Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Plant nutrition wikipedia , lookup
History of herbalism wikipedia , lookup
Plant defense against herbivory wikipedia , lookup
Plant secondary metabolism wikipedia , lookup
History of botany wikipedia , lookup
Plant physiology wikipedia , lookup
Evolutionary history of plants wikipedia , lookup
Ecology of Banksia wikipedia , lookup
Historia Plantarum (Theophrastus) wikipedia , lookup
Ornamental bulbous plant wikipedia , lookup
Plant morphology wikipedia , lookup
Plant use of endophytic fungi in defense wikipedia , lookup
Plant evolutionary developmental biology wikipedia , lookup
Plant ecology wikipedia , lookup
Perovskia atriplicifolia wikipedia , lookup
Gartons Agricultural Plant Breeders wikipedia , lookup
Plant breeding wikipedia , lookup
Flowering plant wikipedia , lookup
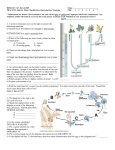
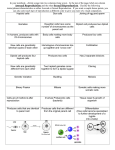
Ó The American Genetic Association. 2009. All rights reserved. For permissions, please email: [email protected]. Journal of Heredity 2010:101(1):119–123 doi:10.1093/jhered/esp071 Advance Access publication August 12, 2009 Ploidy-Mediated Reduced Segregation Facilitates Fixation of Heterozygosity in the Aromatic Grass, Cymbopogon martinii (Roxb.) UMESH C. LAVANIA, SARITA SRIVASTAVA, AND SESHU LAVANIA From the Department of Genetics and Plant Breeding, Central Institute of Medicinal and Aromatic Plants, Lucknow 226 015, Uttar Pradesh, India (U. C. Lavania and Srivastava); and the Department of Botany, University of Lucknow, Lucknow 226 007, Uttar Pradesh, India (S. Lavania). Address correspondence to U. C. Lavania at the address above, or e-mail: [email protected]. In most medicinal and aromatic plants, the vegetative tissue (e.g., roots, stems, leaves) is the source of the economic product. These plants are inherently heterozygous (natural allelic hybrids) and maintain their genetic makeup in nature by obligate vegetative propagation. Under seed cultivation, these plants incur population heterogeneity that reduces biomass and hampers product quality. Therefore, fixation of heterozygosity is vital for maintaining uniformity in quality of the economic product and quantity of biomass under seed cultivation. Although seed-grown clonal progenies identical to the mother plant can be obtained in certain plants that show an unusual breeding system called apomixis, such a breeding system is rare in medicinal and aromatic plants of economic value. Here we show an effective experimental strategy based on a polyploid model that facilitates fixation of heterozygosity in obligate asexual species owing to tetrasomic inheritance and low segregation in C1 progenies from highfertility C0 autopolyploids. Using an obligate asexual species of aromatic grass—Cymbopogon martinii, we demonstrated that progenitor diploids with distal chiasma localization and low chiasmate association in meiosis, when changed into tetraploids, entail high gametic/seed fertility reflected in high bivalent pairing and balanced anaphase segregation. Their seed progenies evince crop homogeneity owing to reduced segregation, indicating fixation of heterozygosity present in the source diploids. Because C. martinii could be maintained through obligate vegetative propagation, here is a unique opportunity to utilize the polyploid advantage through C1 seed progenies for commercial cultivation, as well as maintenance of original C0 stock for raising seeds without losing polyploid heterosis normally threatened in subsequent segregating progenies on account of aneuploidy and gametic instability. Key words: Cymbopogon, diploidized polyploid, fixation of heterozygosity, hybrid vigor, tetrasomic inheritance The apomictic breeding system, if realized to its full potential, could be quite rewarding for fixation of heterozygosity and heterosis breeding in crop plants. It is all the more important in inherently heterozygous obligate asexual economic plants where seed-based cultivation is desirable for commercial cultivation. Although, apomixis is known to be naturally occurring in some 400 wild species from approximately 40 plant families, including various grasses (Ozias-Atkins and van Dijk 2007), the apomictic breeding system is rare in medicinal and aromatic plants of economic value/crop plants. Findings of Ravi et al. (2008) on a recessive mutation ‘‘Dyad’’ that promotes equational division leading to 2n egg cells is an important milestone having far-reaching implications for hybrid fixation in crop plants. However, prospects of these findings have inherent limitations because few 2n egg cells are produced in those Dyad mutants. There are also problems in proper development of triploid seeds owing to endosperm nuclear balance. Additionally, the seeds produced are not always genetically balanced and have a mix of aneuploid