3.35%) were castrated 57 according to methods reported (Aida et al. 1984). Upon recovery, the fish were divided into short photoperiod (8L16D; lights on 0900–1700 h) and long photoperiod (16L8D; lights on 0400–2000 h) groups. The fish were sampled 30 days after the castration. Blood was collected to measure plasma testosterone levels by a TR-FIA (Yamada et al. 1997). For ISH, brains were fixed with 4% PFA and 1% picric acid in 50 mM PB (pH 7.3). Plasma testosterone concentrations decreased compared to initial levels (1.3 ng/ml) and no significant differences were seen between the groups (Fig. 26A), indicating the validity of castration. The number of neurons expressing sGnRH mRNA in the POA was greater in the 8L16D group than the 16L8D group (Fig. 26B). No significant differences were observed in the VT (Fig. 26C). These results indicate that sGnRH neurons in the POA receive short photoperiodic signals and that either short photoperiod or steroid hormone secretion is required for the activation of sGnRH synthesis in 1-year-old precocious male masu salmon (Amano et al. 1999). 6-2. Incomplete development of the brain–pituitary–gonadal axis in male sockeye salmon It is well known that precocious maturation occurs in male salmonid fishes including Atlantic salmon, amago salmon Onvorhynchus rhodurus, masu salmon, and sockeye salmon. However, the age at which fish mature precociously differs according to species. For example, most male masu salmon precociously mature in the first autumn, while precocious males first appear in the second summer in sockeye salmon landlocked in Japanese lakes. Although the cause of this species-specific difference in the initiation of precocious maturation is of much interest, fundamental mechanisms have not yet been elucidated. Since gonadal maturation of teleost fish is regulated by brain–pituitary–gonadal axis, there is a possibility that the periodic difference in the appearance of precocious maturation between masu salmon and sockeye salmon is due to differences in the development of the brain–pituitary–gonadal axis. Indeed, precocious maturation of underyearling male masu salmon can be induced by short photoperiodic treatment (Amano et al. 1995b). Thus, effects of photoperiod on brain–pituitary–gonadal axis of underyearling sockeye salmon were examined. In June, underyearling sockeye salmon (mean BW 8.0 g) were divided into the short photoperiod (8L16D; lights on 0900–1700 h) group and the long photoperiod (16L8D; lights on 0400–2000 h) group. Fish were reared until October. No significant differences in GSI were observed between both groups throughout the experiment (Fig. 27A). Fish were still immature in October. sGnRH contents in the telencephalon including POA tended to be higher in the 8L16D group than in the 16L8D group from late August (Fig. 27B). sGnRH contents in the hypothalamus doi:10.5047/absm.2010.00302.0039 © 2010 TERRAPUB, Tokyo. All rights reserved. 58 M. Amano / Aqua-BioSci. Monogr. 3: 39–72, 2010 Fig. 25. Changes in (A) GSI (%), (B) the number of neurons expressing sGnRH mRNA in the POA, (C) the number of neurons expressing sGnRH mRNA in the VT, (D) pituitary FSH β contents (ng/pituitary), and (E) pituitary LHβ contents (ng/pituitary) during gonadal maturation of underyearling male masu salmon reared under short and long photoperiods (mean ± SEM). In each group, means with differing letters differ significantly (P < 0.05). ***(P < 0.001), **(P < 0.01) and *(P < 0.05) indicate statistical significance between the groups. (A)–(C) are reprinted from General and Comparative Endocrinology, 99, Amano et al., Short photoperiod accelerates preoptic and ventral telencephalic salmon GnRH synthesis and precocious maturation in underyearling male masu salmon, 22–27, © 1995, Academic Press, Inc., with permission from Elsevier. were significantly higher in the 8L16D group in September (Fig. 27C). sGnRH contents in the pituitary were significantly higher in the 8L16D group in September and October (Fig. 27D). Pituitary FSHβ contents were significantly higher in the 8L16D group in early and late August and October (Fig. 27E). In the 8L16D group, a positive correlation was observed between GSI and pituitary FSH β content (correlation coefficient r = 0.8250, P < 0.001, n = 34). LHβ was undetectable in the pituitary in all individuals. These results suggest that synthesis and storage of sGnRH increased under the short photoperiod and then sGnRH stimulates FSHβ synthesis. It is reported that FSH is involved in the early phases of spermatogenesis in salmonid fishes (Kawauchi et al. 1989; Swanson et al. 1989). Indeed, FSHβ was positively correlated with GSI in the short photoperiod group. However, levels of doi:10.5047/absm.2010.00302.0039 © 2010 TERRAPUB, Tokyo. All rights reserved. M. Amano / Aqua-BioSci. Monogr. 3: 39–72, 2010 Fig. 26. (A) Plasma testosterone (ng/ml), (B) the number of neurons expressing sGnRH mRNA in the POA, and (C) the number of neurons expressing sGnRH mRNA in the VT of castrated precocious male masu salmon reared under short and long photoperiods (mean ± SEM). **(P < 0.01) indicates statistical significance. Reprinted from General and Comparative Endocrinology, 115, Amano et al., Effects of photoperiod on salmon GnRH mRNA levels in brain of castrated underyearling precocious male masu salmon, 70–75, © 1999, Academic Press, with permission from Elsevier. pituitary LHβ were not increased. These results suggest that synthesis and release of LH are required for gonadal maturation. Furthermore, it is suggested that the development of the brain–pituitary–gonadal axis of underyearling sockeye salmon is inactive. This species-specific difference in the development of the brain–pituitary–gonadal axis may underlie differences in the initiation of precocious maturation (Amano et al. 1997b). 7. Steroid feedback on GnRH levels in the brain of masu salmon and sockeye salmon 7-1. Effects of steroid hormone administration on GnRH synthesis in masu salmon Accumulation of GTH in the pituitary can be stimulated by aromatizable androgen or estrogen in juvenile fish; this is a known positive feedback system (see Goos 1987; Okuzawa 2002). The involvement of GnRH in this 59 mechanism is speculated because sex steroid administration increased the amount of GnRH in the hypothalamus of the European silver eel (Dufour et al. 1985) and rainbow trout (Goos et al. 1986). On the other hand, the stimulatory effects of testosterone on the expression of pituitary LHβ gene were demonstrated in vitro in juvenile rainbow trout (Xiong et al. 1993, 1994a, b). Thus, whether steroids have direct actions on the pituitary, or act indirectly via GnRH was unclear. Therefore, the effects of 17α-MT on sGnRH mRNA expression in 1-year-old masu salmon (mean BW 44 g) were examined. Fish were fed pellets containing 17αMT 25 µg/g-diet (1.5% of the BW) for 41 days. The fish used in this study consisted of future precocious males which will mature in autumn of that year and of immature females. Blood was sampled for determination of plasma LHβ levels. Pituitary contents of FSHβ and LHβ, and plasma LHβ level were measured by respective RIAs as described in Section 4-1. Pituitary sGnRH was extracted and measured by RIA (Okuzawa et al. 1990). Brains were fixed for ISH. Oral 17α-MT application markedly increased pituitary LHβ, but not FSHβ concentrations in both sexes (Figs. 28A, B). In future precocious males, 17α -MT treatment further increased the number of neurons expressing sGnRH mRNA in the POA (Fig. 28C) but not in the OB and the VT (Figs. 28D, E). However, sGnRH mRNA levels were not changed by 17α -MT in immature females (Figs. 28C, D, E). These results suggest that sGnRH is involved in the positive feedback system at least in future precocious males, and that the difference in the responsiveness of preoptic sGnRH neurons to 17 α-MT is based on the maturational stage of the fish (Amano et al. 1994). Thus, whether sGnRH mRNA levels were influenced by 17α-MT in 2-year-old females which are just before the initiation of gonadal maturation was further examined. 