Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
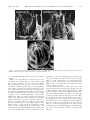
MORPHOLOGY, HISTOLOGY, AND FINE STRUCTURE Functional Morphology of the Male Genitalia of Four Species of Drosophila: Failure to Confirm Both Lock and Key and Male-Female Conflict Predictions WILLIAM G. EBERHARD1 AND NATALIA RAMIREZ Escuela de Biologṍa, Universidad de Costa Rica, Ciudad Universitaria, Costa Rica Ann. Entomol. Soc. Am. 97(5): 1007Ð1017 (2004) ABSTRACT Several hypotheses attempt to explain why male genitalia typically diverge rapidly over evolutionary time. Predictions of the lock and key and the sexually antagonistic coevolution hypotheses were tested by studying the functional morphology of several male genitalic traits by freezing copulating pairs of four species of Drosophila: saltans, willistoni, melanogaster, and malerkotliana. Contrary to the predictions of the hypotheses, there were no species-speciÞc differences in female morphology that corresponded to interspeciÞc differences in the morphology of the male surstylus (which pressed and perhaps spread the distal tip of the oviscape), the epandrium (which grasped the external surface of the extended oviscape), and the aedeagus and paraphyses (which, in willistoni, clamped the oviscape). Antagonistic coevolution could possibly explain the diverse male genitalic morphology if female resistance behavior rather than morphology has coevolved with male morphology, but there are reasons doubt this explanation. KEY WORDS genitalia, cryptic female choice, sexually antagonistic coevolution, lock and key MALE GENITALIA OFTEN DIVERGE relatively rapidly compared with other body parts, and there are several hypotheses that attempt to explain this pattern (Eberhard 1985, Shapiro and Porter 1989, Alexander et al. 1997, Simmons 2001). The lock and key and sexually antagonistic coevolution hypotheses predict that male and female morphology will often coevolve. The lock and key hypothesis (Shapiro and Porter 1989) proposes that female genitalic structures are in effect complex mechanical locks, into which only males of same species can Þt. The sexually antagonistic coevolution hypothesis (Alexander et al. 1997, Holland and Rice 1998, Chapman et al. 2003) proposes that males and females are engaged in coevolutionary arms races over control of copulation and insemination. Female genitalic structures, in particular those that interact mechanically with species-speciÞc male structures, are supposed to constitute antimale devices that impede the manipulative action of male genitalia. Such female defenses result in selection on males favoring further modiÞcations to overcome them. In contrast, a third hypothesis, that male genitalia are under sexual selection by cryptic female choice (Eberhard 1985), does not predict that female morphology will necessarily change when that of the male changes. It is compatible with both female morphological coevolution with male structures, and lack of such evolution. 1 Smithsonian Tropical Research Institute (e-mail: archisepsis@ biologia.ucr.ac.cr). Male and female morphological coevolution has been documented recently in Gerris water striders (Arnqvist and Rowe 1995, 2002a,b), but the lack of species-speciÞc female adjustments to males in several earlier studies of lock and key (Shapiro and Porter 1989) renders the generality of such coevolution uncertain, making further studies desirable. The large genus Drosophila offers advantages for study of interspeciÞc differences in genitalia. The probable relationships between different species groups are well studied (e.g., Pitnick et al. 1999), and male genitalia are generally complex and species-speciÞc. One previous study found a relationship between the form of the ventral process of the male epandrium and the degree of overlap between the femaleÕs eighth and ninth tergites, where the ventral process rests during copulation, in Drosophila melanogaster Meigen and three closely related species; morphological differences were greater between species that showed the greatest mating asymmetries (Robertson 1988). Another study documented male-female coevolution, but between different, internal structures: sperm size and internal female morphology (seminal receptacle) (Pitnick et al. 1999). The current