variants. Therefore, in the absence of a proper apomictic breeding system, an alternative effective strategy that facilitates fixation of heterozygosity in obligate asexuals is needed. The autotetraploid tetrasomic mode of reduced genetic segregation could be a prospective remedy if we could obtain genetically balanced and high-fertility tetraploids. The C0 autopolyploids could be maintained vegetatively in obligate asexuals as a source stock for continual C1 seed production for cultivation. Based on diploid–tetraploid meiotic correlations, Lavania (1995) determined that progenitor diploids harboring distal chiasma localization, when changed to the tetraploid state, are most likely to give rise to high gametic/seed fertility consummated through high bivalent pairing and balanced anaphase segregation. Taking 119 Journal of Heredity 2010:101(1) Table 1. Mean meiotic chromosome associations (±standard error) and pollen fertility in the progenitor diploid (2) and corresponding autotetraploid (4) in Cymbopogon martinii (4 5 40) Quadrivalents Bivalents Ploidy level Ring Chain Trivalents Ring Rod Univalents Pollen fertility Diploid Tetraploid 2.4 ± 0.245 0.15 ± 0.031 0.45 ± 0.045 4.6 ± 0.236 5.2 ± 0.245 5.4 ± 0.248 8.1 ± 0.38 — 1.85 ± 0.245 98.4 ± 2.56 96.6 ± 2.33 SE, standard error. cues from this, we planned to elucidate if the ploidy intervention in obligate asexuals could help facilitate fixation of allelic heterozygosity of the source diploids in the seedraised progenies. Materials and Methods Plant Materials Obligate asexual aromatic grass species Cymbopogon martinii (Palmarosa, family Poaceae, 2n 5 20) was used as a model experimental system. This species can be maintained vegetatively but is grown by seeds under commercial cultivation, leading to crop heterogeneity. This species is found naturally occurring throughout the semiarid plains of Central India. It is highly cross-pollinated and is valued for its essential oil obtained from its aboveground plant biomass. A composite variety ‘‘PRC-1’’ of this species developed at the Central Institute of Medicinal and Aromatic Plants (Lucknow, India) by intermating selected clones in polycross nursery was used as a starting experimental material. Seed progenies from PRC-1 were scored for meiotic behavior and chiasmate association to isolate 4 morphologically different plant types harboring distal chiasma localization. Fast growing vegetative tillers of the 4 selected plant types were used to produce clonal autotetraploids by the colchicine immersion method. Seed progenies from the source diploid clone (i.e., S1 progenies) and from their corresponding C0 tetraploid clones (i.e., C1 progenies) were grown under field conditions and scored for simple exomorphological appearance to elucidate population homogeneity/heterogeneity from phenotypic appearance. Clearly distinguishable and variable exomorphological characters such as width, length, orientation and pigmentation of leaves, length and color of peduncle, bushy/ lax spikelet arrangement, spreading/erect growth habit, etc., were observed to determine gross plant population heterogeneity/homogeneity. Meiotic and Fertility Analysis Thirty well-analyzable cells at metaphase I and anaphase I from each of the 4 progenitor diploid clones and their derived autotetraploid clones were scored for meiotic chromosome associations and anaphase segregation. Pollen fertility was examined after the usual acetocarmine staining; the fully stained pollen grains were considered as fertile. Analysis of Meiotic Configurations and Estimation of Chiasmate Association Frequency In order to assess uniformity for comparison of meiotic configurations, the observed values were converted into frequencies. This could be done by dividing the observed values by the haploid chromosome number (i.e., 10 in this case); however, for estimating univalent frequencies, the observed values for univalents are correspondingly assigned to univalent pairs and to trivalents as accompanying univalents. The simple approach of Sybenga (1975), as detailed out in Lavania (1995) and Lavania et al. (2006), especially suitable for species with predominantly distal chiasma localization was followed, for the estimation of multivalent pairing ( f ), and chiasmate association frequencies of average long arms (a) and short arms (b). In the given equations, the symbols t, rq, cq, r, o, and u represent the frequency of trivalent þ accompanying univalents, ring quadrivalents, chain quadrivalents, ring bivalents, open bivalents, and independent univalent pairs, respectively. Letters a and b represent the weighted average of chiasmate (bound arm) association of average long arm and average short arm, respectively. The simplified equations used are Table 2. Frequencies of various meiotic configurations and their bound-arm chiasmate associations of the progenitor diploid (2) and corresponding autotetraploid (4) in Cymbopogon martinii (4 5 40) Quadrivalents Ploidy level Trivalents þ 2-arm Ring Chain accompanying Ring Open Univalent Multivalent pairing Long arm Short arm association (rq) (cq) univalents (t) (r) (o) pairs (u) frequency (f) association (a) association (b) (a þ b) Diploid — — Tetraploid 0.24 0.015 120 Bivalents — 0.045 0.46 0.54 0.26 0.405 — 0.035 — 0.279 1.00 1.22 0.46 0.24 1.46 1.46 Brief Communications Figure 1. Meiotic metaphase and anaphase in the diploid. 