Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
Hearing Research, 38 (1989) 163-176 Elsevier HRR 163 01181 Response from the exposed intracranial human auditory nerve to low-frequency tones: Basic characteristics Aage R. Mprllerand Hae Dong Jho Department of Neurological Surgery, University of Pittsburgh School of Medicine, Pittsburgh, Pennsylvania, U.S.A. (Received 5 June 1988; accepted 3 November 1988) The responses recorded from the exposed intracranial portion of the eighth nerve in man with normal hearing to short bursts of low-frequency tones (500, 1000, and 1500 Hz) consist of two components; these two components can be separated by adding and subtracting, respectively, the responses to tonebursts of opposite polarity. Subtracting the responses to tones of opposite polarity reveals a waveform that resembles the sinusoidal waveform of the stimulus (frequency-following response = FFR), while adding the responses to tones of opposite polarity reveals a slow component, the waveform of which is more variable than the frequency-following component. The initial deflection of the slow component of the response to 1000 Hz and to 1500 Hz is a positive peak followed by a slow, negative deflection, and the response to 1500-Hz tonebursts often shows a clear off-response. The slow component of the response to 500-Hz tones often has an initial negative peak followed by a slow, positive or negative wave. The temporal relationship between the stimulus tone and the frequency-following component changes only slightly when the intensity of the sound is changed, whereas the latency of the slow potential decreases with increasing stimulus intensity. The FFR can be masked by noise, and the results of masking with highpass-filtered noise indicate that the frequency-following response may be generated at a location on the basilar membrane that is tuned to a frequency that is higher than that of the stimulus tone. Human auditory nerve; Frequency-following response; Intraoperative Introduction Goldstein and Kiang (1958) showed that the gross response that can be recorded from the auditory nerve is related to the discharges in individual nerve fibers. The former is the convolution between the distribution of neural discharges over the population of nerve fibers that contribute to the response and the waveform of a single nerve impulse. Then, in 1962, Teas et al. presented results that were fundamental to the interpretation of auditory nerve action potentials and the distribution of excitation along the basilar membrane in the guinea pig. They used selective masking with narrowband noise of the response to broad- Correspondence to: Aage R. Moller, Dept. of Neurological Surgery, Room 9402, Presbyterian-University Hospital, DeSoto at O’Hara Streets, Pittsburgh, PA 15213, U.S.A. 0378-5955/89/$03.50 0 1989 Elsevier Science Publishers recordings band clicks, to define areas of excitation along the basilar membrane. It was earlier assumed that it was only the rarefaction phase of a sound, corresponding to a deflection of the basilar membrane towards the Scala vestibuli, that was excitatory. More recently, it has been shown that the neural excitation that occurs in the cochlea is far more complex. Thus, Konishi and Nielsen (1973, 1978), Zwislocki and Sokolich (1973), Zwislocki (1974), and Sokolich et al. (1974) found that, when recording from single auditory nerve fibers using sounds with sinusoidal and trapezoidal waveforms, the motion of the basilar membrane towards the Scala tympani was excitatory in the majority of nerve fibers. Some fibers responded to basilar membrane velocity, depending on the frequency to which the nerve fiber is tuned (CF) and on the frequency and intensity of the sound. Since basilar membrane displacement is related to stapes velocity, the B.V. (Biomedical Division) 164 majority of nerve fibers will respond to the derivative of the condensation phase of low-frequency sounds. The timing (phase angle) of excitation of single auditory nerve fibers in response to pure tones shows a transition in the mid CF range, and there is thus a low and high range of CF where the phase angle of excitation shows little change as a function of CF for low stimulus frequencies. In between these two ranges the phase of excitation changes rapidly as a function of its CF (about 540 degrees/octave). Fibers with a CF that is higher than twice that of the stimulus tone change slowly as a function of their CF (about 90 degrees/octave) (Rim et al., 1979; Ruggero and Rich, 1983, 1987; Ruggero et al., 1986). Since the phase-locked component of the gross responses from the auditory nerve to lowfrequency tones of moderate to high intensity can be assumed to be the vector sum of the phaselocked activity in many