1-Year-old and 2-year-old masu salmon were randomly selected from a stock in May. At the start of the experiment, mean BW was 29 and 94 g, and mean GSI was 0.33 and 0.53%, for 1-year-old and 2-year-old fish, respectively. Fish were divided into the control group and 17 α-MT 25 µg/g-diet treated group. The 17 α-MTtreated group was fed pellets containing 17α-MT (1.5% of the BW) for 31 days. Brains were fixed for ISH. The number of neurons expressing sGnRH mRNA in the POA increased by 17α -MT in 2-year-old female masu salmon, not in 1-year-old female masu salmon (Fig. 29A). No significant difference was seen in the number of neurons expressing sGnRH mRNA in the VT by 17 α-MT treatment in both ages (Fig. 29B). These results support the previous hypothesis that the difference in the responsiveness of preoptic sGnRH neurons to 17α -MT is based on the maturational stage of the fish (Amano et al. 1997a). Of note, studies on pubertal development have been conducted in a number of teleost fish species such as the rainbow trout (Gielen et al. 1982; doi:10.5047/absm.2010.00302.0039 © 2010 TERRAPUB, Tokyo. All rights reserved. 60 M. Amano / Aqua-BioSci. Monogr. 3: 39–72, 2010 Fig. 27. Changes in (A) GSI (%), (B) sGnRH contents (pg) in the telencephalon including POA, (C) sGnRH contents (pg) in the hypothalamus, (D) sGnRH contents (pg) in the pituitary, and (E) pituitary FSHβ contents (ng) during gonadal maturation of male sockeye salmon reared under short and long photoperiods (mean ± SEM). In each group, means with differing letters differ significantly (P < 0.05). **(P < 0.01) and *(P < 0.05) indicate statistical significance between the groups. Reprinted with permission from Fisheries Science, 63, Amano et al., Incomplete development of the brain–pituitary–gonadal axis may underlie the delay in the initiation of precocious maturation in male sockeye salmon, 873–876, Figs. 2–4, © 1997, the Japanesel Society of Fisheries Science. Goos et al. 1986), the eel (Dufour et al. 1985; Counis et al. 1987), the platyfish (Schreibman et al. 1986), and African catfish (Schulz et al. 1994a, b). These studies suggest that sex steroids initiate and/or accelerate the development of the brain–pituitary–gonadal axis. Taken together, the response of preoptic sGnRH neurons to steroids may be a necessary condition for gonadal maturation or the development of the brain–pituitary–gonadal axis (Amano et al. 1997a). 7-2. Low GnRH levels in the brain and pituitary in triploid female sockeye salmon Triploid female fish are known to be sterile. One possible explanation is that oocytes cannot undergo normal meiotic division to produce euploid gamates. Another is that triploid fish have a defective reproductive endocrine system, which regulates gonadal development. It is well established that positive feedback system of sex doi:10.5047/absm.2010.00302.0039 © 2010 TERRAPUB, Tokyo. All rights reserved. M. Amano / Aqua-BioSci. Monogr. 3: 39–72, 2010 61 Fig. 29. Changes in the number of neurons expressing sGnRH mRNA in (A) the POA and (B) VT of 1-year-old and 2-year-old female masu salmon of control and 17α-MT-treated groups (mean ± SEM). *(P < 0.05) indicates statistical significance. With kind permission from Springer Science+Business Media: Fish Physiology and Biochemistry, The maturation of the salmon GnRH system and its regulation by gonadal steroids in masu salmon, 17, 1997, 63–70, Amano et al., Fig. 2, © 1997, Kluwer Academic Publishers. Fig. 28. (A) Pituitary FSHβ contents (ng), (B) pituitary LHβ contents (ng), the number of neurons expressing sGnRH mRNA in (C) the POA, (D) VT and (E) OB of masu salmon of control and 17α-MT-treated groups (mean ± SEM). ***(P < 0.001) and **(P < 0.01) indicate statistical significance. Reprinted from General and Comparative Endocrinology, 95, Amano et al., Activation of salmon gonadotropin-releasing hormone synthesis by 17α-methyltestosterone administration in yearling masu salmon, Oncorhynchus masou, 374–380, © 1994, Academic Press, with permission from Elsevier. steroids exists in the brain–pituitary–gonadal system in juvenile fish (Okuzawa 2002). There are some studies on plasma levels of steroid hormones in triploid salmonids and it has been clarified that triploid fish have low levels of steroid hormones. Triploid females are known to have much lower E2 levels than those of maturing diploid of the same age in rainbow trout (Lincoln and Scott 1984; Nakamura et al. 1987; Benfey et al. 1989a), pink salmon Oncorhynchus gorbuscha (Benfey et al. 1989a), coho salmon (Benfey et al. 1989b), and ayu Plecoglossus altivelis (Iguchi et al. 1991). Thus, the triploid female fish is considered to be a good model for examining the feedback system in the context of low steroid hormone levels. No information is available on brain and pituitary GnRH contents of triploid salmonid fish, whereas it has been reported that triploid salmonids have low plasma LH concentration and low pituitary LH content (Benfey et al. 1989b). Therefore, levels of sGnRH in the brain and pituitary of 1-year-old diploid and triploid female sockeye salmon were compared. Eggs of sockeye salmon were artificially fertilized in September, and triploidy was induced by heat shock treatment (30°C, 10 min). Diploid fish were also obtained from artificially fertilized eggs of the same stock used for triploids. Fish were reared under natural photoperiod using natural spring water at constant temperature (9–10°C). Sampling was conducted at 22 months later. doi:10.5047/absm.2010.00302.0039 © 2010 TERRAPUB, Tokyo. All rights reserved. 62 M. Amano / Aqua-BioSci. Monogr. 3: 39–72, 2010 Fig. 30. Levels of (A) GSI (%), (B) plasma testosterone (ng/ml), (C) plasma LH (ng/ml), and (D) pituitary LH content (ng) of diploid and triploid sockeye salmon (mean ± SEM). **(P < 0.01) and *(P < 0.05) indicate statistical significance. Data was derived with permission from Fisheries Science, 64, Amano et al., Low GnRH levels in the brain and the pituitary in triploid female sockeye salmon, 340–341, Table 1, © 1998, the Japanesel Society of Fisheries Science. Triploid fish (mean BW 76 g) had significantly lower GSI than did diploid fish (mean BW 63 g) (Fig. 30A). Diploids had oocytes in the early and late perinucleolus stage, and in some fish, oocytes were in the oil droplet stage, whereas triploids had only oogonia and lacked follicle cells (data not shown). Plasma levels of E2 were below detectable limit (0.03 ng/ml) in both diploid and triploid fish. Plasma levels of testosterone were significantly lower in triploids (Fig. 30B). Plasma LH concentration of diploid fish was 2.24 ng/ml, whereas this was below the detectable limit (0.88 ng/ml) in triploid fish (Fig. 30C). Although no significant differences in pituitary LH contents were observed, they tended to be lower in triploid fish (Fig. 30D). Triploid had significantly lower sGnRH concentrations in the telencephalon including POA, hypothalamus, and pituitary, whereas no significant differences of sGnRH concentration were seen in the other parts of the brain (Fig. 31). These results suggest that the low sGnRH concentrations cause low plasma LH and steroid hormone levels in triploid fish (Amano et al. 1998a). The reason for the low brain sGnRH concentrations is not known; it is not likely to be due to active release of sGnRH in the pituitary but to be due to low sGnRH synthesis in the brain, since plasma LH levels were lower in triploid fish. Low testosterone levels caused by low sGnRH and LH production or by incomplete development of ovarian follicular cells may not stimulate further production of sGnRH in the triploid fish brain. 