Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
Plant breeding wikipedia , lookup
Island restoration wikipedia , lookup
Biological Dynamics of Forest Fragments Project wikipedia , lookup
Human impact on the nitrogen cycle wikipedia , lookup
Coevolution wikipedia , lookup
Punctuated equilibrium wikipedia , lookup
Invasive species wikipedia , lookup
Molecular ecology wikipedia , lookup
Introduced species wikipedia , lookup
Latitudinal gradients in species diversity wikipedia , lookup
Theoretical ecology wikipedia , lookup
Ecological fitting wikipedia , lookup
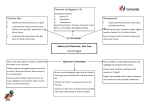
Forum Commentary When two invasion hypotheses are better than one Dozens of hypotheses attempt to explain the success of invasive species (Catford et al., 2009). Though no single hypothesis explains invasions in general (Gurevitch et al., 2011), the temptation to search for a universal mechanism shared by most invasive species has motivated much of the invasion research to date. Instead of studying mechanisms in isolation from one another, perhaps investigating synergies between disparate invasion hypotheses, as Zheng et al. have in this issue of New Phytologist (pp. 1350–1359), will do more to explain the success of invasive species. ‘The strength of this approach is that it used crosscontinental studies to provide evidence for multiple ecological and evolutionary invasion mechanisms in a Multiple biotic factors and the ERH Over seven previously described invasion mechanisms focus on biotic factors (Catford et al., 2009), including enemies, competitors, and mutualists. Some of these hypotheses explicitly consider interactions between multiple factors (e.g. competition and herbivory in the Enemy of My Enemy Hypothesis (Colautti et al., 2004)), yet relatively few empirical studies consider effects of multiple types of biotic interactions simultaneously. We conducted an extensive search of the ERH literature and found only 19 studies that manipulated more than one biotic interaction on native and introduced species simultaneously (E. H. Schultheis et al., unpublished). All but one study was conducted on plants, and most were conducted in the introduced range, not comparing the effects of biotic interactions in both ranges. We found evidence that studying more than one biotic interaction revealed important effects that studying one biotic interaction alone would have missed. For example, herbivores had a stronger effect on invasive plant performance when competition was intense (Suwa & Louda, 2012), although other studies found that the effects of multiple biotic factors were additive, and studying them in isolation provided accurate estimates of their effects (e.g. Cripps et al., 2011). real invasion.’ Resources and the ERH From a basic ecology perspective, it is not terribly surprising that one mechanism does not underlie all invasions, given the wide variety of factors limiting the population growth and distributions of native taxa. At least two potential scenarios likely explain the observed lack of generality in invasions (Gurevitch et al., 2011): first, invaders likely succeed through many different mechanisms. In essence, most of the proposed invasion hypotheses may be relevant for some taxa, in some environments, at some point in time but do not apply in the majority of cases. Second, proposed invasion mechanisms may act synergistically to promote invasions. Though this second point is not a new concept, in the invasion literature (Inderjit et al., 2005; Gurevitch et al., 2011), it has rarely been investigated empirically. Multicausality and interactions between invasion mechanisms Multiple factors may contribute to invasions, and these may act additively or interactively. Effects may be minimal when each mechanism is studied in isolation, and their full effects may only be understood when studied in concert with other mechanisms. Next, we highlight three examples of potential synergies between the Enemy Release Hypothesis (ERH), one of the most extensively tested invasion hypotheses, and other invasion mechanisms. 958 New Phytologist (2015) 205: 958–960 www.newphytologist.com Invasion may be facilitated by disturbance, which frees up resources making them available for introduced species, or anthropogenic addition of nutrients to the environment (Davis et al., 2000). These nutrient effects may interact with enemy release. Faster growing species tend to invest more in growth and less in producing high quality, well-defended tissues. Because these fast growing species may be susceptible to enemies when they become abundant and apparent, these same species may be more likely to benefit from enemy release. As a result, invasiveness may be promoted most when invaders receive the dual benefits of high resource availability and enemy release (Resource Enemy Release Hypothesis, Blumenthal, 2006). Darwin’s naturalization hypothesis and the ERH Darwin’s naturalization hypothesis predicts that species closely related to native community members will be poor invaders because close relatives are more likely to be functionally similar and intense competitors (Darwin, 1859). While the naturalization hypothesis was initially based on assumptions about competitive interactions, novel defenses may also facilitate invasion success through interactions