single auditory nerve fibers, the responses from fibers with CF below twice the frequency of the stimulus tones are probably unlikely to contribute significantly to the gross response, which therefore can be assumed to be a result of the phase-locked activity in fibers with CF above twice the frequency of the tone. Only the phase-locked response to low-intensity tones (near threshold) may be assumed to originate near the location of maximal deflection on the basilar membrane. This was in fact confirmed by Snyder and Schreiner (1985) in experiments in cats. These investigators used forward masking to show that it is only tones near threshold that originate from a region of the basilar membrane that is tuned to the frequency of the tone, and that the location on the basilar membrane which contributes maximally to the phase-locked gross response shifts towards the base of the cochlea as the intensity of the tone is increased. The response to loud tones originates from regions on the basilar membrane that are tuned to frequencies that are about twice the frequency of the stimulus tones (Snyder and Schreiner, 1985). In interpreting these results it must also be taken into account that the width of the tuning increases as the sound intensity is increased (Rhode, 1971, 1978; Sellick et al., 1982b). Recordings from single auditory nerve fibers show a similar widening of tuning when broadband noise is used as the test signal, and there is a shift in the CF that indicates that the location of maximal excitation on the basilar membrane shifts towards the base of the cochlea with increasing stimulus intensity (Moller, 1977, 1983). These results obtained in animals made it seem likely that information about the properties of human neural discharges could be obtained from recordings of gross responses from the human auditory nerve. This would be advantageous, as it is usually not possible to obtain recordings from single units of the human auditory nerve, but it is relatively easy to obtain recordings of gross potentials. The purpose of this study was to provide information about the human ear and auditory nerve that may not be directly inferred from the results of studies in which experimental animals such as the cat are used. Because of possible differences in the cochleas of man and such small animals, e.g., the auditory nerve is much longer in man (Lang, 1981) than it is in such animals, the results of animal studies are not always applicable to man. Studying the responses recorded from the human auditory nerve to tones may permit correlation of physiological measurements with psychoacoustic measurements made in patients with different types of sensorineural hearing loss. Thus, this report presents results of recording from the intracranial portion of the human eighth nerve in response to low-frequency tones. Patients and Methods Recordings were made from the exposed eighth nerves of patients undergoing neurosurgical operations to relieve trigeminal neuralgia (TN), hemifacial spasm (HFS), or disabling positional vertigo (DPV). The technique used was microvascular decompression (MVD) of the respective cranial nerve(s) (see Jannetta, 1981a,b). The patients who were operated upon to relieve TN and DPV were anesthetized using a balanced anesthesia regimen with a strong analgesic (Fentanyl) and a muscle relaxant (Pavulon) together with nitrous oxide. Patients operated upon to relieve HFS were anesthetized using inhalation agents (isoflurane 165 and nitrous oxide) and small amounts of Fentanyl, but no long-lasting muscle relaxants. The stimulus sounds consisted of bursts of pure tones at frequencies of 500, 1000, and 1500 Hz, with a plateau of 10 ms and a rise and fall time of two periods. Clicks were generated by applying rectangular pulses of 100~ps duration at a repetition rate of 19 or 9 pulses per s (Grass Instrument Co., Type SlO CTCMA) using miniature stereo earphones (Radio Shack, Realistic) that were sealed in the patient’s ear with adhesive tape. The masking noise was generated by the same audiostimulator and was added to the clicks and tonebursts after being highpass filtered (roll-off of 54 dB/octave). The characteristic of the sound system was determined by attaching the earphone to a 2-0~ cavity partly filled with cotton, and measuring the sound pressure with a calibrated l/Cinch condenser earphone (Brtiel and Kjaer, Type 4134). The potentials were recorded from the exposed eighth nerve using a monopolar electrode made from a fine, malleable, Teflon-insulated, multistrand, silver wire with a small, cotton wick sutured to its uninsulated tip (Moller and Jannetta, 1983). A noncephalic reference electrode (Grass Instrument Co., needle electrode Type E2) was placed on the shoulder or on