Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
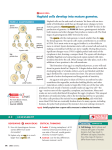
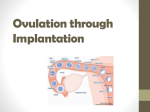
Gametogenesis • Reproduction in vertebrates is by sexual means involving haploid (1N) germ cells – Ovum = female component – Spermatozoa = male component • Both arise through meiosis = cell division where each daughter cell receives ½ genetic material from original cell • Primordial germ cells derived from extraembryonic endoderm (yolk sac) migrate to gonads Gametogenesis • Oogenesis occurs in Ovary within a follicle of epithelial cells • Spermatogenesis occurs in germinal epithelium lining seminiferous tubules of testis • Oogenesis begins with oogonium; Spermatogenesis begins with spermatogonium – Both are normal 2N cells • Reduction in chromosome number accomplished via two meiotic divisions Stages in Gametogenesis 1. Pairing and doubling of chromosomes in ____gonia, followed by growth as primary ____ocyte (2 X 2N) 2. 1st meiotic division produces two 2 X 1N cells (= secondary ____ocytes) 3. 2nd meiotic division produces four haploid cells (spermatids, ova) 4. Spermatids mature and differentiate to form functional spermatozoa • In spermatogenesis, all 4 sperm cells produced are viable • In oogenesis, only 1 of 4 cells produced is viable. – Others become abortive as polar bodies (only small amount of cytoplasm) that later degenerate Fig 14.22 - Oogenesis Fig 14.30 - Spermatogenesis Egg Membranes and Structure • Cytoplasm enclosed within plasma membrane • Vitelline membrane = thin membrane closely attached to plasma membrane – Zona pellucida = glycoprotein layer (mammals) • Corona radiata (mammals) = layer of follicle cells that become sloughed off after fertilization Sperm Structure • Spermatozoa from different animals have a wide variety of forms • All have head and tail regions – Head region serves two functions: • Contains nucleus (genetic function) • Acrosomal cap = contains enzymes that allow sperm to break down membranes around egg and fertilize egg • Tail = flagellum that provides motility • Midpiece between head and tail contains mitochondria that provide ATP to fuel swimming Ovarian follicle Primary follicle Ovum Zona pellucida Spermatozoa Fertilization • Several obstacles must be overcome for successful fertilization: 1. Sperm and egg must come into proximity 2. Cell to cell contact must occur 3. Sperm must penetrate egg cell Mechanisms for Proximity • Transport occurs in liquid medium • EXTERNAL FERTILIZATION – Eggs and sperm simultaneously shed into water – Occurs in fishes (except Chondrichthyes) and most anurans • INTERNAL FERTILIZATION – Sperm introduced directly into female tract – Usually involves copulatory organs in males (none present in tuatara, birds, salamanders = copulation by cloacal “kiss”) – Occurs in animals with shelled eggs or viviparous habits as sperm must reach egg before shell is added (Chondrichthyes, most Amphibians, Amniotes) Mechanisms for Contact • For internal fertilization, sperm travel within female tract by passive transport (dependent on muscular contractions and ciliary currents provided by female tract). – Little active swimming by sperm for transport function • Contact in external fertilization accomplished by random swimming movements of sperm in water Mechanisms for Egg Barrier Penetration • Once contact with egg has been established, the next step is to penetrate the egg so that nuclear materials can unite to form the diploid zygote. • Barrier penetration mechanisms are chemical in nature and involve acrosomal reaction – Sperm Lysins = enzymes that locally dissolve egg membranes – Produced by acrosomal cap – Sperm lysins differ among animal groups as membranes surrounding eggs differ (e.g., jelly coat in amphibians, follicle cells of corona radiata in mammals) Mechanisms for Egg Barrier Penetration • Acrosome Reaction involves … 1. Release of sperm lysins 2. Fusion of egg and sperm membranes — In some animals, acrosomal reaction involves exposure of binding sites on plasma membrane of sperm, via acrosomal tubule or filament, which bind to receptors on p.m. of egg in species-specific manner — This binding precedes fusion of sperm and egg plasma membranes rupture sperm lysins Acrosomal Reaction in Hemichordates Binding sites exposed that bind to receptors on egg plasma membrane Mechanisms for Egg Barrier Penetration • In mammals, there is no development of acrosomal filaments • Instead, fluids of female reproductive tract induce capacitation primes sperm for fertilization and includes removal of some components