8. Ontogenic development of GnRH neurons 8-1. Ontogenic development of GnRH neurons in masu salmon At present, three GnRH neuronal systems are proposed in teleost fish judging from the location of GnRH neuronal somata and their projections (Oka 1997); the TN-GnRH system, the MT-GnRH system, and Fig. 31. Levels of sGnRH concentrations (pg/mg tissue) in diploid and triploid sockeye salmon (mean ± SEM). The letters a–g represent the following brain areas: a, olfactory bulb; b, telencephalon including preoptic area; c, hypothalamus; d, optic tectum-thalamus including midbrain; e, cerebellum; f, medulla oblongata; g, pituitary. ***(P < 0.001) and *(P < 0.05) indicate statistical significance. Reprinted with permission from Fisheries Science, 64, Amano et al., Low GnRH levels in the brain and the pituitary in triploid female sockeye salmon, 340–341, Fig. 1A, © 1998, the Japanese Society of Fisheries Science. the POA-GnRH system (in some species POA-GnRH neurons do not form a well-defined cell cluster and, in this monograph, the author defines the ventral forebrainGnRH system excluding the TN-GnRH system as the POA-GnRH system). The MT-GnRH system is the most conserved among the vertebrate species; it produces cGnRH-II and is present in all the teleost fish examined to date. Among the three GnRH systems, the POA-GnRH system is considered to regulate GTH secretion because it is the main system that projects directly to the pituitary (Oka 1997). Perciform fish have three forms of GnRH, sGnRH, cGnRH-II and sbGnRH. With very few exceptions such as European sea bass Dicentrarchus labrax doi:10.5047/absm.2010.00302.0039 © 2010 TERRAPUB, Tokyo. All rights reserved. M. Amano / Aqua-BioSci. Monogr. 3: 39–72, 2010 (González-Martínez et al. 2001), the TN-GnRH system and the POA-GnRH system are clearly distinguishable and the GnRH forms produced in these two systems are different; sGnRH is produced in the TN and sbGnRH in the POA. The medaka Oryzias latipes has sGnRH, cGnRH-II and medaka GnRH (mdGnRH), expressed in the TN, the MT, and the POA, respectively (Okubo et al. 2000). In contrast, in the other teleost species that have two forms of GnRH in the brain (e.g., salmonid fishes, eel, catfish), clear anatomical identification of the TN-GnRH system and POA-GnRH system is difficult, because the GnRH neurons located in the ventral forebrain are consecutive and the GnRH form(s) produced in these neurons are the same; sGnRH in salmonid fishes, mGnRH in the eel and cfGnRH in the catfish (Okuzawa and Kobayashi 1999). Studies of ontogeny of different GnRH systems are considered to be helpful in understanding the functional role of each GnRH system. The ontogeny of the GnRH system was first studied by IHC in mammalian (Schwanzel-Fukuda and Pfaff 1989; Wray et al. 1989). It was later reported in amphibian (Muske and Moore 1990; Murakami et al. 1992) and avian brains (Murakami et al. 1991). Results indicate that GnRH neurons are derived from the olfactory placode (OP) and migrate into the forebrain during prenatal development. In teleost fish, it has been reported by IHC in chum salmon (Chiba et al. 1994) and sockeye salmon (Parhar et al. 1995) that GnRH (probably sGnRH) neurons originate from the OP and then migrate into the brain along the ON. However, little is known about the differential ontogeny of sGnRH and cGnRH-II systems in the brain of salmonid fish. Thus, the ontogenic development of sGnRH and cGnRHII systems in the brain of masu salmon was examined by IHC and ISH. Masu salmon eggs were artificially fertilized and were reared under natural photoperiod in spring water of constant temperature (9–10°C). Fish were sampled at embryonic stage, alevin stage (still have yolk sacks and stay at the bottom of the tank), and fry stage (initiate feeding and swimming) (Kubo 1980). The procedure for IHC and ISH were basically similar to that described in Amano et al. (1991, 1994). Distribution of sGnRH neurons in the brain is summarized in Fig. 32. sGnRH-ir fibers were initially detected by IHC in the vicinity of the OP of the embryo 36 days after fertilization (Day 36) (Figs. 33A, B), and were distributed widely in the brain as well as the pituitary of alevin just after hatching (Day 80) (Fig. 33C). sGnRH-ir cell bodies were first detected about six months after fertilization in the rostroventral brain area ranging from the ON to the POA. sGnRH neuronal somata were detected earlier by ISH than IHC. Neuronal somata expressing sGnRH mRNA were first detected in the vicinity of the olfactory epithelium (OE) in embryo on Day 40 (Fig. 33D), and then were seen to be migrating from 63 Fig. 32. Microprojection drawing of a series of sagittal sections through whole heads of masu salmon on days (A) 40, (B) 60, (C) 80, and (D) 152 showing the distribution of neuronal somata expressing sGnRH (solid circle). F forebrain, OB olfactory bulb, OE olfactory epithelium, ON olfactory nerve, POA preoptic area, T telencephalon, VT ventral telencephalon. Bar indicates 0.5 mm. With kind permission from Springer Science+Business Media: Cell and Tissue Research, Ontogenic development of salmon GnRH and chicken GnRH-II systems in the brain of masu salmon, 293, 1998, 427–434, Amano et al., Fig. 4, © 1998, Springer-Verlag. the OE, along the ON of alevin (Day 60) (Fig. 33E), and in the transitional areas between the ON and the OB of alevin (Day 80) (Fig. 33F). In the brain, these neurons were first detected in the ventral OB on Day 80, and thereafter they were detected also in the caudal brain regions (data not shown). cGnRH-II system was detected later than sGnRH system; cGnRH-II-ir fibers were first detected of alevin (Day 67) (data not shown). cGnRH-II-ir neuronal somata were not detected during the present observation period. These results suggest that sGnRH neurons derive from the OP and then migrate into the brain, and that sGnRH is synthesized first and cGnRH-II later (Amano et al. 1998b). Among sGnRH neurons distributed from the ON through the POA, those in the VT and the POA are indicated to regulate GTH secretion. It is of interest whether all the sGnRH neurons originate from the OE or not. Thus, whether sGnRH neurons are present in the doi:10.5047/absm.2010.00302.0039 © 2010 TERRAPUB, Tokyo. All rights reserved. 64 M. Amano / Aqua-BioSci. Monogr. 3: 39–72, 2010 Fig. 33. (A) Sagittal section through the OP of masu salmon embryo on Day 36. (B) Higher magnification of “A”. sGnRH-ir fibers (arrowheads) are observed. (C) Sagittal section through the pituitary of masu salmon just after hatching (Day 80). Arrowhead indicates sGnRH-ir fiber. (D) Sagittal section of masu salmon embryo on Day 40 showing neuronal somata expressing sGnRH mRNA in the vicinity of the OE. This figure corresponds to Fig. 32A. (E) Sagittal section through a fish at Day 60 showing neuronal somata expressing sGnRH mRNA as a few small clusters, emerging from the OE, along the ON. This figure corresponds to Fig. 32B. (F) Sagittal section through the ON of masu salmon on Day 80 showing neuronal somata expressing sGnRH mRNA at transitional area between ON and OB. This figure corresponds to Fig. 32C. Bar indicates 20 µm. With kind permission from Springer Science+Business Media: Cell and Tissue Research, Ontogenic development of salmon GnRH and chicken GnRH-II systems in the brain of masu salmon, 293, 1998, 427–434, Amano et al., Figs. 2, 3, © 1998, Springer-Verlag. doi:10.5047/absm.2010.00302.0039 © 2010 TERRAPUB, Tokyo. All rights reserved. M. Amano / Aqua-BioSci. Monogr. 