the contralateral earlobe. A similar needle electrode placed over the sternum served as a ground electrode. The potentials were amplified using differential amplifiers (Grass Instrument Co., Type 12) with a gain of 20,000 times and filter settings of 3 to 3000 Hz (roll-off of 6 dB/octave). After additional lowpass filtering (3.4kHz cut-off and a roll-off of 18 dB/octave), the potentials were averaged using an LSI 11/73 microprocessor with an analog-to-digital converter with 1Zbit resolution. Fifty to 200 responses of the tones were averaged and stored on computer disks, after which the tones were inverted’ and a similar number of responses were averaged and stored. The responses to opposite polarities of the stimuli were added or subtracted to separate the FFR from the slow response. In some cases digital filtering was used to remove noise from the FFR. These filters were zero-phase, bandpass filters that were implemented by convolving the response with a weighting function that consisted of 3/2 sine waves with a frequency equal to that of the stimulus tones. The bandwidths (at 3 dB points) of these filters were 360 Hz at 500 Hz and 700 Hz at 1000 Hz. Brainstem auditory evoked potentials (BAEP) were recorded during the operation using platinum needle electrodes (Grass Instrument Co., Type E2) placed on the vertex and the ipsilateral earlobe. These potentials were amplified and processed in the same way as were the potentials recorded directly from the eighth nerve. The recording that was made before the operation but after the patient was anesthetized served as the baseline recording. Comparison of the potentials that were recorded during the operation to that of the baseline recording was made to ascertain that the auditory nerve had not been injured before the responses to low-frequency tones had been recorded from the nerve. The recordings presented in this report were made as part of the routine intraoperative monitoring of auditory evoked potentials that is done in patients in our institution. Such monitoring minimizes the risk of hearing loss due to manipulation of the eighth nerve. The research is done with patients’ informed consent according to a procedure approved by the Institutional Review Board of the University of Pittsburgh School of Medicine. ReSUltS The results of the present study are based on recordings made in 8 patients with normal hearing. The responses that can be recorded from the exposed eighth nerve in man to bursts of 500-Hz pure tones typically show an initial positive peak followed by a pattern of negative peaks (Fig. 1). Inverting the phase of the tone (Fig. 1, bottom tracings) results in the peaks in the response being shifted one-half wave of the sound; this indicates that it is predominantly one phase of the sound that gives rise to the peaks in the response. In order to extract the components of the responses to low-frequency tones that are related to the envelope of the sound (slow potential) from. those that are related to the waveform of the sound we added and subtracted, respectively, the responses to opposite phases of the tones. When the response to tone is subtracted from the re- 166 sponse to the same tone presented with reversed phase, a nearly sinusoidal pattern with the same frequency as the stimulus tone is seen; this phaselocked part of the response lasts as long as the toneburst (Fig. 2). We have chosen to call this part of the response the ‘frequency-follo~ng response’ (FFR). However, the same term was used by Worden and Marsh (1968) and Moushegian et al. (1978), as well as others, to describe the entire neural response to low-frequency tones (in contrast to cochlear microphonic (CM) potentials). When the responses to tones of opposite phase are added, the result is a slow potential that may be regarded to reflect the response to the envelope SOUND I ’ : , ’ ., 1: ‘/ ’ ‘, ‘, ,’ : I I I I I 0 5 10 15 20 i , 25 ms Fig. 2. Responses to 500-Hz tonebursts of both polarities superimposed (top tracing), together with the sums and differences (middle and lower tracings), to show the separation of the two components of the response. The sound intensity was 110 dB SPL, and the recordings were obtained in a patient who was undergoing an operation for DPV. ,‘, , DIFFERENCE I RESPONSE i : ‘, ; i ,, ,\, ‘, : ,,$,: it:!