Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Plant tolerance to herbivory wikipedia , lookup
History of herbalism wikipedia , lookup
Gartons Agricultural Plant Breeders wikipedia , lookup
Plant stress measurement wikipedia , lookup
Historia Plantarum (Theophrastus) wikipedia , lookup
History of botany wikipedia , lookup
Ornamental bulbous plant wikipedia , lookup
Plant use of endophytic fungi in defense wikipedia , lookup
Venus flytrap wikipedia , lookup
Plant defense against herbivory wikipedia , lookup
Evolutionary history of plants wikipedia , lookup
Plant breeding wikipedia , lookup
Plant nutrition wikipedia , lookup
Plant secondary metabolism wikipedia , lookup
Plant evolutionary developmental biology wikipedia , lookup
Plant ecology wikipedia , lookup
Plant morphology wikipedia , lookup
Plant physiology wikipedia , lookup
Sustainable landscaping wikipedia , lookup
Plant reproduction wikipedia , lookup
Flowering plant wikipedia , lookup
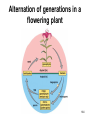
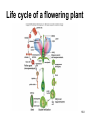
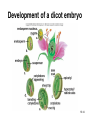
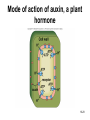
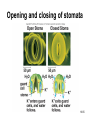
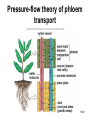
Chapter 10: Plant Reproduction, Growth, and Development 10-1 Sexual Reproduction in Flowering Plants Sexual reproduction requires gametes, often as egg and sperm. In flowering plants, the structures that produce the egg and sperm are located within the flower. Flowers have special structures to enable fertilization of egg by sperm. 10-2 Structure of Flowers The reproductive portions of a flower are the stamen, consisting of a stalk-like filament bearing an anther, and the pistil, made up of stigma, style, and ovary. The ovary contains one or more ovules. Sepals enclose a whorl of petals that are usually colored to attract pollinators. 10-3 Flower structure 10-4 Alternation of Generations Plant life cycles includes two alternating generations. The sporophyte (2N) produces haploid spores by meiosis. A spore develops into a haploid gametophyte that produces gametes. The sporophyte of flowering plants is dominant and produces two types of spores, microspores and megaspores. 10-5 Alternation of generations in a flowering plant 10-6 Microspores within the anther mature into pollen grains that are the microgametophyte generation. A megaspore within an ovule develops into the megagametophyte generation that produces an egg. During pollination pollen grains move by wind or animal carrier from the anther to the stigma where a pollen grain produces a pollen tube. 10-7 Sperm travel down the pollen tube; one sperm unites with the egg and becomes a zygote that develops into an embryo. The other sperm unites with the polar nuclei within the megagametophyte. This produces triploid (3n) endosperm that nourishes the embryo. These two fusions are known as double fertilization. 10-8 Life cycle of a flowering plant 10-9 The ovule wall hardens and becomes the seed coat. The seed consists of the sporophyte embryo, stored food, and a seed coat. The ovary may develop into a fruit. 10-10 Growth and Development in Plants Plant growth and development involve cell division, cellular elongation, and differentiation of cells into tissues and organs. Development is a programmed series of stages from a simpler to a more complex form. Cellular differentiation is specialization during development. 10-11 Development of the Dicot Embryo After double fertilization, the singlecelled zygote lies beneath the endosperm nucleus. The dicot embryo has two cotyledons or seed leaves that take up and store nutrients from the endosperm. The monocot embryo has only one cotyledon that rarely stores food. 10-12 The epicotyl is the portion above the cotyledons and becomes shoot; the hypocotyl is the portion below the cotyledons and will become stem. The hypocotyl ends in the radicle that will grow into root tissue. The embryo plus stored food is now contained within a seed. 10-13 Development of a dicot embryo 10-14 Development of Seeds and Fruits In flowering plants, seeds are enclosed within a fruit that usually develops from the ovary. The ovary wall becomes the pericarp. Fleshy fruits have a fleshy pericarp; dry fruits have a dry pericarp. An aggregate fruit such as blackberry is derived from many ovaries on the flower. 10-15 Fruit diversity 10-16 Dispersal of Seeds Seeds are modified to be distributed far from the parent plant. Many disperse by wind or animal carrier and some float by ocean currents. Plants have various means, such as hooks, spines, or wings, to ensure that dispersal occurs. Birds and animals eat fruits and defecate seeds away from the parent plant. 10-17 Germination of Seeds Germination of seeds occurs if there is sufficient water, warmth, and oxygen to sustain growth. Dormancy may be required before germination, and some seeds require periods of cold or minimal moisture. In many fruits, germination does not occur until seeds are removed and washed; mechanical action may also be required. 