study addresses the external mechanical genitalic mesh between the genitalia of males and females in four species, D. melanogaster, Drosophila saltans Sturtevant, Drosophila malerkrotliana Parshad & Paika, and Drosophila willistoni Sturtevant. All are in the subgenus Sophophora; D. melanogaster and D. malerkotliana are in different subgroups of the melanogaster species group, whereas 0013-8746/04/1007Ð1017$04.00/0 䉷 2004 Entomological Society of America 1008 ANNALS OF THE ENTOMOLOGICAL SOCIETY OF AMERICA Vol. 97, no. 5 Fig. 1. Posterior views of the resting genitalia of noncopulating males of D. saltans, willistoni, melanogaster, and malerkotliana. the other two belong to the saltans and willistoni species groups. Materials and Methods Copulating pairs of virgin ßies were frozen in liquid nitrogen and immediately transferred to an open container of liquid N before they thawed. This container was placed in a ⫺20⬚C freezer, where the N was allowed to boil off, and where the frozen pairs were then ßooded with absolute ethanol at ⫺20⬚C. The pairs were thus not allowed to thaw before being Þxed. After a week in ethanol at ⫺20⬚C, when their frozen tissues had presumably become Þxed, the pairs were brought to room temperature and placed in 80% eth- anol. This treatment guaranteed that the ßiesÕ positions at the moment they were frozen were faithfully preserved (Huber 1993, Eberhard and Pereira 1996). Samples of ⬎10 noncopulating ßies of each sex were prepared the same way, or by direct Þxation in 80% ethanol. The genitalia of males frozen in copula were exposed by carefully dissecting away the female in three to seven pairs of each species. Similar numbers of pairs (four to six) were left intact, except in D. melanogaster, in which 14 intact pairs were examined. Specimens were processed for study with the scanning electron microscope by sublimation drying and coating with gold. In the descriptions of copulating ßies, orientations refer to the position of the female. Flies were September 2004 EBERHARD AND RAMIREZ: FUNCTIONAL MORPHOLOGY Drosophila GENITALIA 1009 Fig. 2. Posterior close-up views of genitalia of noncopulating males of D. saltans (resting), willistoni (resting), melanogaster (directed posteriorly), and malerkotliana. identiÞed by D. Grimaldi (American Museum of Natural History, New York). Results The positions of the genitalia of noncopulating males varied. In the “resting” position, the aedeagus tip was directed dorsally in D. melanogaster but posteriorly in the other species (Fig. 1). Positions of the epandria varied intraspeciÞcally; they were sometimes relatively spread (e.g., D. saltans in Fig. 1) and sometimes more closed (e.g., D. willistoni in Fig. 1). In some D. melanogaster, the aedeagus tip was lowered and directed posteriorly (Fig. 2). In all but D. malerkotliana the phallotreme was visible as a curved slit just anterior to the tip of the aedeagus (Figs. 1 and 2). Some aspects of copulation did not vary intraspeciÞcally and were similar in all four species. The fe- male oviscape was extended distally, a position also seen occasionally in noncopulating females killed in ethanol (Figs. 3 and 4) (in D. melanogaster extension of the oviscape, which was hidden during copulation, was conÞrmed by dissections). The epandria of the male were ßexed medially (Fig. 5), and their distal surfaces pressed against the membranous sides of the extended oviscape (Figs. 6 and 7) (this detail was conÞrmed in D. melanogaster by dissections). The male surstyli, whose species-speciÞc arrays of stout setae (Fig. 8) were directed medially or posteriorly when at rest (Figs. 1 and 2), were ßexed medially ⬇180⬚C, so that their stout setae were directed more or less dorsally. In intact pairs, the stout setae were nearly (D. saltans in Figs. 6, 7, and 9) or completely hidden from view (D. willistoni, D. malerkotliana in Figs. 6 and 9). In D. willistoni and D. saltans (and possibly the other species), these setae pressed Fig. 3. Ventral views of exerted oviscapes of D. saltans, willistoni, melanogaster, and malerkotliana. Fig. 4. Lateral views of exerted oviscapes of D. saltans, willistoni, melanogaster, and malerkotliana. September 2004 EBERHARD AND RAMIREZ: FUNCTIONAL MORPHOLOGY Drosophila GENITALIA 1011 Fig. 5. Ventral views