3: 39–72, 2010 VT-POA of fish whose OE including sGnRH clusters were cauterized just after hatching, was examined by ISH. Just hatched larvae (Day 44) were collected and were anesthetized. For the olfactory epithelium lesioned (OEL) group, both nares including sGnRH clusters were cauterized using an extra fine hot needle attached to a handy type soldering iron for about 2 seconds under a binocular microscope. Then, larvae were revived under aerated running water. For the control group, fish were also anesthetized and were put on the desk for about 30 seconds and then revived in aerated running water. Fish were sampled 7 months after the operation. Neurons expressing sGnRH mRNA were detected in the ON, ventral OB and TN in all fish of the control group (9 fish). In the OEL group (12 fish), however, a few neurons expressing sGnRH mRNA were detected in these regions in 3 fish. The rate of appearance of neurons expressing sGnRH mRNA in the ON-TN was significantly lower in the OEL group than the control group. Neurons expressing sGnRH mRNA were detected in the VT and the POA in 5 fish of the control group. However, neurons expressing sGnRH mRNA were not detected in these regions in any of the OEL group fish. The rate of appearance of neurons expressing sGnRH mRNA in the VT-POA was significantly lower in the OEL group than the control group. Pituitary sGnRH content in the OEL group was just above the detectable limit (1.85 pg) and was significantly lower than the corresponding control value both in males and in females (Fig. 34). These results indicate that sGnRH neurons in the VT-POA are derived from the OE in masu salmon, although the possibility cannot be ruled out that sGnRH neurons in the VT-POA arise from the VT-POA but were delayed in expressing sGnRH because of the trauma of cauterization (Amano et al. 2002c). 8-2. Ontogenic development of GnRH neurons in barfin flounder The barfin flounder experiences a metamorphosis early in development. Barfin flounder expresses sGnRH, cGnRH-II, and sbGnRH (Amano et al. 2002a). In the barfin flounder, the TN-GnRH system, the MT-GnRH system and the POA-GnRH system are clearly distinguishable, as in perciform fish and medaka. sbGnRH is physiologically the most important hypophysiotropic factor for the reproduction of the barfin flounder (Amano et al. 2002a, b, 2004a, 2008). To elucidate the ontogenic origin of the neurons that produce these GnRH molecules, the development of three GnRH systems was examined by IHC and ISH. Barfin flounder larvae used are shown in Fig. 35. Distribution of GnRH neurons in the brain is summarized in Fig. 36. cGnRH-II mRNA-expressing neuronal somata were first identified in the MT near the ventricle on Day 7 65 Fig. 34. Pituitary sGnRH content (pg) in male and female masu salmon of the control and OEL groups. ***(P < 0.001) and **(P < 0.01) indicate the level of statistical difference between the two groups. Reprinted from General and Comparative Endocrinology, 127, Amano et al., Ontogenic origin of salmon GnRH neurons in the ventral telencephalon and the preoptic area in masu salmon, 256–262, © 2002, Elsevier Science (USA), with permission from Elsevier. (Fig. 37A). cGnRH-II-ir fibers were first found in the brain on Day 7 (data not shown). Neuronal somata that express sGnRH mRNA were detected first in the vicinity of the OE on Day 21 (Fig. 37B), and then in the transitional area between the ON and OB (Fig. 37C) and the TN on Day 28 (Fig. 37D). sbGnRH mRNA-expressing neuronal somata were first detected in the POA on Day 42 (Fig. 37E). sbGnRH-ir fibers were localized in the POA-hypothalamus and formed a distinctive bundle of axons projecting to the pituitary on Day 70 (data not shown). These results indicate that three forms of GnRH neurons have separate embryonic origins in the barfin flounder as in other perciform fish such as tilapia Oreochromis niloticus and red seabream (Amano et al. 2004b). In contrast, it has been reported that sbGnRH neurons derive from the olfactory placode and migrate into the brain in African cichlid (White and Fernald 1998) and European sea bass (González-Martínez et al. 2002). Although the possibility cannot be ruled out that sbGnRH neurons migrate without expressing sbGnRH in the barfin flounder, the origin of sbGnRH neurons (POA-GnRH system) may differ according to the fish species. 9. Conclusion and future direc tions Currently, 15 forms of GnRH molecules have been characterized throughout the vertebrates and the existence of multiple forms of GnRH in the brain was also observed in teleost fish. A salmonid fish, masu salmon, has sGnRH and cGnRH-II, and a pleuronectiform fish, barfin flounder, has sGnRH, cGnRH-II and sbGnRH. In masu salmon, sGnRH neurons are scattered from the ON and the TN through the VT doi:10.5047/absm.2010.00302.0039 © 2010 TERRAPUB, Tokyo. All rights reserved. 66 M. Amano / Aqua-BioSci. Monogr. 3: 39–72, 2010 Fig. 35. Barfin flounder larvae of (A) 7 days after hatching (yolk-sack larva), (B) 21 days after hatching (flexion larva), (C) 28 days after hatching (postflexion larva, onset of metamorphosis), and (D) 42 days after hatching (postflexion larva, late metamorphosis). Bars indicate 5 mm. and the POA. sGnRH neurons in the VT and the POA are considered to be involved in gonadal maturation via stimulation of GTH secretion. The other sGnRH neurons and cGnRH-II neurons are suggested to have a neuromodulatory function in the brain. In barfin flounder, sbGnRH neurons, which are located in the POA, are considered to be involved in gonadal maturation, and sGnRH and cGnRH-II are suggested to function as a neuromodulator in the brain. It has recently been reported that cGnRH-II modifies food intake in the musk shrew Suncus murinus (Kauffman and Rissman 2004); cGnRH-II administration in the brain reduced 24-h ad libitum food intake. Supposing that melaninconcentrating hormone (MCH) also stimulates food intake in the musk shrew, it is reasonable to speculate that MCH and cGnRH-II interact in the brain and regulate food intake. Indeed, in the barfin flounder, it is indicated that cGnRH-II stimulates food intake by interacting with MCH; food intake regulation might be a novel function of cGnRH-II in teleost fish (Amiya et al. 2008). Furthermore, interaction between GnRH and other neuropeptide (e.g., neuropeptide Y, orexin) should be examined in detail. It has been clarified that three GnRH systems exist in the barfin flounder, judging from the location of cell bodies and their projections; the TN-, the MT-, and the POA-GnRH systems. However, in masu salmon, clear anatomical identification of the TN- and the POA-GnRH system is difficult because the GnRH neurons located in the ventral forebrain are consecutive and the GnRH form produced in these neurons is the same (sGnRH). Thus, it is suggested in masu salmon that sGnRH neurons are derived from the OE, migrate into the brain, and play different roles according to the location in the brain. Very recently, the KiSS1/GPR54 system was discovered in mammals (Ohtaki et al. 2001; de Roux et al. 2003; Seminara et al. 2003). It is now confirmed that KiSS1/GPR54 signaling is central to the regulation of GnRH and, consequently, FSH and LH secretion (Zohar et al. 2010). In teleost fish, the identification and doi:10.5047/absm.2010.00302.0039 © 2010 TERRAPUB, Tokyo. All rights reserved. M. Amano / Aqua-BioSci. Monogr. 