/ ’ I ,’ ( ‘, 1 ,, :’ e’ ! ,8 / ,’ ’ , ’ : : / ‘s, : ‘, ‘, I : ; ,’ ,,_ I I I I 0 5 10 15 I 20 Ins Fig. 1. Responses recorded from the exposed etghth nerve to MO-Hz tonebursts of both polarities in a patient who was operated upon for trigeminal neuralgia. The sound pressure, delivered by the earphone and measured in a slightly damped ~-CCcavity, was 113 dB SPL. The waveform of the stimulus sound is shown below each recordii with rarefaction as upward deflections. The same convention is used in all subsequent recordings. Negative potentials are shown as upward deflections in this and all subsequent figures. of the sound. The FFR is largely cancelled as the result of this procedure. The amplitude of the FFR decreases with increasing frequency of the tones (Fig. 3,A), and the waveform of the slow component of the response is often different for different frequencies of the stimulus tones. Thus, the slow component of the response to 1000 Hz and 1500 Hz tones typically has an initial positive deflection followed by a slow negativity that lasts as long as the sound, as may be seen in Fig. 3,B. The response to 1500-I& tones, in addition, has a clear off-response. The response to 500 Hz is often different in that it frequently has an initial negative peak followed by a slow wave, and its amplitude is usually smaller than that of the response to 1000 and 1500 Hz (Fig. 3,B). It may also be seen from Fig. 3 that the latency of the onset of the slow potential, as well as that 167 B DIFFERENCE 500 500 Hz Hz 4 1500 1000 Hz 1500 Hz H> -d CLICK I 3 15pv I I I I I 0 5 10 15 20 Ins 0 I I I 5 10 15 I 20 ms Fig. 3. Differences (A) and sums (B) of the responses to tonebursts of opposite polarities. Responses to tonebursts at 500, 1000, and 1500 Hz are shown for another patient who underwent an operation to relieve DPV. The sound levels were 104, 95, and 87 dB SPL for the three frequencies. The lower curves show the responses to click stimulation (dashed lines represent rarefaction clicks and solid lines are the responses to condensation clicks). of the FFR, is longest for the response to 500 Hz and shortest for the response to 1500 Hz, but the latency of the response to tones is longer than the latencies of the responses to click sounds. These latter responses are shown both for rarefaction (dashed lines) and condensation (solid lines) clicks presented at a peak SPL of 115 dB. Results of a more detailed analysis of the latencies of the FFR component will be given later in this paper. The amplitudes of both the FFR and the slow components of the response decrease as the stimulus intensity is decreased. However, the slow potential in response to lOOO-Hz tones can usually be discerned at lower sound intensities than the FFR (Fig. 4), while the slow component of the response to 500 Hz is often poorly developed (Fig. 5). The latency of the FFR in the range of relatively high stimulus intensities used is nearly independent of the stimulus intensity, but the latency of the initial positive peak of the slow component increases when the stimulus intensity is decreased. It may also be seen from Fig. 4 that the amplitude of the individual peaks of the response at the highest sound intensity is largest in the beginning of the toneburst, after which it decreases, indicating adaptation of the phaselocked response. The FFR component of the response to 500-Hz tonebursts at different sound intensities also shows that the latency is independent of the stimulus intensity in the intensity range that was studied Fig. 4. The FFR component (A) and the slow component (B) of the response to lC@O-Hz tonebursts at different intensities (given in dB SPL). The results were obtained in a patient who was undergoing a MM, operation for HFS. (Fig. 5), but the slow component is much less prominent. When the sound level is decreased, the slow component becomes indiscernible when the FFR still is clearly visible. The responses to low-frequency sounds of high intensities have distorted waveforms: peaks appear between the main peaks of the response (Fig. 6). Such distortion products are cancelled when the responses to tones of opposite polarities are subtracted to obtain the FFR, but the distortion can be observed when the responses to stimuli of opposite polarity are added: at this time it is seen as a ripple superimposed on the slow potential (Fig. 5,B). The response to 500-Hz tones usually shows a higher degree of distortion than the response to lOO@Hz and 1500-Hz tones. However, there is considerable variation from person to person. The traditional way of determining the latency of a response is to measure the time between the beginning of the sound and the beginning of the response. Since the sound must have a smooth onset to avoid transients, this method may be inaccurate when low-frequency tones are used, so we determined the latency of the FFR in a different way as well, namely, by measuring how much the waveform of the sound needed to be shifted in time to match the waveform of the response (Fig. 7). To do this, we assumed that it is the rarefaction phase of the sound that results in neural excitation, and that neural excitation produces a negative nerve potential. In