Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
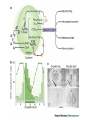
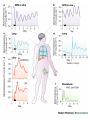
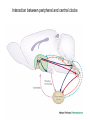
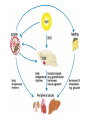
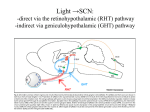
The Big PICTURE: influence of the SCN timing system on various functions From Zigmond et al Fundamental Neuroscience, AP 1999 Individual neurons dissociated from rat suprachiasmatic nucleus express independently phased circadian firing rhythms Neuron 14: 697-706, 1995 DK Welsh, DE Logothetis, M Meister and SM Reppert • Within the mammalian hypothalamus, the suprachiasmatic nucleus (SCN) contains a circadian clock for timing of diverse neuronal, endocrine, and behavioral rhythms. • By culturing cells from neonatal rat SCN on fixed microelectrode arrays, we have been able to record spontaneous action potentials from individual SCN neurons for days or weeks, revealing prominent circadian rhythms in firing rate. • Despite abundant functional synapses, circadian rhythms expressed by neurons in the same culture are not synchronized. • After reversible blockade of neuronal firing lasting 2.5 days, circadian firing rhythms re-emerge with unaltered phases. • These data suggest that the SCN contains a large population of autonomous, single-cell circadian oscillators, and that synapses formed in vitro are neither necessary for operation of these oscillators nor sufficient for synchronizing them. Electrical synapses coordinate activity in the suprachiasmatic nucleus MA Long, MJ Jutras, BW Connors, RD Burwell Nature Neuroscience 8, 61 - 66 (2004) • In the suprachiasmatic nucleus (SCN), the master circadian pacemaker, neurons show circadian variations in firing frequency. There is also considerable synchrony of spiking across SCN neurons on a scale of milliseconds, but the mechanisms are poorly understood. • Using paired whole-cell recordings, we have found that many neurons in the rat SCN communicate via electrical synapses. Spontaneous spiking was often synchronized in pairs of electrically coupled neurons, and the degree of this synchrony could be predicted from the magnitude of coupling. • In wild-type mice, as in rats, the SCN contained electrical synapses, but electrical synapses were absent in connexin36-knockout mice. The knockout mice also showed dampened circadian activity rhythms and a delayed onset of activity during transition to constant darkness. • We suggest that electrical synapses in the SCN help to synchronize its spiking activity, and that such synchrony is necessary for normal circadian behavior. Electrical coupling in SCN- the data • (a) In a well-coupled pair (coupling coefficient = 0.13), hyperpolarizing current steps resulted in symmetric membrane-potential deflections in the coupled cell. In the left panel, we injected current into cell 1 and recording voltage responses in that cell and its pair. In the right panel, current was injected into cell 2. Traces are averaged from 50 trials. (b) Single action-potential response in a coupled pair in the presence of APV, DNQX and picrotoxin. Shown here is the mean postsynaptic response to 25 spontaneously occurring action potentials (dashed lines = s.e.m.). (c) Scatterplot of the coupling strength of SCN cell pairs as a function of the time of day recorded. Note the greater incidence of coupling in the middle of the subjective day (ZT 4−8) as compared the end of the subjective day (ZT 8−12). Coupled vs non-coupled SCN cells • All experiments were conducted in the presence of APV, DNQX and picrotoxin. (a) Examples of intracellular recordings from a pair showing direct electrical coupling (left) and a pair lacking such a connection (right). Below are spike cross-correlograms describing long epochs (>200 s) of spiking data, including the above traces. The time scale for the cross-correlograms is in milliseconds. Correlogram values are normalized by the total spike count. Baseline correlations are subtracted from these analyses. (b) Examples of cell-attached recordings (top) from a coupled (left) and noncoupled (right) pair, and the corresponding spike cross-correlograms (below). (c) The spiking correlation coefficient of electrically coupled pairs is directly related to the strength of coupling. This scatterplot shows data from cellattached recordings ( ) as well as intracellular recordings ( Electrophysiology and circadian behavior in wild-type (WT) and Cx36-knockout (KO) mice. Bridging the gap: coupling single-cell oscillators in the suprachiasmatic nucleus CS Colwell