10-18 Dicot Versus Monocot Development In dicots, the cotyledons supply nutrients to the developing plant. The epicotyl bears young leaves called a plumule. The hypocotyl becomes the stem and the radicle becomes the root. The dicot shoot is hook-shaped to protect the delicate plumule. 10-19 Common garden bean, a dicot 10-20 Germination in darkness causes etiolation, which involves elongation and lack of color. Monocots have only one cotyledon and the endosperm, rather than the cotyledon, is food-storage tissue. The coleoptile and coleorhiza are protective sheaths around the monocot plumule and radicle. 10-21 Corn, a monocot 10-22 Asexual Reproduction in Flowering Plants Non-differentiated meristem tissue allows a plant to reproduce by asexual vegetative propagation. Strawberry stolons, potato tuber eyes, sweet potato roots, tree suckers, African violet cuttings, and grapevine cuttings are common methods of asexual propagation. In horticulture, identical offspring produced by vegetative cuttings are clones. 10-23 Propagation of Plants in Tissue Culture Plant cells are totipotent, having all the genetic potential to become mature specialized plants. Using tissue culture, whole plants can be grown from protoplasts, naked plant cells produced by enzymes that digest cell walls. Cell suspension culture permits production of chemicals from single cells derived from leaf, stem, or root tissue. 10-24 Tissue culture trees 10-25 Genetic Engineering of Plants Various techniques introduce foreign DNA into protoplasts that are propagated in tissue culture. Adult plants are generated from these cells and could produce insecticideresistant plants or plants that can grow in nutrient-limited soil. Plants can also be engineered to treat human diseases. 10-26 Control of Plant Growth and Development Since each plant cell is totipotent, hormones have a role in determining cellular differentiation. Plant Hormones There are five common groups of plant hormones: auxins, gibberellins, cytokinins, abscisic acid, and ethylene. 10-27 Natural and synthetic hormones are also called plant growth regulators. Plant hormones elicit a physiological response in target cells by binding to a receptor protein in the plasma membrane. 10-28 Mode of action of auxin, a plant hormone 10-29 Stimulatory Hormones Auxins include indoleacetic acid (IAA). Apically produced auxin prevents the growth of axillary buds (a phenomenon called apical dominance), causes roots to develop, and prevents fruit drop. Auxin’s ability to cause cell elongation is involved in a plant’s response to light and gravity. 10-30 Cytokinins, such as zeatin and kinetin, promote cell division and help to regulate the plant cell cycle. Cytokinins interact with auxin to affect differentiation, producing roots or shoots or flowers depending on the ratio of these hormones. Cytokinin also prevents senescence, death of plant parts due to aging. Gibberellins promote growth of stems and can break the dormancy of seeds. 10-31 Effects of plant hormones 10-32 Inhibitory Hormones Abscisic acid (ABA), the stress hormone, initiates and maintains seed and bud dormancy and closes stomata in the fall. Ethylene ripens fruit by increasing enzyme activity, particularly cellulase that hydrolyzes cellulose. Ethylene in the air inhibits growth of plants in general, and along with low levels of auxin and gibberellin in a leaf, initiates abscission. 10-33 Effects of ethylene 10-34 Plant Responses to Environmental Stimuli Plant growth and development are influenced by environmental stimuli such as light, day length, gravity, and touch. Environmental signals determine the seasonality of growth, reproduction, and dormancy. The plant’s ability to respond to the environment enhances its survival. 10-35 Plant Tropisms Positive phototropism (stems bend toward the light) is due to the migration of auxin from the bright side to the shady side of a stem. After auxin arrives, the cells on that side elongate and the stem bends toward the light. In negative gravitropism, stems curve away from gravity due to auxin on the lower side of the stem. 10-36 Positive phototropism 10-37 Negative gravitropism 10-38 Flowering Photoperiod is the ratio light to darkness in a 24-hour cycle. Short-day (long night) plants flower when the days get shorter than a critical length and long-day (short night) plants flower when the days get longer than a critical length. Day-neutral plants do not depend on day length for flowering. 10-39 Effect of day/night length on flowering 10-40 Phytochrome and Plant Flowering If flowering is dependent on length of day and night, plants must have some way to detect photoperiod. Phytochrome is a plant pigment believed to be involved in regulating the response of plants to day length. Phytochrome exists in two forms, Pfr (active form) that is converted to Pr (inactive form) as night approaches; phytochrome conversion may signal day length. 