turn, the assumption 169 B A DIFFERENCE 104 dB 104 il dB 94 dB 94 dB 84 dB 84 dB 74 dB 74 dB SOUND I 0 I I I 5 10 15 I 20 Ins I 0 I I I I 5 10 15 20 ms Fig. 5. Differences (A) and sum (B) of the responses to 500 Hz shown in a way similar to that in Fig. 4 to 500-I-k tones. that neural excitation produces a negative potential on the surface of the nerve presumes that the recording is made from a position on the nerve where a propagated wave of nerve activity passes under the electrode. This can be achieved by recording from the eighth nerve at a distance from its entry into the brainstem (as long as the nerve is not injured so that neural transmission is blocked). The latency of the FFR to 500-Hz and lOOO-Hz tones measured in that way was 3.2 and 2.2 ms, respectively, in the patient whose responses are shown in Fig. 7. This should be compared with the latency of the main negative peak in the CAP, recorded in response to rarefaction clicks immediately before the response to tonebursts was obtained; the latency of the main negative peak in the CAP to clicks was 3.3 ms in the patient depicted in Fig. 7. The time difference between the second rarefaction wave of the sound (as measured in a calibration coupler) and the second negative peak in the response to 500-Hz and lOOO-Hz tonebursts in the 8 patients in this study was slightly longer for the 500-Hz tonebursts than for the 1000~Hz tonebursts (Table I). The latency when measured in this way is a measure of how much the sound waves had to be shifted to obtain the best match between the waveform of the responses and that of the tonebursts (c.f., Fig. 7). The latency of the negative peak in the CAP in response to high-intensity rarefaction clicks was slightly longer than the latency for the response to lOOO-Hz tones. There was, however, considerable individual variation. The assumption that the responses to the lowfrequency tones that we studied are generated by 94 de ‘, ! ’ , (‘,j’, , 1 ‘, ,’ I 5vv reduced to about half its amplitude when the cut-off frequency of the masking noise is 1800 Hz (spectrum level of 60 dB SPL/Hz). When the cut-off frequency is increased to 3300 Hz the effect on the FFR is minimal. Recordings from another patient show that masking with noise with a cut-off frequency of 1900 Hz (60 dB spectrum level) almost eliminates the response to lOOO-Hz tones at 95 dB SPL, and noise with a cut-off frequency of 2800 Hz, and the same spectrum level causes a marked reduction in the response to the same lOOO-Hz tones. In the same patient, noise with a cut-off frequency of 1200 Hz reduces the response to 500-Hz tones (104 dB SPL) by about 50% (Fig. 9,B). There is no noticeable effect on the timing of the FFR as a result of masking. The effect from masking on the slow component of the response is more difficult to assess, because this component of the response is more variable than the FFR. SOUND Discussion Fig. 6. Unprocessed responses to SOO-Hz tones of both polarities at two different stimulus intensities to show how distortion increases with increasing stimulus intensity. The data are the same as shown in Fig. 5. The results of the present study, obtained by monopolar recording from the eighth nerve in man, are essentially similar to those obtained by Snyder and Schreiner (1984, 1985) by recording from cats’ eighth nerves using bipolar electrodes. TABLE the intracranial portion of the audito& nerve is supported by the finding that the latencies of the responses changed when the recording electrode was moved along the intracranial portion of the eighth nerve: as can be seen from Fig. 8, the latency was shorter when the recording electrode was placed more distally compared to when it was placed more proximally. Further, the latency of the response to click sounds changed slightly less than did the latency of the FFR. Simultaneous masking with continuous highpass-filtered noise reduces the amplitude of the FFR in response to low-frequency tones when the cut-off frequency of the noise is lower than about twice the frequency of the stimulus sound (Fig. 9,A). The FFR to lOOO-Hz tones at 95 dB SPL is I TIME DIFFERENCE BETWEEN THE SECOND RAREFACTION WAVE IN THE SOUND AND THE SECOND NEGATIVE DEFLECTION IN THE RESPONSE TO TONEBURSTS, TOGETHER WITH THE LATENCY OF THE NEGATIVE PEAK IN THE RESPONSE TO RAREFACTION CLICKS Patient 500 Hz 1000 Hz N, 1. 2. 3. 4. 5. 6. 7. 8. 