Nature Neuroscience 8, 10 - 12 (2005) • Top, schematic of pairs of SCN neurons (blue) from wild-type (WT) and Cx36-/- mice. Individual SCN neurons contain the molecular machinery necessary to generate circadian oscillations. One gap in our knowledge is the lack of understanding of how these single-cell oscillators are coupled. The new study3 demonstrates that SCN neurons are coupled through direct electrical connections. This coupling is lost in mice deficient in Cx36. Bottom, schematics of wheel-running activity records from WT and Cx36-deficient mice. Animals maintained in constant darkness show rhythms driven by the endogenous timing system. Each horizontal row represents the activity record for a 24-hour day. Successive days are plotted from top to bottom. The colored bars represent activity. The WT mice express robust circadian rhythms of locomotor activity with period shorter then 24 h. The onset of activity is typically under precise control. In contrast, the Cx36-deficient mice showed rhythms that were weaker and less coherent than controls. Without the Cx36, the circadian clock still keeps time but lacks the temporal precision that typically characterizes the behavioral output. A CLOCKWORK WEB: CIRCADIAN TIMING IN BRAIN AND PERIPHERY, IN HEALTH AND DISEASE Michael H. Hastings, Akhilesh B. Reddy & Elizabeth S. Maywood Nature Neuroscience Reviews August 2003 Time after time: inputs to and outputs from the mammalian circadian oscillators David Morsea, Paolo Sassone-Corsi TINS 25: 632-637, 2002 • • • • Fig. 1: Rhythmic oscillation of clock gene expression. (a) The mammalian circadian system has two negative feedback loops, one involving inhibition of bmal1 transcription by a heterodimer of BMAL1 (brain–muscle Arnt-like protein 1) and CLOCK (left), the other involving inhibition of period ( per) and cryptochrome ( cry) transcription by PER and CRY (right). The two loops are linked by the positive action of PER on bmal1 transcription and the positive action of the CLOCK–BMAL1 heterodimer (indicated by the yellow box) on E-boxcontaining promoters. Grey parts of the genes represent promoters; wavy lines indicate that levels of bmal1, per and cry transcripts oscillate. (b) In the mouse suprachiasmatic nucleus (SCN), per and bmal1 transcripts oscillate almost 12 h out of phase, with per levels at a maximum around midday. PER and BMAL1 proteins also oscillate, but PER lags behind per RNA. (c) In peripheral tissues, such as the liver, the same clock gene components oscillate as in the SCN, but with peaks occurring later. The black and white bars above the graphs indicate night and day, respectively. Nature Reviews Neuroscience 2, 521-526 (2001); HYPOTHALAMIC INTEGRATION OF CENTRAL AND PERIPHERAL CLOCKS Ruud M. Buijs & Andries Kalsbeek • During sleep, our biological clock prepares us for the forthcoming period of activity by controlling the release of hormones and the activity of the autonomic nervous system. Here, we review the history of the study of circadian rhythms and highlight recent observations indicating that the same mechanisms that govern our central clock might be at work in the cells of peripheral organs. Peripheral clocks are proposed to synchronize the activity of the organ, enhancing the functional message of the central clock. We speculate that peripheral visceral information is then fed back to the same brain areas that are directly controlled by the central clock. Both clock mechanisms are proposed to have a complementary function in the organization of behaviour and hormone secretion. Interaction between peripheral and central clocks The Circadian Gene Period2 Plays an Important Role in Tumor Suppression and DNA Damage Response In Vivo Fu, L et al. Cell 111: 41-50, 2002 • The Period2 gene plays a key role in controlling circadian rhythm in mice. We report here that mice deficient in the mPer2 gene are cancer prone. After γ radiation, these mice show a marked increase in tumor development and reduced apoptosis in thymocytes. The core circadian genes are induced by γ radiation in wild-type mice but not in mPer2 mutant mice. Temporal expression of genes involved in cell cycle regulation and tumor suppression, such as Cyclin D1, Cyclin A, Mdm-2, and Gadd45α, is deregulated in mPer2 mutant mice. In particular, the transcription of c-myc is controlled directly by circadian regulators and is deregulated in the mPer2 mutant. Our studies suggest that the mPer2 gene functions in tumor suppression by regulating DNA damage-responsive pathways. Drosophila melanogaster