3: 39–72, 2010 Fig. 36. Microprojection drawing of a series of sagittal sections through whole head of barfin flounder larvae on (A) Days 7, (B) 21, (C) 28, and (D) 42, showing the distribution of neuronal somata expressing sGnRH (blue circle), cGnRH-II (red circle) and sbGnRH (yellow circle); MT midbrain tegmentum, OB olfactory bulb, OE olfactory epithelium, ON olfactory nerve, OT optic tectum, POA preoptic area, T telencephalon. Bars indicate 0.5 mm. Reprinted with permission from Zoological Science, 21, Amano et al., Ontogenic development of three GnRH systems in the brain of a pleuronectiform fish, barfin flounder, 311–317, Figs. 1A, C, D, E, © 2004, Zoological Society of Japan. characterization of a KiSS1 gene has been reported in zebrafish Danio rerio (van Aerle et al. 2008) and medaka (Kanda et al. 2008). In future, the relationship between GnRH system and the KiSS1/GPR54 system in teleost fish should be clarified to understand the reproductive biology thoroughly. Acknowledgments I thank Professor Katsumi Aida for giving me the opportunity to write this monograph. I also extend my thanks to the following persons for their contributions to the original research: Prof. Makito Kobayashi, Dr. Koichi Okuzawa, Dr. Naoto Okumoto, Dr. Shoji Kitamura, Dr. Kazumasa Ikuta, Prof. Yoshihisa Hasegawa, Prof. Yoshitaka Oka, Prof. Hiroshi Kawauchi, Prof. Kunio Yamamori, Prof. Akiyoshi Takahashi, Prof. Akihisa Urano, Dr. Susumu Hyodo, Dr. Masayuki Iigo, 67 Fig. 37. (A) Sagittal section through the brain of barfin flounder on Day 7. Neuronal somata expressing cGnRH-II mRNA are observed in the MT near the ventricle (single arrowheads). (B) Sagittal section through the OE of barfin flounder on Day 21. Neuronal somata expressing sGnRH mRNA are observed in the vicinity of the OE (single arrowhead). (C) Sagittal section through the ON and OB of barfin flounder on Day 28. Neuronal somata expressing sGnRH mRNA are observed in the TN (single arrowheads). (D) Sagittal section through the terminal nerve ganglion of barfin flounder on Day 28. Neuronal somata expressing sGnRH mRNA are observed in the TN (single arrowheads). (E) Sagittal section through the POA of barfin flounder on Day 42. Neurons expressing sbGnRH mRNA are observed (single arrowheads). Bars indicates 20 µm. Reprinted with permission from Zoological Science, 21, Amano et al., Ontogenic development of three GnRH systems in the brain of a pleuronectiform fish, barfin flounder, 311–317, Figs. 3B, 2B, 2D, 2F, 4B, © 2004, Zoological Society of Japan. Dr. Arimune Munakata, Dr. Kataaki Okubo, Mr. Takeshi Yamanome, and Dr. Noriko Amiya. doi:10.5047/absm.2010.00302.0039 © 2010 TERRAPUB, Tokyo. All rights reserved. 68 M. Amano / Aqua-BioSci. Monogr. 3: 39–72, 2010 References Adams BA, Tello JA, Erchegyi J, Warby C, Hong DJ, Akinsanya KO, Mackie GO, Vale W, Rivier JE, Sherwood NM. Six novel gonadotropin-releasing hormones are encoded as triplets on each of two genes in the protochordate, Ciona intestinalis. Endocrinology 2003; 144: 1907–1919. Adelman JP, Mason AJ, Hayflick JS, Seeburg PH. Isolation of the gene and hypothalamic cDNA for the common precursor of gonadotropin-releasing hormone and prolactin release-inhibiting factor in human and rat. Proc. Natl. Acad. Sci. USA 1986; 83: 179–183. Aida K, Kato T, Awaji M. Effects of castration on the smoltification of precocious male masu salmon Oncorhynchus masou. Bull. Japan. Soc. Sci. Fish. 1984; 50: 565–571. Amano M, Oka Y, Aida K, Okumoto N, Kawashima S, Hasegawa Y. Immunocytochemical demonstration of salmon GnRH and chicken GnRH-II in the brain of masu salmon, Oncorhynchus masou. J. Comp. Neurol. 1991; 314: 587–597. Amano M, Aida K, Okumoto N, Hasegawa Y. Changes in salmon GnRH and chicken GnRH-II contents in the brain and pituitary, and GTH contents in the pituitary in female masu salmon, Oncorhynchus masou, from hatching through ovulation. Zool. Sci. 1992; 9: 375–386. Amano M, Hyodo S, Urano A, Okumoto N, Kitamura S, Ikuta K, Suzuki Y, Aida K. Activation of salmon gonadotropinreleasing hormone synthesis by 17α-methyltestosterone administration in yearling masu salmon, Oncorhynchus masou. Gen. Comp. Endocrinol. 1994; 95: 374–380. Amano M, Hyodo S, Kitamura S, Ikuta K, Suzuki Y, Urano A, Aida K. Salmon GnRH synthesis in the preoptic area and the ventral telencephalon is activated during gonadal maturation in female masu salmon. Gen. Comp. Endocrinol. 1995a; 99: 13–21. Amano M, Hyodo S, Kitamura S, Ikuta K, Urano A, Suzuki Y, Aida K. Short photoperiod accelerates preoptic and ventral telencephalic salmon GnRH synthesis and precocious maturation in underyearling male masu salmon. Gen. Comp. Endocrinol. 1995b; 99: 22–27. Amano M, Ikuta K, Kitamura S, Aida K. The maturation of the salmon GnRH system and its regulation by gonadal steroids in masu salmon. Fish Physiol. Biochem. 1997a; 17: 63–70. Amano M, Okumoto N, Aida K. Incomplete development of the brain-pituitary-gonadal axis may underlie the delay in the initiation of precocious maturation in male sockeye salmon. Fish. Sci. 1997b; 63: 873–876. Amano M, Kobayashi M, Okumoto N, Aida K. Low GnRH levels in the brain and the pituitary in triploid female sockeye salmon. Fish. Sci. 1998a; 64: 340–341. Amano M, Oka Y, Kitamura S, Ikuta K, Aida K. Ontogenic development of salmon GnRH and chicken GnRH-II systems in the brain of masu salmon. Cell Tissue Res. 1998b; 293: 427–434. Amano M, Ikuta K, Kitamura S, Aida K. Effects of photoperiod on salmon GnRH mRNA levels in brain of castrated underyearling precocious male masu salmon. Gen. Comp. Endocrinol. 1999; 115: 70–75. Amano M, Takahashi A, Yamanome T, Okubo K, Aida K, Yamamori K. Molecular cloning of three cDNAs encod- ing different GnRHs in the brain of barfin flounder. Gen. Comp. Endocrinol. 2002a; 126: 325–333. Amano M, Oka Y, Yamanome T, Okuzawa K, Yamamori K. Three GnRH systems in the brain and pituitary of a pleuronectiform fish, barfin flounder Verasper moseri. Cell Tissue Res. 2002b; 309: 323–329. Amano M, Okubo K, Ikuta K, Kitamura S, Okuzawa K, Yamada H, Aida K, Yamamori K. Ontogenic origin of salmon GnRH neurons in the ventral telencephalon and the preoptic area in masu salmon. Gen. Comp. Endocrinol. 2002c; 127: 256–262. Amano M, Okubo K, Yamanome T, Yamada H, Aida K, Yamamori K. Changes in brain GnRH mRNA and pituitary GnRH peptide during testicular maturation in barfin flounder. Comp. Biochem. Physiol. B 2004a; 138: 435–443. Amano M, Okubo K, Yamanome T, Oka Y, Kawaguchi N, Aida K, Yamamori K. Ontogenic development of three GnRH systems in the brain of a pleuronectiform fish, barfin flounder. Zool. Sci. 2004b; 21: 311–317. Amano M, Ikuta K, Kitamura S. Effects of a gonadotropinreleasing hormone antagonist on gonadotropin levels in masu salmon and sockeye salmon. J. Exp. Zool. A 2007; 307A: 535–541. Amano M, Pham KX, Amiya N, Yamanome T, Yamamori K. Changes in brain seabream GnRH mRNA and pituitary seabream GnRH peptide levels during ovarian maturation in female barfin flounder. Gen. Comp. Endocrinol. 2008; 158: 168–172. Amiya N, Amano M, Yamanome T, Yamamori K, Takahashi A. Effects of background color on GnRH and MCH levels in the barfin flounder brain. Gen. Comp. Endocrinol. 2008; 155: 88–93. Ando H, Swanson P, Kitani T, Koide N, Okada H, Ueda H, Urano A. Synergistic effects of salmon gonadotropinreleasing hormone and estradiol-17 β on gonadotropin subunit gene expression and release in masu salmon pituitary cells in vitro. Gen. Comp. Endocrinol. 2004; 137: 109–121. Ashihara M, Suzuki M, Kubokawa K, Yoshiura Y, Kobayashi M, Urano A, Aida K. Two differing precursor genes for the salmon-type gonadotropin-releasing hormone exist in salmonids. J. Mol. Endocrinol. 1995; 15: 1–9. Benfey TJ, Dye HM, Solar II, Donaldson EM. The growth and reproductive endocrinology of adult triploid Pacific salmonids. Fish Physiol. Biochem. 1989a; 6: 113–120. Benfey TJ, Dye HM, Donaldson EM. Estrogen-induced vitellogenin production by triploid coho salmon (Oncorhynchus kisutch), and its effect on plasma and pituitary gonadotropin. Gen. Comp. Endocrinol. 