from sperm surface. • After capacitation, hyaluronidase on the sperm head is exposed and breaks down the hyaluronic acid cementing the follicle cells of corona radiata (which surround the egg) together allows sperm passage through corona radiata to contact zona pellucida (a glycoprotein layer surrounding the egg) Mechanisms for Egg Barrier Penetration • Zona pellucida has species-specific receptors for binding sperm – Binding causes rupture of acrosome, which releases contents that break down zona pellucida and allow contact with egg plasma membrane – Binding also exposes proteins on sperm surface that bind with receptors on egg plasma membrane to facilitate fusion of sperm and egg • Fusion of plasma membranes releases sperm genetic material into egg as sperm pronucleus • Male and female genetic material will soon combine forming a diploid zygote Post-fertilization Responses in Zygote 1. Formation of Fertilization Cone = outward bulge of egg cytoplasm that serves to engulf sperm – Occurs upon fusion of sperm and egg plasma membranes – Recession of cone brings sperm nucleus into egg cytoplasm 2. Egg Activation — Upon fusion (within 3 sec) get membrane depolarization or hyperpolarization (speciesdependent) blocks entrance of > 1 sperm (= Fast block to polyspermy) Post-fertilization Responses in Zygote — Next, get Ca2+ release from internal stores within egg triggers cortical reaction release of cortical granules to perivitelline space around egg — Cortical granule release causes development of fertilization membrane blocking further sperm entry (= Slow block to polyspermy) — Slow block to polyspermy occurs about 25-30 sec post-fusion — Seems to occur only for microlecithal eggs (e.g., mammals); entrance of > 1 sperm into eggs of birds, reptiles and some amphibians common, but only 1 sperm contributes to zygote (others somehow inactivated) Fast Block to Polyspermy Slow Block to Polyspermy Post-fertilization Responses in Zygote 3. Rearrangement of internal constituents within egg — Sets up gradients of certain substances and plane of bilateral symmetry within zygote for some animals 4. Fusion of Haploid Nuclei — In most vertebrates, meiosis within egg arrested after 1st meiotic division. Sperm entry stimulates 2nd meiotic division to produce female pronucleus (and 2nd polar body) — Once this 2nd division occurs, female pronucleus is ready for union with male pronucleus Post-fertilization Responses in Zygote 4. Fusion of Haploid Nuclei (cont.) — Male and female pronuclei next approach each other (mechanism by which this movement occurs is not known with certainty) — Next get fusion of pronuclei — In some animals (including most vertebrates), pronucleus membrane degenerates free chromosomes arrange themselves at spindle (metaphase of mitosis) completion of mitosis dipolid zygote Parthenogenesis • Definition = development of the egg in the absence of sperm • Occurrence suggests that: – (1) egg activation and nuclear fusion are separate developmental processes – (2) the ovum contains all the capacities necessary for embryo formation – all that is necessary is some triggering agent • Eggs can be activated by a number of chemical, thermal, electrical or mechanical means Parthenogenesis • Parthenogenetic individuals are expected to be haploid (and many are), but these embryos are often diploid. • Doubling of chromosomes accomplished in 3 ways: 1. Suppression of 2nd meiotic division – occurs only in eggs completing this division after fertilization 2. Refusion with second polar body 3. Suppression of 1st mitotic division (= 1st cleavage division) Parthenogenesis • Haploid embryos generally show premature developmental arrest • Parthenogenetic diploid embryos also usually show premature developmental arrest • However, in several invertebrates parthenogenesis is normal (e.g., male drones of bee colony) and there are several species of naturally occurring parthenogenetic lizards (the entire population is female) • Artificial selection procedures have developed parthenogenetic strain of turkeys The asexual, all-female whiptail species Cnemidophorus neomexicanus (center), which reproduces via parthenogenesis, is shown flanked by two sexual species having males, C. inornaus (left) and C. tigris (right), which hybridized naturally to form the C. neomexicanus species. Methods of Bearing Young • Oviparous = egg laying – Primitive condition for vertebrates – Occurs in most fishes, amphibians, reptiles, all birds, monotremes • Viviparous = live-bearing – Advanced condition in vertebrates – Some live-bearers occur