showing spread oviscape valves of females that were frozen during or immediately after copulation ended in saltans, willistoni, and malerkotliana (compare with Fig. 2). on or near the distal tips of the oviscape valves (Figs. 6 and 9). There were also differences between species. In D. saltans, an aedeagal process emerged from the tip of the ejaculatory duct in two males killed during copulation (Fig. 10), but it was not visible in Þve other copulating males and or in noncopulating males (Figs. 1 and 2). No similar aedeagal processes were seen in the other species. The curved plates of the aedeagus of D. malerkotliana (Fig. 9) were unique in being realigned during copulation to form a pair of tubes, one of which apparently contained the phallotreme (Fig. 10). In D. melanogaster, the tips of the male and female abdomens pressed together tightly and hid the oviscape and the positions of the male genitalia relative to it (Fig. 11) (two intact pairs of D. willistoni were also tight together). The distal portions of the relatively prominent ventral lobes of the epandrium in D. melanogaster and D. malerkotliana meshed with the female in different ways. The medial surface of the ventral lobe of the epandrium of D. malerkotliana pressed against the exposed, membranous side of the female oviscape (Fig. 7). In D. melanogaster, in contrast, the distal portion of the ventral lobe of the epandrium was hidden from view under the rear margin of female tergite 8 (Figs. 7 and 11). In two intact pairs of D. saltans, the edge of the surstylus and its stout setae pressed against a process of the paraphysis, apparently trapping the tip of the femaleÕs genital valve between them (Fig. 6). The processes of the paraphysis seemed to cause the tips of femaleÕs genital valves to diverge from each other. In one pair of D. saltans, however, these processes of the paraphyses were not visible (Fig. 9). In D. willistoni, the dorsally curved, sharply pointed paraphyses pressed on the outer, dorsal wall of the oviscape (Fig. 11), evidently trapping the wall in the pincer formed by these points and the sharp ventral tips of the processes of the aedeagus and the aedeagal sheath (Fig. 11). 1012 ANNALS OF THE ENTOMOLOGICAL SOCIETY OF AMERICA Vol. 97, no. 5 Fig. 6. Ventral views of copulating pairs (male above, female below) of D. saltans, willistoni, melanogaster, and malerkotliana, showing grasp with epandria, male cerci, and the medially ßexed sustyli of saltans and willistoni. Discussion In all four species, the male surstyli ßexed medially ⬇180⬚ during copulation. Their stout setae, whose numbers, form, and distribution differ both among the species of this study (Fig. 8) and in other Drosophila (e.g., Pitnick and Heed 1994), apparently pressed against the femaleÕs oviscape, probably on or near the tips of the oviscape valves. They seem to serve to clamp the oviscape and also may spread the oviscape valves apart (Fig. 5), perhaps to allow intromission. Contrary to the predictions of mechanical lock and key and male-female conßict of interests hypotheses, the tips of the oviscape and of the oviscape valves did not show species-speciÞc modiÞcations that corresponded to differences in the setae of the male surstyli (Fig. 3). The male-female conßict hypothesis might escape these criticisms if female genitalic defenses against males involve behavior of the oviscape, rather than its morphology (Eberhard and Pereira 1996). This possibility cannot be ruled out deÞnitively because we did not observe the behavior of the oviscape. Nevertheless, it seems unlikely, because it is difÞcult to imagine what oviscape behavior could have selected for the observed differences in surstylus design. In D. willistoni, the paraphyses press on the outer surface of the femaleÕs oviscape and act as a clamp in opposition to ventral processes on the aedeagus and the aedeagus sheath, which press on the inner surface of the vagina at nearly the same point (Fig. 11). No grasping device of this sort occurred in the other species. There was no special female morphological Fig. 7. Lateral and ventro-lateral views of copulating pairs (male on right, female on left), showing mesh of male epandrium and surstyli with the female oviscape in D. saltans, willistoni, melanogaster, and malerkotliana. Fig. 8. Surstyli in noncopulating males of D. saltans, willistoni, melanogaster, and malerkotliana. 