1989b; 75: 83–87. Bogerd J, Zandbergen T, Andersson E, Goos H. Isolation, characterization and expression of cDNAs encoding the catfish-type and chicken-II-type gonadotropin-releasinghormone precursors in the African catfish. Eur. J. Biochem. 1994; 222: 541–549. Bond CT, Francis RC, Fernald RD, Adelman JP. Characterization of complementary DNA encoding the precursor for gonadotropin-releasing hormone and its associated peptide from a teleost fish. Mol. Endocrinol. 1991; 5: 931–937. Breton B, Motin A, Billard R, Kah O, Geoffre S, Precigoux G. Immunoreactive gonadotropin-releasing hormone-like doi:10.5047/absm.2010.00302.0039 © 2010 TERRAPUB, Tokyo. All rights reserved. M. Amano / Aqua-BioSci. Monogr. 3: 39–72, 2010 material in the brain and pituitary gland during the periovulatory period in the brown trout (Salmo trutta L.): Relationships with the plasma and pituitary gonadotropin. Gen. Comp. Endocrinol. 1986; 61: 109–119. Burgus R, Butcher M, Amoss M, Ling N, Monahan M, River J, Fellows R, Blackwell R, Vale W, Guillemin R. Primary structure of the ovine hypothalamic luteinizing hormonereleasing factor (LRF). Proc. Natl. Acad. Sci. USA 1972; 69: 278–282. Chang JP, Cook H, Freedman GL,Wiggs AJ, Somoza GM, de Leeuw R, Peter RE. Use of a pituitary cell dispersion method and primary culture system for the studies of gonadotropinreleasing hormone action in the goldfish, Carassius auratus. I. Initial morphological, static, and cell column perfusion studies. Gen. Comp. Endocrinol. 1990; 77: 256–273. Chang JP, Wildman B, Van Goor F. Lack of involvement of arachidonic acid metabolism in chicken gonadotropinreleasing hormone II (cGnRH-II) stimulation of gonadotropin secretion in dispersed pituitary cells of goldfish, Carassius auratus: Identification of a major difference in salmon GnRH and chicken GnRH-II mechanisms of action. Mol. Cell. Endocrinol. 1991; 79: 75–83. Chiba A, Oka S, Honma Y. Ontogenetic development of gonadotropin-releasing hormone-like immunoreactive neurons in the brain of the chum salmon, Oncorhynchus keta. Neurosci. Lett. 1994; 178: 51–54. Counis R, Dufour S, Ribot G, Querat B, Fontaine YA, Jutisz M. Estradiol has inverse effects on pituitary glycoprotein hormone α -subunit messenger ribonucleic acid in the immature European eel and the gonadectomized rat. Endocrinology 1987; 121: 1178–1184. de Roux N, Genin E, Carel JC, Matsuda F, Chaussain JL, Milgrom E. Hypogonadotropic hypogonadism due to loss of function of the KiSS1-derived peptide receptor GPR54. Proc. Natl. Acad. Sci. USA 2003; 100: 10972–10976. Dufour S, Pasqualini C, Kerdelhue B, Fontaine YA. Presence and distribution of radioimmunoassayable LHRH in the European eel, Anguilla anguilla. Neuropeptides 1982; 3: 159–171. Dufour S, Fontaine YA, Kerdelhue B. Increase in brain and pituitary radio immunoassayable gonadotropin releasing hormone (GnRH) in the European silver eel treated with sexual steroid or human chorionic gonadotropin. Neuropeptides 1985; 6: 495–502. Dufour S, Montero M, Le Belle N, Bassompierre M, King JA, Millar RP, Peter RE. Differential distribution and response to experimental sexual maturation of two forms of brain gonadotropin-releasing hormone (GnRH) in the European eel, Anguilla anguilla. Fish Physiol. Biochem. 1993; 11: 99–106. Dunn IC, Chen Y, Hook C, Sharp PJ, Sang HM. Characterization of the chicken preprogonadotrophin-releasing hormone-I gene. J. Mol. Endocrinol. 1993; 11: 19–29. Gen K, Maruyama O, Kato T, Tomozawa K, Wakabayashi K, Kato Y. Molecular cloning of cDNAs encoding two types of gonadotropin α subunit from the masu salmon, Oncorhynchus masou; construction of specific oligonucleotides for the α 1 and α 2 subunits. J. Mol. Endocrinol. 1993; 11: 265–273. 69 Gentile F, Lira O, Marcano-de Cotte D. Relationship between brain gonadotropin-releasing hormone (GnRH) and seasonal reproductive cycle of “caribe colorado,” Pygocentrus notatus. Gen. Comp. Endocrinol. 1986; 64: 239–245. Gielen JTh, Goos HJTh, Peute J, van den Bosch RA, van Oordt PGWJ. The brain-pituitary-gonadal axis in the rainbow trout, Salmo gairdneri: Gonadal hormones and the maturation of gonadotropic cells. Cell Tissue Res. 1982; 225: 45–56. González-Martínez D, Madigou T, Zmora N, Anglade I, Zanuy S, Zohar Y, Elizur A, Muñoz-Cueto JA, Kah O. Differential expression of three different prepro-GnRH (gonadotrophin-releasing hormone) messengers in the brain of the European sea bass (Dicentrarchus labrax). J. Comp. Neurol. 2001; 429: 144–155. González-Martínez D, Zmora N, Zanuy S, Sarasquete C, Elizur A, Kah O, Muñoz-Cueto JA. Developmental expression of three different prepro-GnRH (gonadotrophinreleasing hormone) messengers in the brain of the European sea bass (Dicentrarchus labrax). J. Chem. Neuroanat. 2002; 23: 255–267. Goos HJTh. Steroid feedback on gonadotropin secretion. In: Idler DR, Crim LW, Walsh JM (eds). Proc. Third Int. Symp. Reprod. Physiol. Fish. Memorial University Press, St. John’s. 1987; pp. 16–20. Goos HJTh, de Leeuw R, Cook H, van Oordt PGWJ. Gonadotropic hormone-releasing hormone (GnRH) bioactivity in the brain of immature rainbow trout, Salmo gairdneri: The effect of testosterone. Gen. Comp. Endocrinol. 1986; 64: 80–84. Grober MS, Myers TR, Marchaterre MA, Bass AH, Myers DA. Structure, localization, and molecular phylogeny of a GnRH cDNA from a paracanthopterygian fish, the plainfin midshipman (Porichthys notatus). Gen. Comp. Endocrinol. 1995; 99: 85–99. Guilgur LG, Moncaut NP, Canario AVM, Somoza GM. Evolution of GnRH ligands and receptors in gnathostomata. Comp. Biochem. Physiol. A, Mol. Integr. Physiol. 2006; 144: 272–283. Habibi HR. Homologous desensitization of gonadotropinreleasing hormone (GnRH) receptors in the goldfish pituitary: effects of native GnRH peptides and a synthetic GnRH antagonist. Biol. Reprod. 1991; 44: 275–283. Hassin S, Clarie M, Holland H, Zohar Y. 1999. Ontogeny of follicle-stimulating hormone and luteinizing hormone gene expression during pubertal development in the female striped bass, Morone saxatilis (Teleostei). Biol. Reprod. 1999; 61: 1608–1615. Iguchi K, Ito F, Ikuta K, Yamaguchi M. Sexual dimorphism in the anal fin of ayu Plecoglossus altivelis. Nippon Suisan Gakkaishi 1991; 57: 1501–1505. Iwakoshi E, Takuwa-Kuroda K, Fujisawa Y, Hisada M, Ukena K, Tsutsui K, Minakata H. Isolation and characterization of a GnRH-like peptide from Octopus vulgaris. Biochem. Biophys. Res. Commun. 2002; 291: 1187–1193. Kah O, Anglade I, Lepretre E, Dubourg P, de Monbrison D. The reproductive brain in fish. Fish. Physiol. Biochem. 1993; 11: 85–98. Kanda S, Akazome Y, Matsunaga T, Yamamoto N, Yamada S, Tsukamura H, Maeda K, Oka Y. Identification of KiSS-1 doi:10.5047/absm.2010.00302.0039 © 2010 TERRAPUB, Tokyo. All rights reserved. 70 M. Amano / Aqua-BioSci. Monogr. 3: 39–72, 2010 product kisspeptin and steroid-sensitive sexually dimorphic kisspeptin neurons in medaka (Oryzias latipes). Endocrinology 2008; 149: 2467–2476. Kato Y, Gen K, Maruyama O, Tomizawa K, Kato T. Molecular cloning of cDNAs encoding two gonadotrophin β subunits (GTH-Iβ and GTH-IIβ) from the masu salmon, Oncorhynchus masou: rapid divergence of the GTH-Iβ gene. J. Mol. Endocrinol. 1993; 11: 275–282. Kauffman AS, Rissman EF. The evolutionarily conserved gonadotropin-releasing hormone II modifies food intake. Endocrinology 2004; 145: 686–691. Kavanaugh SI, Nozaki M, Sower SA. Origins of gonadotropinreleasing hormone (GnRH) in vertebrates: identification of a novel GnRH in a basal vertebrate, the sea lamprey. Endocrinology 2008; 149: 3860–3869. Kawauchi H, Sower SA. The dawn and evolution of hormones in the adenohypophysis. Gen. Comp. Endocrinol. 