2.92 3.32 3.20 2.82 3.72 3.72 3.62 3.30 2.72 3.00 2.85 2.80 2.45 2.20 2.44 3.08 3.12 2.90 3.20 2.92 3.00 3.20 Mean S.D. 3.33 0.32 2.67 0.27 2.98 0.23 171 I I I I I I 0 5 10 15 20 25 In* I I I I I 0 5 10 15 20 I 25 m* Fig. 7. Responses to 500-Hz (left) and 1000-Hz (right) tonebursts of both polarities (A,B), together with the dtiierencc between these recordings (FFR) (C) (solid lines). The waveform of the stimulus sound, after the sound was shifted 3.2 and 2.2 ms, respectively (dashed lines), is also shown to demonstrate how latency of the FFR is measured. Below (D) is the response to broadband clicks. The results were obtained in a patient who was operated upon to relieve trigeminal neuralgia (same patient as illustrated in Fig. 1). The effects on the response of masking with continuous highpass-filtered noise that we see in man are similar to those described by Snyder and Schreiner (1985) using forward mashing in the cat. Thus, our results support these investigators’ findings that the FFR are nerve potentials and that also, in man, the FFR is generated by nerve fibers that are tuned to frequencies higher than those of the stimulus tones. That the timing of the FFR is relatively independent of stimulus intensity, while the latency of the slow potential decreases with increasing stimulus intensity, indicates that the two components of the response, the FFR and the slow potential, may not have the same origin. The latter seems to be dependent on a synaptic build-up of excitatory postsynaptic potentials (or generator potentials), which occurs at a slower rate when the stimulus intensity is low than when the stimulus intensity is high. At the relatively high stimulus levels used in this study, the timing of the FFR is largely independent of the stimulus intensity. Similar results were obtained when utilizing crosscorrelograms of the responses to continuous amplitude-modulated noise (Moller, 1981a,b). The difference (0.66 ms) in the delays of the responses to 500 Hz and to 1000 Hz may be taken as a measure of the difference in traveling time on the basilar membrane from which the responses to these two sounds originate (probably locations that are tuned to 1000 and 2000 Hz). The method used to measure the latency of the FFR is based on the assumption that it is the rarefaction phase of a sound that is excitatory. If instead it was the condensation phase that was excitatory, then the latency measurements would 172 2PV I 0 I I 5 I ,o I 15 I 20 ms Fig. 8. FFR recorded from two different locations on the eighth nerve: near the porus acousticus (distal), and at about the middle of the intracranial portion of the nerve. The CAP recorded from these two locations on the nerve in response to click sounds are also shown. The results were obtained in,a patient who was undergoing an MVD operation for disabling positional vertigo (same patient as illustrated in Fig. 3). be in error by the duration of one-half period of the sound. In the same way, if it is the velocity of a sound that is excitatory, then the error would be one quarter of a wave or three quarters of the duration of a wave. The fact that the FFR response is well defined makes it unlikely that the match between the sound wave and the FFR component of the response should be in error by a full period of the sound wave due to uncertainty in the determination of the beginning and the end of the FFR response. The phase shift of the middle ear is not taken into account when the reference for comparing the FFR with the stimulus is based on the sound pressure that is measured in a calibration cavity. If the timing of the FFR is to be compared with the timing of the motion of the cochlear fluid, then that phase shift must be taken into account. The phase shift in the middle ear is a function of the frequency of the sound, and it is less for a NO-Hz tone than for a lOOO-Hz tone. On the basis of measurements made in cats (Msller, 1963) the displacement of the stapes (and the cochlear fluid) will lag the sound pressure near the tympanic membrane by about 30 to 40 degrees at 500 Hz, while at 1000 Hz the displacement of the stapes may be shifted by as much as 90 to 120 degrees, relative to the sound pressure waveform at the tympanic membrane, because that frequency is slightly higher than the principal resonance frequency of the human middle ear (about 800 Hz) (Msller, 1963). The distortion that is seen in the recorded potentials to 500-Hz tones of high intensity is dominated by second harmonics, thus resembling a (partial) full-wave rectification. The extra peaks are indications that the condensation phase of the stimulus may become excitatory at high sound levels. This may indicate that neural excitation in the cochlea for low-frequency tones occurs for deflections of the basilar membrane in both directions. However, there may be other reasons for this type of distortion. Thus, several investigators have described ‘peak splitting’ that can occur in the responses from single nerve fibers to lowfrequency tones (Sellick et al., 1982a; Ruggero and Rich, 1983; Ruggero et al., 1986; Kiang, 1984). It seems less likely