1014 ANNALS OF THE ENTOMOLOGICAL SOCIETY OF AMERICA Vol. 97, no. 5 Fig. 9. Close-up views of male genitalia and the distal portion of female oviscape during copulation (male above, female below) in D. saltans, willistoni, and malerkotliana. courterpart to this male grasping mechanism on the oviscape of female D. willistoni. There are no sclerites or obvious irregularities on the outer surface of the female in this region that could mesh with the male structures in a lock and key manner or that could impede male grasping as expected by the male-female conßict hypothesis. Female D. willistoni shake energetically from side to side during copulation more than the other species (approximately one-quarter of the 8.1 ⫾ 2.0-min copulation) (mean ⫾ SD, N ⫽ 30) (N. Ramirez, unpublished data); but the males were never dislodged, even though they always released their hold with all their legs, folding them immobile and hanging on only with their genitalia (N. Ramirez, unpublished data). Perhaps the female shaking, which seems not to serve to dislodge the male, serves instead to allow the female to sense or test the maleÕs genitalic grip. The positions of our copulating pairs of D. melanogaster were similar to those observed by Robertson (1988) except that both the male epandrium and the female tergite 8 protruded less. Perhaps this difference was due to our Þxing pairs before they thawed. Robertson argued that the mesh between the ventral lobe of the epandrium and the overlap in female tergites eight and nine represented a lock and key mechanism of species isolation. But neither his data nor ours support this idea convincingly: 1) the female tergites are joined by a ßexible membrane, and thus the amount of overlap is not rigid like a mechanical lock but varies with the degree of extension of the femaleÕs abdomen; and 2) there is no physical barrier in the September 2004 EBERHARD AND RAMIREZ: FUNCTIONAL MORPHOLOGY Drosophila GENITALIA 1015 Fig. 10. Genitalia during copulation (female dissected away) of males of D. saltans, willistoni, melanogaster, and malerkotliana. female that would prevent a cross-speciÞc male from seizing her oviscape with his ventral lobes. Data from other Drosophila suggest instead a role for genitalic stimulation of the female. Female D. simulans respond differently when coupled with males of the probable derived sister species D. mauritiana (an island endemic) in which the ventral processes of the epandrium have a different form (Coyne 1993), suggesting that females are able to sense differences in the form of the epandrium. The 19 quantitative trait loci that contribute to the genitalic difference between these species nearly all act in the same direction (Zeng et al. 2000), indicating that the differences are the result of selection; this suggests, in turn, that females can apparently distinguish very small differences (perhaps on the order of 1/19 of the difference between these species). We are unable to assign possible functions to many portions of the male genitalia that enter the female rather than remaining on her outer surface during copulation, including the aedeagus and its processes (in species other than D. willistoni), the gonopods, and the paraphyses (in species other than D. willistoni). Some of these structures, such as the aedeagus, are often species-speciÞc in form in Drosophila. Study of sections of copulating pairs would probably help clarify their functions. It is important to note several limitations of this study. Male and female genitalia may change positions during copulation, as suggested by the differences in the position of the paraphysis and aedeagal processes in D. saltans (see Lachmann 1996 and Eberhard and Huber 1998 for changes in other acalyptrate ßies). Position changes may be especially likely in the very early stages of coupling. Our small sample sizes and lack of information on the stage of copulation of each pair did not allow us to examine these possibilities. Rhythmic movements, such as apparent male scraping on the sides of the oviscape with his epandrium in D. saltans (N. Ramirez, unpublished data) and possible female movements, also were not covered by this study (see Eberhard 2001a,b for rhythmic movements of male genitalia in other ßies). 