2 5 20 (A, C) and autotetraploid, 4 5 40 (B, D) of Cymbopogon martinii. Note complete bivalent pairing and balanced anaphase segregation in the diploid as well as in the tetraploid. For tetraploids: 2 Multivalent pairingðf Þ 5 ðt þ 2cq þ 4rqÞ =16rq a b 5 rq þ 1=2cq þ 1=4t þ r a þ b 2ab 5 1=2cq þ 1=4t þ o: For diploids: Frequency of ring bivalents ðrÞ 5 a b; Frequency of open bivalentsðoÞ 5 a þ b 2ab: Furthermore, the individual values of a and b in all cases could be deduced using the quadratic equation: qffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffi ða þ bÞ± ða þ bÞ2 4ab : a; b; respectively 5 2 Results estimated as frequencies and are given in Table 2. The representative meiotic configurations depicting high bivalent pairing and balanced anaphase segregation are shown in Figure 1. Plant population from S1 and C1 seed progenies representing source diploid and corresponding autotetraploid are shown in Figure 2, depicting morphological crop heterogeneity in the diploid but homogeneity in the tetraploid. The observations suggest that the weighted average of bound-arm association frequency of less than 2 in the progenitor diploid inculcate high bivalent pairing in the derived autotetraploids, as seen from the low frequency of bound-arm association of the 2 arms in the autotetraploids and also reduced multivalent pairing. This is accompanied by balanced anaphase segregation followed by high gametic fertility in the autotetraploids. Meiotic Association and Gametic Fertility Population Homogeneity versus Heterogeneity in Seed Progenies The C0 autotetraploids realized for all the 4 selected diploid clones evinced high (.95%) gamete fertility. Such a high fertility in C0 autotetraploids was realized from high bivalent pairing and balanced anaphase segregation. Data recorded on meiotic chromosome associations and gametic fertility for the diploid vis-à-vis the autotetraploid (for a representative clone no. 2) is given in Table 1. For uniformity in comparison for bound-arm chromosome associations and multivalent pairing, the observed values for various meiotic configurations were Different specimens of seed progenies grown from the selfed seeds obtained from the source diploid clone (S1 progeny) and its corresponding C0 clone (C1 progeny) are shown in Figure 2. The C1 seed progenies exhibit population homogeneity against the heterogeneity found in the diploid seed progenies (S1). This clearly suggests that the native allelic heterozygosity present in the progenitor diploids is maintained (fixed) in C1 seed progenies through ploidy-mediated reduced segregation. 121 Journal of Heredity 2010:101(1) Figure 2. Plant population from S1 and C1 seed progenies from source diploid and corresponding autotetraploid. (A) Representative phenotypic variability in the diploid. (B) S1 seed progenies from source diploid. (C) C1 seed progenies from the autotetraploid. Note morphological crop heterogeneity in the diploid but homogeneity in the tetraploid (depicting reduced segregation/fixation of native heterozygosity in the tetraploid). Discussion Global duplication events are very common in plant evolution (Adams and Wendel 2005) and the genomes of most extant angiosperms retain evidence of one or more ancient genome-wide duplications (Cui et al. 2006). Polyploidy is particularly widespread in angiosperms and, frequently, involves unreduced gametes and interspecific hybridization (Leitch AR and Leitch I 2008; Soltis PS and Soltis DE 2009). Whereas incidence of allopolyploidy is quite usual in plants, but autopolyploidy, although it does 122 occur in nature, is found only in low frequency due to maladaptive features associated with meiotic disturbances (Ramsey and Schemske 2002; Otto 2007; Soltis et al. 2007). Nevertheless, there are several instances where induced autotetraploids are known to exhibit balanced anaphase segregation leading to high gamete fertility and reasonably good seed set in C0. A more regular anaphase segregation in polyploids could be best achieved by increasing the frequency of bivalents at the expense of other configurations, as observed in natural tetraploids of Lotus corniculatus, Allium porum, and Dactylis glomerata (Lavania 1995). Weiss and Brief Communications Maluszynska (2000) observed almost complete bivalent formation in established autotetraploids