that the observed waveform distortion is caused by nonlinearities in sound transmission to the cochlea. Studies of sound transmission in the middle ear in the cat show linearity at sound levels well above those used in the present study. The sound produced by the earphones has no noticeable distortion at the sound levels that were used in the present study. It is interesting that the responses recorded in humans seem to have less distortion than those seen in the cat (Snyder and Schremer, 1984). This difference is particularly noticeable at low sound levels, at which the waveform of the responses in the cat has a noticeable distortion, while the responses in man have little distortion below 95 dB SPL at 500 Hz and at 1000 Hz. 173 1 1 I I I 0 5 10 15 20 I 25 ms In interpreting the results of recording of auditory evoked potentials intraoperatively it must be taken into account that such recordings are hampered by technical problems that do not exist when similar recordings are made in experimental animals in the laboratory. For instance, it is not practical to use shielded earphones intraoperatively, nor can bipolar recordings be made, and therefore monopolar recordings have to be used. In addition, the electrical and acoustic noise levels in the operating room are much higher than they are in the laboratory, and there are limitations on the time that can be spent for such recordings. We, however, do not find any evidence that these factors have introduced noticeable errors in the recordings reported on in this paper, but the acoustic noise level in the operating room as well as electrical noise and the limitations of time for recording have not allowed us to record over a large range of sound intensities, as can be done in the laboratory. Contrary to what has been a problem in similar 1200 I I I I I I 0 5 10 15 20 25 1 Ill.3 0 HZ HP I I I I 5 10 15 20 I 25 Ins Fig. 9. The effect on the FFR of masking with highpass-filtered noise. (A) Response to lOOft-Hz tonebursts, 95 dB SPL, without and with masking at cut-off frequencies of 1800 and 3300 Hz. The level of the masking noise was 60 dB SPL/Hz (spectrum level). The results were obtained in a patient operated upon for HFS. (B) Recordings similar to those obtained in A, but from another patient using SOO-Hzand MOO-Hz tonebursts at 104 dB SPL and 95 dB SPL, respectively. The spectrum level of the masking noise was 60 dB. The responses were filtered digitally. The results were obtained in a patient operated upon for TN. 174 recordings in the cat (Snyder and Schreiner, 1984) the recordings from the intracranial portion of the eighth nerve in man were not noticeably contaminated by the CM potential. The reason for this is most likely due to the longer distance between the recording site and the cochlea. In man this distance is about 1.5 cm (Lang, 1981) while in the cat it is probably less than 0.5 cm. The fact that the eighth nerve in man is not usually immersed in fluid when the recordings are made may reduce the conduction of the CM potential to the recording site. When recordings from the human eighth nerve are used to study the normal physiology of the auditory system, it is naturally important to make sure that surgical manipulations to expose the eighth nerve do not injure the auditory nerve, since if this happens the results would naturally not reflect the normal functioning of the ear and the auditory nerve. To help identify cases in which such injuries may have occurred, we always record the farfield response (BAEP) before the beginning of the operation and compare this recording with recordings obtained simultaneously with the recordings from the exposed eighth nerve. For this study, we excluded the results obtained in patients in whom there had been a noticeable change in the latencies of the peaks in the BAEP (about 0.5 ms). This does not, however, assure that the function of the auditory nerve was totally unchanged by the surgical manipulations needed to expose the nerve, but it is the best possible way available to monitor auditory function in anesthetized patients. These problems are not restricted to intraoperative recording in man, but are just as applicable to results obtained in experimental animals in which the eighth nerve must be exposed to obtain the recordings. Acknowledgements This study was supported by a grant from the National Institutes of Health (Grant No. 1 ROlNS21378-04). The authors are grateful to Peter Jannetta, M.D., for making the patients under his care available for this study, and to Margareta B. Moller, M.D., Dr. Med. Sci., for audiological and otological evaluation of the patients in this study. References Goldstein, J.L. and Kiang, N.Y.S. (1958) Synchrony of neural activity in electric response