1016 ANNALS OF THE ENTOMOLOGICAL SOCIETY OF AMERICA Vol. 97, no. 5 Fig. 11. Top, lateral view of male epandrium (right) and its mesh with the female in a copulating pair (left; male is on the right) of D. melanogaster; the ventral process of epandrium in inserted beneath the female tergite 8. Bottom, lateral view of the close apposition of processes of the maleÕs aedeagus and aedeagal sheath with the tips of his paraphyses during copulation in D. willistoni (female dissected away) (right) and of one male paraphysis pressed against the external dorsal surface the femaleÕs oviscape during copulation (left). Acknowledgments We thank David Grimaldi for kindly identifying ßies and for help with genitalic terminology, Jerry Coyne (University of Chicago) for helpful comments, and Maribelle Vargas (Universidad de Costa Rica) for outstanding technical assistance. Financial support was provided by the Smithsonian Tropical Research Institute. References Cited Alexander, R. D., D. C. Marshall, and J. R. Cooley. 1997. Evolutionary perspectives on insect mating, pp. 4 Ð31. In J. Choe and B. Crespi [eds.], The evolution of mating systems in insects and arachnids. Cambridge University Press, Cambridge, United Kingdom. Arnqvist, G., and L. Rowe. 1995. Sexual conßict and arms races between the sexes: a morphological adaptation for control of mating in a female insect. Proc. R. Soc. Lond B. 261: 123Ð127. Arnqvist, G., and L. Rowe. 2002a. Antagonistic evolution between the sexes in a group of insects. Nature (Lond.) 415: 787Ð789. Arnqvist, G., and L. Rowe. 2002b. Correlated evolution of male and female morphologies in water striders. Evolution 56: 936 Ð947. Chapman, T., G. Arnqvist, J. Bangham, and L. Rowe. 2003. Sexual conßict. Trends Ecol. Evol. 18: 41Ð 47. Coyne, J. A. 1993. The genetics of an isolating mechanism between two sibling species of Drosophila. Evolution 47: 778 Ð788. Eberhard, W. G. 1985. Sexual selection and animal genitalia. Harvard University Press, Cambridge, MA. Eberhard, W. G. 2001a. Species-speciÞc genitalic copulatory courtship in sepsid ßies (Diptera, Sepsidae, Microsepsis). Evolution 55: 93Ð102. Eberhard, W. G. 2001b. Genitalic behavior during copulation in Hybosciara gigantea (Diptera: Sciaridae) and the evolution of species-speciÞc genitalia. J. Kans. Entomol. Soc. 74: 1Ð9. Eberhard, W. G., and B. A. Huber. 1998. Copulation and sperm transfer in Archisepsis ßies (Diptera: Sepsidae) and the evolution of their intromittent genitalia. Studia Dipt. 5: 217Ð248. Eberhard, W. G., and F. Pereira. 1996. Functional morphology of male genitalic surstyli in the dungßies Achisepsis September 2004 EBERHARD AND RAMIREZ: FUNCTIONAL MORPHOLOGY Drosophila GENITALIA diversiformis and A. ecalcarata (Diptera: Sepsidae). J. Kans. Entomol. Soc. 69: 43Ð 60. Holland, B., and W. R. Rice. 1998. Chase-away selection: antagonistic seduction vs. resistance. Evolution 52: 1Ð7. Huber, B. A. 1993. Genital mechanics and sexual selection in the spider Nesticus cellulanus (Araneae: Nesticidae). Can. J. Zool. 71: 2437Ð2447. Lachmann, A. D. 1996. Copulation and engagement of male and female genitalia in Þve Coproica Rondani species (Diptera: Sphaeroceridae). Ann. Entomol. Soc. Am. 89: 759 Ð769. Pitnick, S., and W. B. Heed. 1994. New species of cactusbreeding Drosophila (Diptera: Drosophilidae) in the nannoptera species group. Ann. Entomol. Soc. Am. 87: 307Ð 310. Pitnick, S., T. Markow, and G. S. Spicer. 1999. Evolution of multiple kinds of female sperm-storage organs in Drosophila. Evolution 53: 1804 Ð1822. 1017 Robertson, H. M. 1988. Mating asymmetries and phylogeny in the Drosophila melanogaster species complex. Pac. Sci. 42: 72Ð 80. Shapiro, A. M., and A. H. Porter. 1989. The lock-and-key hypothesis: evolutionary and biosystematic interpretation of insect genitalia. Annu. Rev. Entomol. 34:231Ð245. Simmons, L. 2001. Sperm competition and its evolutionary consequences in insects. Princeton University Press, Princeton, NJ. Zeng, Z.-B., J. Lin, L. F. Stam, C.-H. Kao, J. M Mercer, and C. C. Laurie. 2000. Genetic architecture of a morphological shape difference between two Drosophila species. Genetics 154: 299 Ð310. Received 31 July 2003; accepted 3 March 2004.