with the ERH. Introduced species with very different defenses may succeed because native herbivores or predators may not have the capacity to recognize or tolerate unique defense compounds or behaviors to which they have limited Ó 2015 The Authors New Phytologist Ó 2015 New Phytologist Trust New Phytologist Commentary evolutionary experience. For example, introduced plants more distantly related to the community they invade often experience reduced herbivore damage (e.g. Harvey et al., 2012). A necessary future step for these types of studies, however, is determining whether reduced herbivory received by more distantly related invaders actually promotes invasiveness. Synergy between ERH, EICA and the NWH Synergies between some invasion mechanisms may be more likely than between others. Given that many novel weapons and defenses in plants involve the production of potentially costly chemicals, Forum 959 synergies between the Novel Weapons Hypothesis (NWH) and the ERH and/or the Evolution of Increased Competitive Ability (EICA) hypothesis may be especially likely (Fig. 1; see also Callaway & Ridenour, 2004; Bossdorf, 2013). The NWH suggests that the success of some invaders is due to the novelty of their competitive, defensive, or predatory traits (Callaway & Ridenour, 2004). In essence, native species may have no evolutionary experience with, and therefore may be extremely susceptible to, allelopathic or defensive compounds produced by invaders. The EICA hypothesis, in turn, proposes that escape from herbivores causes evolutionary reductions in defense and resource reallocation to growth and competitive ability (Blossey & N€otzold, 1995). While EICA Native range community Generalist herbivores Specialist herbivores Competing plants 1 Invaded range community Generalist herbivores Specialist herbivores Naïve competing plants 2 6 Reduced attack from specialist enemies 4 Reduced defense 8 3 Increased allelochemical effectiveness 5 7 Increased competitive ability, growth, fitness Fig. 1 Plant species encounter very different communities in their invaded range compared to their native range; they typically escape specialist enemies and encounter different competing plant species (1). Because of the absence of many specialist enemies in the introduced range, they may experience reduced attack from specialist herbivores (2). Because herbivory can reduce plant growth and competitive abilities, plants may experience increased fitness in the introduced range (3). Reduced attack from specialist enemies may also relax selection on plant defenses (4). Because of growth–defense trade-offs, relaxed selection on plant defenses may lead to the evolution of increased growth or competitive ability (5). Invading plants also face different competitive environments (6). Competing plants in the introduced range may be na€ıve to and more affected by novel competitive traits of the invader such as allelochemicals, leading to higher growth and relatively higher competitive abilities in the introduced range (7). These relationships represent three common invasion mechanisms: Enemy Release Hypothesis (ERH: yellow symbols, 1–2–3), the Evolution of Increased Competitive Ability (EICA) hypothesis (gray symbols, 1–2–4–5) and the Novel Weapons Hypothesis (NWH: green symbols, 1–6–7). The ERH/EICA and NWH may be linked if biochemical or resource-based trade-offs between defensive and offensive chemical production exist (blue symbol, 8). In this case, escape from enemies may relax selection on defenses allowing for increased allocation to offensive chemical compounds over evolutionary time. Solid and dashed arrows indicate ecological and evolutionary processes respectively. Ó 2015 The Authors New Phytologist Ó 2015 New Phytologist Trust New Phytologist (2015) 205: 958–960 www.newphytologist.com 960 Forum New Phytologist Commentary explicitly assumes a growth–defense trade-off, EICA and NWH may be tightly related if evolutionary increases in competitive ability do not arise from a growth–defense trade-off but instead from an offense–defense trade-off. For example, reduced allocation to defensive chemicals may allow increased production of allelochemicals. If these trade-offs between defensive chemical production and allelochemical production are common, then novel weapons may be especially potent mechanisms of invasion when accompanied by enemy release. Integrating these hypotheses provides a mechanism explaining increased allelochemical production and competitive ability following invasion. Several recent studies have used very different approaches for investigating connections between NWH and EICA/ERH. Zheng et al. link findings from separate experiments to build their case that novel weapons and EICA are related. Their approach focuses on identifying differences between invasive and native range genotypes in a variety of biotic contexts (presence/absence of competitors and enemies in both the native and introduced range). The strength of this approach is that it used crosscontinental studies to provide evidence for multiple ecological and evolutionary invasion mechanisms in a real invasion. As a result, synergistic effects of NWH and EICA seem plausible, but the connections between the two invasion mechanisms remain tenuous; for example, NWH and EICA are tested in separate