Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
Metalloprotein wikipedia , lookup
Ridge (biology) wikipedia , lookup
Gene expression wikipedia , lookup
Metabolic network modelling wikipedia , lookup
Multi-state modeling of biomolecules wikipedia , lookup
Endogenous retrovirus wikipedia , lookup
Deoxyribozyme wikipedia , lookup
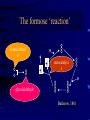
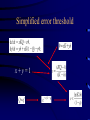
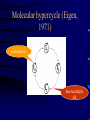
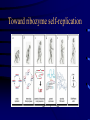
Evolution and the origin of life Eörs Szathmáry München Collegium Budapest Eötvös University Chemical evolution Units of evolution 1. multiplication 2. heredity 3. variation hereditary traits affecting survival and/or reproduction Gánti’s chemoton model (1974) metabolism template copying membrane growth ALL THREE SUBSYSTEMS ARE AUTOCATALYTIC The latest edition: OUP 2003 • After several editions in Hungarian • Two previous books (the Principles and Contra Crick) plus one essay • Essays appreciating the biological and philosophical importance Pathways of supersystem evolution metabolism MB boundary MT template BT MBT INFRABIOLOGICAL SYSTEMS What about replication? • Replication from a chemical point of view always rests on autocatalysis • The basic form is A+X 2A+Y • very important for biology • Much more general than DNA The formose ‘reaction’ formaldehyd e autocatalysi s glycolaldehyde Butlerow, 1861 Replication in the formose reaction • • • • • • • Replication is non-informational Autocatalysis – YES Heredity – NO Good for metabolism Not good for genetics Butlerow was born on the 15th Sept, 1829 He was regarded as one of the best lecturers of his time. His lectures were lucid and thorough, yet his language was colourful. Local society often preferred his lectures to the theatre Primitive ancestry of the reverse citric acid cycle • Was proposed by Günter Wächtershäuser (1990) • Coupled to CO2 fixation and pyrite formation around deep-sea hydrothermal vents The main problem of the origin of life is metabolite channelling • Enzymes speed up reactions relative to the unwanted reactions • Spontaneous decay reactions abound • Maintenance, not only reproduction, requires autocatalysis dx/ dt = k x – d x = 0 All network models neglecting side reactions are seriously incomplete • E.g. protein networks • In model assumptions, a reaction is either good or neutral for the system – but the number of harmful transformations is in fact much higher • Did life emerge from a chemical canyon? Chemical evolution was a race between tar formation and life formation Chemical networks Life Tar What fraction of planets would end up with just tar? Another case: von Kiedrowski’s replicators Von Kiedrowski’s replicator Peptide replicator networks • Theory with experiment • J. Mol. Evol., forthcoming Does temperature cycling work? Elongation taxes the system badly Classification of replicators Limited heredity Holistic formose Modular Von Kiedrowski Unlimited heredity genes Limited (number of individuals) > (number of types) Unlimited (# of individuals) << (# of types) A crucial insight: Eigen’s paradox (1971) • Early replication must have been errorprone • Error threshold sets the limit of maximal genome size to <100 nucleotides • Not enough for several genes • Unlinked genes will compete • Genome collapses • Resolution??? Simplified error threshold x+y=1 Molecular hypercycle (Eigen, 1971) autocatalysis heterocatalytic aid Parasites in the hypercycle (JMS) short circuit parasite “Hypercyles spring to life”… • Cellular automaton simulation on a 2D surface • Reaction-diffusion • Emergence of mesoscopic structure • Conducive to resistance against parasites • Good-bye to the wellstirred flow reactor …but then die if modelled in more detail • Are not resistant to short-circuits • Collapse if the adhesive surface is patchy (the mesoscopic structure collapses) • Only compartmentation saves them RNA structure and the error theshold: Kun, Santos, Szathmáry (2005) Nature Genetics 37, 1008-1011. • • • • • The 3D shape of the molecule Enzymatic activity depends on the structure Phenotype of a ribozyme is the structure There are fewer structures than sequences A few mutations in the sequence usually do not change the structure • The 2D structure can be computed easily Hairpin Ribozyme N = 50 H1 1 H2 H3 H4 20 AAC A G A A 10 30 5’ aaacaGAGAAGUcaACCAg CAC G ||||| loop B ||| u loop A AUGGUc CA GUG u 3’ G U UUAUA 50 40 39/50 (78%) of the positions were mutated, we used 142 mutants Neurospora Varkund Satellite Ribozyme IV 680 V 690 a cuGaAauuG-U-CguAgCAGU U G ||||||||| A u |||||||| aCuUuaaC GUaUUGUCA u G g C U710 670 G U 700 C-G UUA C-G III A-U A 660 C-G U-A720 730 640 650 740 C-G A A AAGUG-A-CGGuAuUggc g A GCU gcgguaguaaGc AgG 5’ |||||| ||| ||| ||| |||||||||| u cguuCg-CcC GAACACGA CACGACGUUaUgAcuga a 3’ uaagag 770 760 750 N = 144 780 II VI 83/144 (57%) of the positions were mutated, we used 183 mutants Neutral mutions tame the error threshold • Extrapolation from the available mutants as samples to the whole fitness landscape • Accuracy of viral RNA polymerases would be sufficient to run the genome of a ribo-organism of about 70 genes Error rates and the origin of replicators Nature 420, 360-363 (2002). Replicase RNA Other RNA Increase in efficiency • Target efficiency: the acceptance of help • Replicase efficiency: how much help it gives • Copying fidelity • Trade-off among all three traits: worst case The dynamics becomes interesting on the rocks! Evolving population Error rate Replicase activity • Molecules interact with their neighbours • Have limited diffusion on the surface Toward ribozyme self-replication Towards a general polymerase Anabolic autocatalysis I Anabolic autocatalysis II Szostak’s vesicles E - FLUX Fet Open Contract n° FP7-225167 Evolvability of molecular systems (analysis of the GARD model) The trigger: Doron Lancet’s GARD model E - FLUX Fet Open Contract n° FP7-225167 The mathematical framework for the analysis GARD dynamics Eigen equation Eigen matrix with huge off-diagonal terms E - FLUX Fet Open Contract n° FP7-225167 Hidden compartmentalization and no selection Kauffman: Reflexively autocatalytic protein networks (1986) Current investigations • • • • • Evolvability is possible only in compartments Occasionally new autocatalytic loops appear Can be inherited from one cell to the daugther Can be selected for, give some evolution GARD is shadow of protein networks is a shadow of template replicators The stochastic corrector model for compartmentation Szathmáry, E. & Demeter L. (1987) Group selection of early replicators and the origin of life. J. theor Biol. 128, 463-486. Grey, D., Hutson, V. & Szathmáry, E. (1995) A re-examination of the stochastic corrector model. Proc. R. Soc. Lond. B 262, 29-35. Dynamics of the SC model • Independently reassorting genes • Selection for optimal gene composition between compartments • Competition among genes within the same compartment • Stochasticity in replication and fission generates variation on which natural selection acts • A stationary compartment population emerges Group selection of early replicators • Many more compartments than templates within any compartment • No migration (fusion) between compartments • Each compartment has only one parent • Group selection is very efficient • Selection for replication synchrony Chromosomes! Open questions • • • • • Origin of efficient replication Origin of full protocells Origin of transcription Origin of highly specific enzymes Origin of translation