evoked by transient acoustic stimuli. J. Acoust. Sot. Am. 30, 107-114. Jannetta, P.J. (1981a) Hemifacial spasm. In: M. Samii and P.J. Jannetta @is.), The Cranial Nerves. Springer-Verlag, New York, pp. l-8. Jannetta, P.J. (1981b) Vascular decompression in trigeminal neuralgia. In: M. Samii and P.J. Jannetta (I%.), The Cranial Nerves. Springer-Verlag, New York, pp. 331-340. Kiang, N.Y.S. (1984) Peripheral neural processing of auditory information. In: S.R. Geiger (Exe. Ed.), J.M. Brookhart, V.B. Montcastle (Eds.), I. Darian-Smith (Vol. Ed.), Handbook of Physiology, The Nervous System III, Chap. 15. American Physiological Society, Bethesda, Maryland, pp. 6399673. Kim, D.O., Siegel, J.H. and Molnar, C.E. (1979) Co&ear nonlinear phenomena in two-tone responses. Stand. Audiol. (Stockh.) Suppl. 9, 63-81. Konishi, T. and Nielsen, D.W. (1973) The temporal relationship between motion of the basilar membrane and initiation of nerve impulses in the auditory nerve fibers. J. Acoust. Sot. Am. 53, 325. Konishi, T. and Nielsen, D.W. (1978) The temporal relationship between basilar membrane motion and nerve impulse initiation in auditory nerve fibers of guinea pigs. Jpn. J. Physiol. (Tokyo) 28, 291-307. Lang, J. (1981) Facial and vestibulocochlear nerve, topographic anatomy and variations. In: M. Samii and P.J. Jannetta (Eds.), The Cranial Nerves. Springer-Verlag, New York, pp. 363-377. Msller, A.R. (1963) Transfer function of the middle ear. J. Acoust. Sot. Am. 35, 1526-1534. Msller, A.R. (1977) Frequency selectivity of single auditory nerve fibers in response to broadband noise stimuli. J. Acoust. Sot. Am. 62. 135-142. Moller, A.R. (1981a) Neural delay in the ascending auditory pathway. Exp. Brain Res. 43, 93-100. Moller, A.R. (1981b) Latency in the ascending auditory pathway determined using continuous sounds: Comparison between transient and envelope latency. Brain Res. 207, 184-188. Moller, A.R. (1983) Frequency selectivity of phase-locking of complex sounds in the auditory nerve of the rat. Hear. Res. 11, 267-284. Msller, A.R. and Jamretta. P.J. (1983) Monitoring auditory functions during cranial nerve microvascular decompression operations by direct recording from the eighth nerve. J. Neurosurg. 59, 493-499. Moushegian. G., Rupert, A.L. and Stillman, R.D. (1978) Evaluation of frequency-following potentials in man: Masking and clinical studies. Electroenceph. Clin. Neurophysiol. 45, 711-718. Rhode, W.S. (1971) Observations of the vibration of the basilar membrane in squirrel monkeys using the Mossbauer technique. J. Acoust. Sot. Am. 49, 1218-1231. 175 Rhode, W.S. (1978) Some observations on cochlear mechanics. J. Acoust. Sot. Am. 64, 158-176. Ruggero, MA. and Rich, N.C. (1983) Chinchilla auditory-nerve responses to low-frequency tones. J. Acoust. Sot. Am. 73, 2096-2108. Ruggero, M.A., Robles, L. and Rich, N.C. (1986) Basilar membrane mechanics at the base of the chinchilla cochlea. II. Responses to low-frequency tones and relationship to microphonics and spike initiation in the VIII nerve. J. Acoust. Sot. Am. 80, 137551383. Ruggero, M.A. and Rich, N.C. (1987) Timing of spike initiation in cochlear afferents: Dependence on site of innervation. J. Neurophysiol. 58, 3799403. Sellick, P.M., Patuzzi, R. and Johnstone, B.M. (1982a) Modulation of responses of spiral ganglion cells in the guinea pig cochlea by low frequency sound. Hear. Res. 7,199-221. Sellick, P.M., Patuzzi, R. and Johnstone, B.M. (1982b) Measurement of basilar membrane motion in the guinea pig using the Mossbauer technique. J. Acoust. Sot. Am. 72, 131-141. Snyder, R.L. and Schreiner, C.E. (1984) The auditory neurophonic: Basic properties. Hear. Res. 15, 261-280. Snyder, R.L. and Schreiner, C.E. (1985) Forward masking of the auditory nerve neurophonic (ANN) and the frequency following response (FFR). Hear. Res. 20, 45-62. Sokolich, W.G., Hamemik, R.P., Zwislocki, J.J. and Schmiedt, R.A. (1974) Inferred polarities of co&dear hair cells. J. Acoust. Sot. Am. 59, 963-974. Teas, D.C., Eldredge, D.H. and Davis, H. (1962) Cochlear responses to acoustic transients: An interpretation of whole-nerve action potentials. J. Acoust. Sot. Am. 34, 1438-1459. Worden, F.G. and Marsh, J.T. (1968) Frequency-following (microphonic-like) neural responses evoked by sound. Electroenceph. Clin. Neurophysiol. 25, 42-52. Zwislocki, J.J. and Sokolich, W.G. (1973) Velocity and displacement responses in auditory-nerve fibers. Science 182, 64-66. Zwislocki, J.J. (1974) A possible neuro-mechanical sound analysis in the cochlea. Acustica 31, 354-359.


















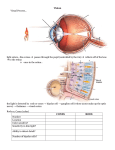





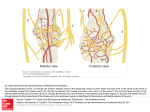
![Unit 8 Review Sheet[1]](http://s1.studyres.com/store/data/001686639_1-accaddf9a4bef8f1f5e508cc8efafb82-150x150.png)