experiments and not linked together by testing for genetic correlations between allelochemical and defense production. Using a different approach, Uesugi & Kessler (2013) employ experimental evolution to show the potential for enemy release to select for increased competitive ability and allelochemical production, but it is unclear whether increased allelochemical production evolved in invaded range populations of their study taxa or what trade-offs produced this evolutionary shift. Both approaches illustrate that NWH and EICA are two invasion mechanisms with obvious potential for synergistic effects, but the details of this synergism remain speculative. For example, we now understand the role of Odoratin in allelopathic interactions, but does it also play a role in deterring belowground herbivores or altering belowground microbial communities, a hypothesis consistent with Zheng et al. findings of increased defenses against belowground enemies and increased Odoratin production in invaded range genotypes? Similarly, do trade-offs between the production of the allelochemical Odoratin and other defense traits explain the observed reductions in resistance and increases in allelopathy in introduced populations? Only more detailed functional ecology studies of the chemical compounds involved, and explicit quantitative genetic studies to identify the potential for correlated evolution and genetic trade-offs, will fully link these invasion mechanisms. As these studies illustrate, simultaneously considering multiple invasion mechanisms is difficult, requiring a number of separate experiments, long-term experimental evolution approaches, or ideally fully factorial experiments with the capacity for considering both ecological and evolutionary responses. Yet, these types of experiments may help unify the many disparate hypotheses of invasions and identify when invasion mechanisms are most likely to New Phytologist (2015) 205: 958–960 www.newphytologist.com act synergistically. In short, while no magic bullet fully explains invasiveness, combining multiple invasion hypotheses may lead to important insights on the drivers of biological invasions and the constraints and opportunities faced by species embedded in complex natural communities more generally. Acknowledgements This is Kellogg Biological Station publication no. 1843. Jennifer A. Lau* and Elizabeth H. Schultheis Kellogg Biological Station & Department of Plant Biology, Michigan State University, Hickory Corners, MI 49060, USA (*Author for correspondence: tel +1 269 671 2107; email [email protected]) References Blossey B, N€otzold R. 1995. Evolution of increased competitive ability in invasive nonindigenous plants: a hypothesis. Journal of Ecology 83: 887–889. Blumenthal DM. 2006. Interactions between resource availability and enemy release in plant invasion. Ecology Letters 9: 887–895. Bossdorf O. 2013. Enemy release and evolution of increased competitive ability: at last, a smoking gun!. New Phytologist 198: 638–640. Callaway RM, Ridenour WM. 2004. Novel weapons: invasive success and the evolution of increased competitive ability. Frontiers in Ecology and the Environment 2: 436–443. Catford JA, Jansson R, Nilsson C. 2009. Reducing redundancy in invasion ecology by integrating hypotheses into a single theoretical framework. Diversity and Distributions 15: 22–40. Colautti RI, Ricciardi A, Grigorovich IA, MacIsaac HJ. 2004. Is invasion success explained by the enemy release hypothesis? Ecology Letters 7: 721–733. Cripps MG, Bourdot GW, Saville DJ, Hinz HL, Fowler SV, Edwards GR. 2011. Influence of insects and fungal pathogens on individual and population parameters of Cirsium arvense in its native and introduced ranges. Biological Invasions 13: 2739–2754. Darwin C. 1859. On the origin of species by means of natural selection. London, UK: John Murray. Davis MA, Grime JP, Thompson K. 2000. Fluctuating resources in plant communities: a general theory of invasibility. Journal of Ecology 88: 528–534. Gurevitch J, Fox GA, Wardle GM, Inderjit S, Taub D. 2011. Emergent insights from the synthesis of conceptual frameworks for biological invasions. Ecology Letters 14: 407–418. Harvey KJ, Nipperess DA, Britton DR, Hughes L. 2012. Australian family ties: does a lack of relatives help invasive plants escape natural enemies? Biological Invasions 14: 2423–2434. Inderjit S, Cadotte MW, Colautti RI. 2005. The ecology of biological invasions: past, present and future. In: Inderjit S, ed. Invasive plants: ecological and agricultural aspects. Basel, Switzerland: Birkh€auser Verlag, 19–44. Suwa T, Louda SM. 2012. Combined effects of plant competition and insect herbivory hinder invasiveness of an introduced thistle. Oecologia 169: 467–476. Uesugi A, Kessler A. 2013. Herbivore-exclusion drives the evolution of plant competitiveness via increased allelopathy. New Phytologist 198: 916–924. Zheng Y-L, Feng Y-L, Zhang L-K, Callaway RM, Valiente-Banuet A, Luo D-Q, Liao Z-Y, Barclay GF, Silva-Pereyra C. 2015. Integrating novel chemical weapons and evolutionarily increased competitive ability in success of a tropical invader. New Phytologist 205: 1350–1359. Key words: allelopathy, biological invasion, defense, enemy release, invasive species, novel weapons, trade-off. Ó 2015 The Authors New Phytologist Ó 2015 New Phytologist Trust