of Arabidopsis attributing it to small size of the chromosomes and to a process of partial diploidization. Bivalent frequency was high in the autotetraploids on account of a low number of autonomous pairing sites (Santos et al. 2003)/low chiasmate association in diverse species (Lavania et al. 1991; Srivastava et al. 1992). An exhaustive account of quantitative genetic analysis of meiotic configurations in highly fertile autotetraploids compared with diploid progenitors, underpinned that progenitor diploids with distal chiasma localization, caused high bivalent pairing in polyploids and consequently balanced anaphase segregation and high gamete and seed fertility in derived polyploids (Lavania 1995). One of the possibilities of the enhanced bivalent pairing observed in the autoteraploid of Cymbopogon martini could be the gradual shift in the point of pairing partner exchange toward one of the chromosome ends during pairing just before chiasma formation. This is reflected in the enlarged differences observed in the estimated frequencies of bound associations in the 2 chromosome arms. When the point of partner exchange reaches the chromosome end in time, 2 bivalents are formed, and these may be observed either as ring or as rod (open) bivalents (Lavania et al. 1991). The almost complete bivalent pairing observed in the present case points to the possibility of obtaining high fertility in C0 tetraploids and the consequent reduced segregation/fixation of heterozygosity in C1 seed progenies. The cytological considerations mentioned above could serve as possible basis for genotypic selection of prospective diploid progenitors to develop high-fertility autotetraploids. These findings have application in plant breeding where seeds are required in cultivation. In addition to harnessing the polyploid advantage per se in crop improvement, this approach offers the opportunity of hybrid fixation in obligate asexuals because the source C0 autotetraploids could be maintained vegetatively and used as genetic stock for continual seed production. The tetrasomic segregation pattern would help maintain allelic hybridity from the source material to a large extent (.95%) in C1 progenies in genetically balanced high fertility autopolyploids, as demonstrated here. References Adams KL, Wendel JF. 2005. Polyploidy and genome evolution in plants. Curr Opin Plant Biol. 8:135–141. Cui L, Wall PK, Leebens-Mack JH, Lindsay BG, Soltis DE, Doyle JJ, Soltis PS, Carlson JE, Arumuganathan K, Barakat A, et al. 2006. Wide spread genome duplications throughout the history of flowering plants. Genome Res. 16:738–749. Lavania UC. 1995. Genesis of high bivalent pairing in autotetraploids and autotriploids, and reduction in bound arm associations over generations. In: Brandham PE, Bennett MD, editors. Kew Chromosome Conference IV. Kew (UK): Royal Botanic Gardens. p. 397–406. Lavania UC, Srivastava S, Lavania S. 2006. High bivalent frequencies in the autotetraploids of aromatic grass, Cymbopogon martinii (Roxb.) Wats. facilitates polyploid stability. In: Bang J-W, Mukai Y, editors. Advances chromosome science 2. Daejeon (Korea): Daeheung Press Inc. p. 9–18. Lavania UC, Srivastava S, Sybenga J. 1991. Cytogenetics of fertility improvement in artificial autotetraploids of Hyoscyamus niger L. Genome. 34:190–194. Leitch AR, Leitch I. 2008. Genomic plasticity and the diversity of polyploid plants. Science. 320:481–483. Otto SP. 2007. The evolutionary consequences of polyploidy. Cell. 131:452–462. Ozias-Atkins P, van Dijk PJ. 2007. Mendelian genetics of apomixis in plants. Annu Rev Genet. 41:509–537. Ramsey J, Schemske DW. 2002. Neopolyploidy in flowering plants. Annu Rev Ecol Syst. 33:589–639. Ravi M, Marimuthu MPA, Siddiqi I. 2008. Gamete formation without meiosis in Arabidopsis. Nature. 451:1121–1124. Santos JL, Alfaro D, Sanchez-Moran E, Armstrong SJ, Franklin FCH, Jones GH. 2003. Partial diploidization of meiosis in autotetraploid Arabidopsis thaliana. Genetics. 165:1533–1540. Soltis DE, Soltis PS, Schemske DW, Hancock JF, Thompson JN, Husband BC, Judd WS. 2007. Autopolyploidy in angiosperms: have we grossly underestimated the number of species? Taxon. 56:13–30. Soltis PS, Soltis DE. 2009. The role of hybridization in plant speciation. Annu Rev Plant Biol. 60:561–588. Srivastava S, Lavania UC, Sybenga J. 1992. Genetic variation in meiotic behaviour and fertility in tetraploid Hyoscyamus muticus: correlation with diploid meiosis. Heredity. 68:231–239. Sybenga J. 1975. Meiotic configurations. Berlin (Germany): Springer-Verlag. Weiss H, Maluszynska J. 2000. Chromosomal rearrangement in autotetraploid plants of Arabidopsis thaliana. Hereditas. 133:255–261. Funding Received January 28, 2009; Revised June 25, 2009; Accepted July 20, 2009 New Idea Fund Scheme of CSIR, New Delhi (to U.C.L.). Corresponding Editor: Prem Jauhar 123