in all vertebrate classes except cyclostomes and birds – Evolved by retention of eggs within body to increase survival of young “Placental Connections” in Viviparous Vertebrates • Anamniotes with connection between yolk sac and maternal tissues through which exchange of metabolites occurs (e.g., Chondrichthyes) • Reptiles use yolk sac, chorion, allantois (extraembryonic membranes) or some combination for connection • Mammals with a variety of connections Early Development/Placentation in Mammals • After formation of zygote cleavage produces blastula • Blastula forms before embryo reaches uterus • Mammalian blastula consists of trophoblast and inner cell mass (ICM becomes embryo) • Upon reaching uterus, trophoblast overlying ICM makes contact with uterine endometrium trophoblast cells rapidly multiply and insert among epithelial cells lining endometrium and endometrial cells degenerate implantation • Continued trophoblast cell division placentation; embryo becomes buried w/in endometrial lining Fig 5.32 Mammalian Placenta Formation • Structure produced by apposition and fusion of extraembryonic membranes of embryo with uterine endometrium of mother • Extraembryonic membranes = tissues external to embryo not participating in embryo formation, but functioning in maintenance of the embryo • In Amniotes, four extraembryonic membranes exist Extraembryonic Membranes • Yolk Sac = forms from extraembryonic hypomere (splanchnopleure) that expands to enclose yolk – This is the only extraembryonic membrane present in Anamniotes, so it occurs in all vertebrates – Functions to derive nutrients from yolk in yolky eggs to nourish developing embryo • In Amniotes, extraembryonic somatopleure grows over embryo by folding back on itself producing a double hood of somatopleure • From this structure develop Amnion and Chorion Extraembryonic Membranes • Amnion = forms from inner somatopleure + ectoderm (inside) • Chorion = forms from outer somatopleure + ectoderm (outside) • Amnion serves as fluid-filled sac for embryonic development – Replicates aquatic developmental environment of primitive vertebrates. – Allows complete conquest of terrestrial habitats • Chorion functions in protection of embryo and in exchange of gases (and metabolites in placenta) Extraembryonic Membranes • Outgrowth of splanchnopleure from posterior region of gut in Amniotes eventually expands to fill extraembryonic coelom (= space between amnion and chorion) • This membrane is the Allantois = composed of splanchnic mesoderm (outside) + endoderm • Mesoderm fuses with mesoderm of chorion to form Chorioallantoic Membrane = main gas exchange organ for Amniote embryos • Allantois also serves waste storage function Fig 5.29 – Extraembryonic membrane formation in a bird Fig 5.30 Fig 5.30 – Extraembryonic membrane formation in a bird Mammalian Placenta • From chorion (outermost extraembryonic membrane), finger-like processes grow outward to interlock with uterine endometrium • Blood streams of mother and fetus never mix – always separated by epithelial membrane, so exchange of gases and nutrients occurs by diffusion across this membrane • Chorion is not in direct contact with embryo so some means of blood supply from embryo to placenta (and back) must occur Mammalian Placenta • Blood Supply to developing embryo differs between marsupial and placental mammals • Marsupials = mostly Choriovitelline fetal placenta – Yolk sac associated with inner surface of chorion – Blood vessels develop in mesoderm of yolk sac – This situation also occurs to some extent in several placental groups (e.g., rodents) • Placentals = Chorioallantoic fetal placenta – Dominant connection to chorion provided by allantois, yolk sac usually degenerates – Allantoic mesoderm forms blood vessels that function in gas & nutrient exchange, waste removal Fig 5.33 – Fetal extraembryonic membranes in various Amniotes Mammalian Placenta Types • Primitive Condition = apposition without fusion (non-deciduous) • Advanced Condition = fusion of maternal and fetal tissues (deciduous) • Four Types occur: 1. Epitheliochorial = most primitive — Occurs in pig and some other mammals — Maternal and fetal blood separated by 6 layers: endothelium, CT, epithelium, epithelium, CT, endothelium Mammalian Placenta Types 2. Syndesmochorial = no uterine epithelium — Occurs in ruminant mammals (cattle, sheep, etc.) 3. Endotheliochorial = no maternal epithelium or CT — Occurs in carnivores 4. Hemochorial = advanced condition — Chorionic epithelium bathed in maternal blood — Occurs in primates and many rodents