2006; 148: 3–14. Kawauchi H, Suzuki K, Itoh H, Swanson P, Naito N, Nagahama Y, Nozaki M, Nakai Y, Itoh S. The duality of teleost gonadotropins. Fish Physiol. Biochem. 1989; 7: 29–38. Kim M, Oka Y, Amano M, Kobayashi M, Okuzawa K, Hasegawa Y, Kawashima S, Suzuki Y, Aida K. Immunocytochemical localization of sGnRH and cGnRH-II in the brain of goldfish, Carassius auratus. J. Comp. Neurol. 1995; 356: 72–82. Kim M, Kobayashi M, Oka Y, Amano M, Kawashima S, Aida K. Effects of olfactory tract section on the immunohistochemical distribution of brain GnRH fibers in the female goldfish, Carassius auratus. Zool. Sci. 2001; 18: 241–248. King JA, Millar RP. Evolution of gonadotropin-releasing hormones. Trends Endocrinol. Metab. 1992; 3: 339–346. Klungland H, Lorens JB, Andersen Ø, Kisen GØ, Alestrøm P. The Atlantic salmon prepro-gonadotropin releasing hormone gene and mRNA. Mol. Cell. Endocrinol. 1992; 84: 167–174. Kobayashi M, Aida K, Sakai H, Kaneko T, Asahina K, Hanyu I, Ishii S. Radioimmunoassay for salmon gonadotropin. Nippon Suisan Gakkaishi 1987; 53: 995–1003. Kobayashi M, Amano M, Hasegawa Y, Okuzawa K, Aida K. Effects of olfactory tract section on brain GnRH distribution, plasma gonadotropin levels, and gonadal stage in goldfish. Zool. Sci. 1992; 9: 765–773. Kobayashi M, Amano M, Kim M, Furukawa K, Hasegawa Y, Aida K. Gonadotropin-releasing hormones of terminal nerve origin are not essential to ovarian development and ovulation in goldfish. Gen. Comp. Endocrinol. 1994; 95: 192–200. Kubo T. Studies on the life history of the “masu” salmon (Oncorhynchus masou) in Hokkaido. Sci. Rep. Hokkaido Salmon Hatchery 1980; 34: 1–95 (in Japanese). Kudo H, Ueda H, Kawamura H, Aida K, Yamauchi K. Ultrastructural demonstration of salmon-type gonadotropin-releasing hormone (sGnRH) in the olfactory system of masu salmon (Oncorhynchus masou). Neurosci. Lett. 1994; 166: 187–190. Kudo H, Hyodo S, Ueda H, Hiroi O, Aida K, Urano A, Yamauchi K. Cytophysiology of gonadotropin-releasing-hormone neurons in chum salmon (Oncorhynchus keta) forebrain before and after upstream migration. Cell Tissue Res. 1996; 284: 261–267. Lethimonier C, Madigou T, Muñoz-Cueto JA, Lareyre JJ, Kah O. Evolutionary aspects of GnRHs, GnRH neuronal systems and GnRH receptors in teleost fish. Gen. Comp. Endocrinol. 2004; 135: 1–16. Lewis KA, Swanson P, Sower SA. Changes in brain gonadotropin-releasing hormone, pituitary and plasma gonadotropins, and plasma thyroxine during smoltification in chinook salmon (Oncorhynchus tschawytscha). Gen. Comp. Endocrinol. 1992; 87: 461–470. Lin X-W, Peter RE. Expression of salmon gonadotropinreleasing hormone (GnRH) and chicken GnRH-II precursor messenger ribonucleic acids in the brain and ovary of goldfish. Gen. Comp. Endocrinol. 1996; 101: 282–296. Lincoln RF, Scott AP. Sexual maturation in triploid rainbow trout, Salmo gairdneri Richardson. J. Fish Biol. 1984; 25: 385–392. Lundqvist H. Influence of photoperiod on growth in Baltic salmon parr (Salmo salar L.) with special reference to the effect of precocious sexual maturation. Can. J. Zool. 1980; 58: 940–944. Mason AJ, Hayflick JS, Zoeller RT, Young WS III, Philips HS, Nikolics K, Seeburg PH. A deletion truncating the gonadotropin-releasing hormone gene is responsible for hypogonadism in the hpg mouse. Science 1986; 234: 1366–1371. Matsuo H, Baba Y, Nair RMG, Arimura A, Schaly AV. Structure of the porcine LH- and FSH-releasing hormone. I. The proposed amino acid sequence. Biochem. Biophys. Res. Commun. 1971; 43: 1334–1339. Murakami S, Seki T, Wakabayashi K, Arai Y. The ontogeny of luteinizing hormone-releasing hormone (LHRH) producing neurons in the chick embryo: possible evidence for migrating LHRH neurons from the olfactory epithelium expressing a highly polysialylated neural cell adhesion molecule. Neurosci. Res. 1991; 12: 421–431. Murakami S, Kikuyama S, Arai Y. The origin of the luteinizing hormone-releasing hormone (LHRH) neurons in the newts (Cynops pyrrhogaster): the effect of olfactory placode ablation. Cell Tissue Res. 1992; 269: 21–27. Muske LE, Moore FL. Ontogeny of immunoreactive gonadotropin-releasing hormone neuronal systems in amphibians. Brain Res. 1990; 534: 177–187. Naito N, Hyodo S, Okumoto N, Urano A, Nakai Y. Differential production and regulation of gonadotropins (GTH I and GTH II) in the pituitary gland of rainbow trout, Oncorhynchus mykiss, during ovarian development. Cell Tissue Res. 1991; 266: 457–467. Nakamura M, Nagahama Y, Iwahashi M, Kojima M. Ovarian structure and plasma steroid hormones of triploid female rainbow trout. Nippon Suisan Gakkaishi 1987; 53: 1105 (in Japanese). Ohtaki T, Shintani Y, Honda S, Matsumoto H, Hori A, Kanehashi K, Terao Y, Kumano S, Takatsu Y, Masuda Y, Ishibashi Y, Watanabe T, Asada M, Yamada T, Suenaga M, Kitada C, Usuki S, Kurokawa T, Onda H, Nishimura O, Fujino M. Metastasis suppressor gene KiSS-1 encodes peptide ligand of a G-protein-coupled receptor. Nature 2001; 411: 613–617. doi:10.5047/absm.2010.00302.0039 © 2010 TERRAPUB, Tokyo. All rights reserved. M. Amano / Aqua-BioSci. Monogr. 3: 39–72, 2010 Oka Y. Gonadotropin-releasing hormone (GnRH) cells of the terminal nerve as a model neuromodulator system. Neurosci. Lett. 1992; 142: 119–122. Oka Y. GnRH neuronal system of fish brain as a model system for the study of peptidergic neuromodulation. In: Parhar IS, Sakuma Y (eds). GnRH Neurons: Gene to behavior. Brain Shuppan Publishers, Tokyo. 1997; pp. 245–276. Oka Y. Three types of gonadotrophin-releasing hormone neurons and steroid-sensitive sexually dimorphic kisspeptin neurons in teleosts. J. Neuroendocrinol. 2009; 21: 334– 338. Oka Y, Ichikawa M. Gonadotropin-releasing hormone (GnRH) immunoreactive system in the brain of the dwarf gourami (Colisa lalia) as revealed by light microscopic immunocytochemistry using a monoclonal antibody to common amino acid sequence of GnRH. J. Comp. Neurol. 1990; 300: 511–522. Oka Y, Matsushima T. Gonadotropin-releasing hormone (GnRH)-immunoreactive terminal nerve cells have intrinsic rhythmicity and project widely in the brain. J. Neurosci. 1993; 13: 2161–2176. Okubo K, Nagahama Y. Structural and functional evolution of gonadotropin-releasing hormone in vertebrates. Acta Physiol. 2008; 193: 3–15. Okubo K, Amano M, Yoshiura Y, Suetake H, Aida K. A novel form of gonadotropin-releasing hormone in the medaka, Oryzias latipes. Biochem. Biophys. Res. Commun. 2000; 276: 298–303. Okuzawa K. Puberty in teleosts. Fish Physiol. Biochem. 2002; 26: 31–41. Okuzawa K, Kobayashi M. Gonadotropin-releasing hormone neuronal systems in the teleostean brain and functional significance. In: Prasada Rao PD, Peter RE (eds). Neural Regulation in the Vertebrate Endocrine System. Kluwer Academic/Plenum Publishers, New York. 1999; pp. 85–100. Okuzawa K, Amano M, Kobayashi M, Aida K, Hanyu I, Hasegawa Y, Miyamoto K. Differences in salmon GnRH and chicken GnRH-II contents in discrete brain areas of male and female rainbow trout according to age and stage of maturity. Gen. Comp. Endocrinol. 1990; 80: 116–126. Okuzawa K, Araki K, Tanaka H, Kagawa H, Hirose K. Molecular cloning of a cDNA encoding the prepro-salmon gonadotropin-releasing hormone of the red seabream. Gen. Comp. Endocrinol. 1994; 96: 234–242. Okuzawa K, Granneman J, Bogerd J, Goos HJTh, Zohar Y, Kagawa H. Distinct expression of GnRH genes in the red seabream brain. Fish Physiol. Biochem. 1997; 17: 71–79. Parhar IS, Koibuchi N, Sakai M, Iwata M, Yamaoka S. Gonadotropin-releasing hormone (GnRH): expression during salmon migration. Neurosci. Lett. 1994; 172: 15– 18. Parhar IS, Iwata M, Pfaff DW, Schwanzel-Fukuda M. Embryonic development of gonadotropin-releasing hormone neurons in the sockeye salmon. J. Comp. Neurol. 