10-41 Other Functions of Phytochrome The Pr → Pfr cycle also signals seeds when sunlight is present and conditions are good for germination, and it causes stems to etiolate. The Pfr binds to regulatory proteins in the cytoplasm and the complex migrates to the nucleus where it binds to specific genes. 10-42 Phytochrome control of a growth pattern 10-43 Transport in the Mature Plant Water and Mineral Transport in Xylem A plant uses active transport to concentrate minerals in root cells and xylem. Thereafter minerals are transported within water, in tracheids and vessel elements that form a pipeline to the leaves. Water entering root cells forms a positive root pressure. 10-44 Conducting cells of xylem 10-45 Cohesion-Tension Model The cohesion-tension model of xylem transport explains how water is transported to great heights against gravity. Polar water molecules are cohesive and adhere to the walls of the xylem vessel and fill the water pipeline. Transpiration, evaporation of water from leaves, creates a negative pressure that pulls the water column upward. 10-46 Cohesion-tension theory of xylem transport 10-47 Mineral Transport In addition to the carbon, hydrogen and oxygen obtained from water and carbon dioxide, a plant needs mineral elements. A plant uses active transport to take in minerals beyond what is provided by the flow of water. Plants are important for concentrating minerals that are used by consumers including humans. 10-48 Adaptations of Roots for Mineral Uptake Bacteria in the root nodules of legumes are symbionts that convert the nitrogen in the atmosphere to NH4+. Fungi have a symbiotic relationship with plant roots, called mycorrhizae, that increase water and mineral uptake and improve nutrient transfer. Plants without mycorrhizae grow in limited environments. 10-49 Root nodules 10-50 Mycorrhizae 10-51 Some plants have poorly developed or no roots and have minerals and water supplied by other means. Epiphytes (air plants) do not grow in soil and therefore must use roots to extract moisture from air and catch rain and minerals in leaves. Parasitic plants send out root-like haustoria that tap into the xylem and phloem of the host stem. A few carnivorous plants supplement their diet by capturing insects. 10-52 Opening and Closing of Stomata Guard cells on either side of a stoma regulate its opening and closing. When water enters guard cells, the stoma opens; when water leaves, it closes. When a plant is photosynthesizing, a pump actively transports hydrogen ions (H+) out of guard cells. Then potassium (K+) ions followed by water enter guard cells and the stoma opens. 10-53 When the pump is not working, K+ followed by water exits guard cells and the stoma closes. The blue-light component of sunlight appears to be absorbed by a pigment that leads to activation of the H+ pump. Abscisic acid also causes stomata to close. Stomata continue to open and close when kept in the dark, indicating an internal biological clock. 10-54 Opening and closing of stomata 10-55 Organic Nutrient Transport in Phloem Green parts of plants produce sucrose that is transported in phloem. Conducting cells are sieve-tube elements, accompanied by a companion cell and strands of cytoplasm called plasmodesmata. Transport of organic materials within phloem is termed translocation. 10-56 Sieve-tube elements of phloem 10-57 Pressure-Flow Model of Phloem Transport The pressure-flow model of phloem transport explains how organic nutrients are transported in a plant. During the growing season, the leaves are a source of sugar. Using energy from ATP, sugar is actively transported into sieve-tube elements and water follows passively. 10-58 Buildup of water creates a pressure and starts a flow of phloem sap toward a sink where sugar is actively transported out of sieve-tube elements. Because the direction that phloem sap flows is from the source to the sink, this explains transport to newly formed leaves or to fruits. 10-59 Pressure-flow theory of phloem transport 10-60 Chapter Summary In a plant life cycle, pollen grains carry sperm from flower to flower. Seeds within fruits are the products of sexual reproduction in flowering plants. Seeds must be dispersed and must germinate to complete the life cycle. 10-61 Plants reproduce asexually; this ability can be commercially utilized to mass produce identical plants, sometimes after genetic engineering. Plants respond to outside stimuli by changing their growth patterns. Plant hormones regulate plant growth patterns. Some plant responses are controlled by the length of daylight (photoperiod). 10-62 Transpiration pulls water and minerals from the roots to the leaves in xylem according to the cohesion-tension model. Stomates are open for evaporation to occur. The pressure-flow model of phloem transport states that sugar is actively transported into phloem at a source, and water follows by osmosis. The resulting increase in pressure creates a flow, which moves water and sucrose to a sink. 10-63