1995; 362: 256–270. Peter RE, Yu KL, Marchant TA, Rosenblum PM. Direct neural regulation of the teleost adenohypophysis. J. Exp. Zool. 1990; S4: 84–89. Pham KX, Amano M, Amiya N, Kurita Y, Shimizu A, Yamamori K. Immunohistochemical localization of three 71 GnRH systems in the brain and pituitary of Japanese flounder. Fish. Sci. 2007; 73: 1113–1122. Powell JF, Reska-Skinner SM, Prakash MO, Fischer WH, Park M, Rivier JE, Craig AG, Mackie GO, Sherwood NM. Two new forms of gonadotropin-releasing hormone in a protochordate and the evolutionary implications. Proc. Natl. Acad. Sci. USA 1996; 93: 10461–10464. Prat F, Sumpter JP, Tyler CR. Validation of radioimmunoassays for two salmon gonadotropins (GTH I and GTH II) and their plasma concentrations throughout the reproductive cycle in male and female rainbow trout (Oncorhynchus mykiss). Biol. Reprod. 1996; 54: 1375–1382. Schreibman MP, Halpern-Sebold L, Ferin M, MargolisKazan H, Goos HJTh. The effect of hypophysectomy and gonadotropin administration on the distribution and quantity of LH-RH in the brains of platyfish: A combined immunocytochemistry and radioimmunoassay study. Brain Res. 1983; 267: 293–300. Schreibman MP, Margolis-Nunno H, Halpern-Sebold LR, Goos HJTh, Perlman PW. The influence of androgen administration on the structure and function of the brainpituitary-gonadal axis of sexually immature platyfish, Xiphophorus maculatus. Cell Tissue Res. 1986; 245: 519–524. Schulz RW, van der Sanden MCA, Bosma PT, Goos HJTh. Effects of gonadotropin-releasing hormone during the pubertal development of the male African catfish (Clarias gariepinus): gonadotrophin and androgen levels in plasma. J. Endocrinology 1994a; 140: 265–273. Schulz RW, van der Corput L, Janssen-Dommerholt J, Goos HJTh. Sexual steroids during puberty in male African catfish (Clarias gariepinus); serum levels and gonadotropinstimulated testicular secretion in vitro. J. Comp. Physiol. B 1994b; 164: 195–205. Schwanzel-Fukuda M, Pfaff DW. Origin of luteinizing hormonereleasing hormone neurons. Nature 1989; 338: 161–164. Seeburg PH, Adelman JP. Characterization of cDNA for precursor of human luteinizing hormone releasing hormone. Nature 1984; 311: 666–678. Seminara SB, Messager S, Chatzidaki EE, Thresher RR, Acierno JS Jr, Shagoury JK, Bo-Abbas Y, Kuohung W, Schwinof KM, Hendrick AG, Zahn D, Dixon J, Kaiser UB, Slaugenhaupt SA, Gusella JF, O’Rahilly S, Carlton MBL, Crowley WF Jr, Aparicio SAJR, Colledge WH. The GPR54 gene as a regulator of puberty. N. Engl. J. Med. 2003; 349: 1614–1627. Senthilkumaran B, Okuzawa K, Gen K, Ookura T, Kagawa H. Distribution and seasonal variations in levels of three native GnRHs in the brain and pituitary of perciform fish. J. Neuroendocrinol. 1999; 11: 181–186. Sherwood NM, Lovejoy DA, Coe IR. Origin of mammalian gonadotropin-releasing hormones. Endocr. Rev. 1993; 14: 241–254. Suzuki K, Kawauchi H, Nagahama Y. Isolation and characterization of two distinct gonadotropins from chum salmon pituitary glands. Gen. Comp. Endocrinol. 1988a; 71: 292–301. Suzuki K, Kawauchi H, Nagahama Y. Isolation and characterization of subunits from two distinct salmon gonadotropins. Gen. Comp. Endocrinol. 1988b; 71: 302–306. doi:10.5047/absm.2010.00302.0039 © 2010 TERRAPUB, Tokyo. All rights reserved. 72 M. Amano / Aqua-BioSci. Monogr. 3: 39–72, 2010 Suzuki K, Kanamori A, Nagahama Y, Kawauchi H. Development of salmon GTH I and GTH II radioimmunoassays. Gen. Comp. Endocrinol. 1988c; 71: 459–467. Suzuki M, Hyodo S, Kobayashi M, Aida K, Urano A. Characterization and localization of mRNA encoding the salmon-type gonadotrophin-releasing hormone precursor of the masu salmon. J. Mol. Endocrinol. 1992; 9: 73–82. Swanson P, Bernard M, Nozaki M, Suzuki K, Kawauchi H, Dickhoff WW. Gonadotropins I and II in juvenile coho salmon. Fish Physiol. Biochem. 1989; 7: 169–176. Takashima F, Yamada Y. Control of maturation in masu salmon by manipulation of photoperiod. Aquaculture 1984; 43: 243–257. Tyler CR, Sumpter JP, Kawauchi H, Swanson P. Involvement of gonadotropin in the uptake of vitellogenin into vitellogenic oocytes in the rainbow trout, Oncorhynchus mykiss. Gen. Comp. Endocrinol. 1991; 84: 291–299. van Aerle R, Kille P, Lange A, Tyler CR. Evidence for the existence of a functional Kiss1/Kiss1 receptor pathway in fish. Peptides 2008; 29: 57–64. Volkoff H, Peter RE. Actions of two forms of gonadotropin releasing hormone and a GnRH antagonist on the spawning behavior of the goldfish Carassius auratus. Gen. Comp. Endocrinol. 1999; 116: 347–355. White RB, Fernald RD. Ontogeny of gonadotropin-releasing hormone (GnRH) gene expression reveals a distinct origin for GnRH-containing neurons in the midbrain. Gen. Comp. Endocrinol. 1998; 112: 322–329. White SA, Bond CT, Francis RC, Kasten TL, Fernald RD, Adelman JP. A second gene for gonadotropin-releasing hormone: cDNA and expression pattern in the brain. Proc. Natl. Acad. Sci. USA 1994; 91: 1423–1427. White SA, Kasten TL, Bond CT, Adelman JP, Fernald RD. Three gonadotropin-releasing hormone genes in one organism suggest novel roles for an ancient peptide. Proc. Natl. Acad. Sci. USA 1995; 92: 8363–8367. Wray S, Grant P, Gainer H. Evidence that cells expressing luteinizing hormone-releasing hormone mRNA in the mouse are derived from progenitor cells in the olfactory placode. Proc. Natl. Acad. Sci. USA 1989; 86: 8132–8136. Xiong F, Chin R, Gong Z, Suzuki K, Kitching R, MajumdarSonnylal S, Elsholtz HP, Hew CL. Control of salmon pituitary hormone gene expression. Fish Physiol. Biochem. 1993; 11: 63–70. Xiong F, Liu D, Elsholtz HP, Hew CL. The chinook salmon gonadotropin IIβ subunit gene contains a strong minimal promoter with a proximal negative element. Mol. Endocrinol. 1994a; 8: 771–781. Xiong F, Liu D, Le Drean Y, Elsholtz HP, Hew CL. Differential recruitment of steroid hormone response elements may dictate the expression of the pituitary gonadotropin II β subunit gene during salmon migration. Mol. Endocrinol. 1994b; 8: 782–793. Yamada H, Satoh R, Yamashita T, Kambegawa A, Iwata M. Development of a time-resolved fluoroimmunoassay (TR-FIA) for testosterone: measurement of serum testosterone concentrations after testosterone treatment in the rainbow trout (Oncorhynchus mykiss). Gen. Comp. Endocrinol. 1997; 106: 181–188. Yamamoto N, Oka Y, Amano M, Aida K, Hasegawa Y, Kawashima S. Multiple gonadotropin-releasing hormone (GnRH)-immunoreactive systems in the brain of the dwarf gourami, Colisa lalia: Immunohistochemistry and radioimmunoassay. J. Comp. Neurol. 1995; 355: 354–368. Yamamoto N, Oka Y, Kawashima S. Lesions of gonadotropinreleasing hormone-immunoreactive terminal nerve cells: effects on the reproductive behavior of male dwarf gouramis. Neuroendocrinol. 1997; 65: 403–412. Yamamoto N, Parhar IS, Sawai N, Oka Y, Ito H. Preoptic gonadotropin-releasing hormone (GnRH) neurons innervate the pituitary in teleosts. Neurosci. Res. 1998; 31: 31–38. Yu KL, Nahorniak CS, Peter RE, Corrigan A, Rivier JE, Vale WW. Brain distribution of radioimmunoassayable gonadotropin-releasing hormone in female goldfish: Seasonal variation and periovulatory changes. Gen. Comp. Endocrinol. 1987; 67: 234–246. Yu KL, Sherwood NM, Peter RE. Differential distribution of two molecular forms of gonadotropin-releasing hormone in discrete brain areas of goldfish (Carassius auratus). Peptides 1988; 9: 625–630. Zhang L, Tello JA, Zhang W, Tsai PS. Molecular cloning, expression pattern, and immunocytochemical localization of a gonadotropin-releasing hormone-like molecule in the gastropod mollusk, Aplysia californica. Gen. Comp. Endocrinol. 2008; 156: 201–209. Zohar Y, Muñoz-Cueto JA, Elizur A, Kah O. Neuroendocrinology of reproduction in teleost fish. Gen. Comp. Endocrinol. 2010; 165: 438–455. doi:10.5047/absm.2010.00302.0039 © 2010 TERRAPUB, Tokyo. All rights reserved.