Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
History of herbalism wikipedia , lookup
Ecology of Banksia wikipedia , lookup
Photosynthesis wikipedia , lookup
Gartons Agricultural Plant Breeders wikipedia , lookup
Plant nutrition wikipedia , lookup
Plant stress measurement wikipedia , lookup
History of botany wikipedia , lookup
Plant secondary metabolism wikipedia , lookup
Plant defense against herbivory wikipedia , lookup
Plant use of endophytic fungi in defense wikipedia , lookup
Ornamental bulbous plant wikipedia , lookup
Pollination wikipedia , lookup
Evolutionary history of plants wikipedia , lookup
Plant breeding wikipedia , lookup
Plant physiology wikipedia , lookup
Plant morphology wikipedia , lookup
Plant evolutionary developmental biology wikipedia , lookup
Plant ecology wikipedia , lookup
Sustainable landscaping wikipedia , lookup
Perovskia atriplicifolia wikipedia , lookup
Flowering plant wikipedia , lookup
PLANT KINGDOM
Kingdom Plantae popularly known as the ‘plant kingdom’. We must know that
the plant kingdom has changed over time. Fungi, and members of the Monera
and Protista having cell walls have now been excluded from Plantae though
earlier classifications put them in the same kingdom. So, the cyanobacteria that
are also referred to as blue green algae are not ‘algae’ any more.
Let us look to different form of classifications:
Artificial System of Classification of Plants:
i. The system of classification in which one or very few character are considered
as the key feature of classification is called as artificial system.
ii. This system of classification never throws light on the affinities or relationship
of the plants with one another.
iii. This classification is made only on the basis of presence on absence of the
particular character that taken into account leaving the others.
iv. In artificial system plants are categorized into a particular taxonomical rank
but the ranks are not stepwise.
v. In this classification very few species are included which were discovered until
that period.
There are many taxonomist classify their own way of understanding in to
different group taking one or few characters. These plants are grouped in to
artificial groups or classes or orders. Out of these artificial system, the system
which primarily proposed by Linnaeus was taken to consideration.
Natural Classification:
George Bentham (1800-1884) and Sir Joseph Dalton Hooker (1817-1911) were
great plant explorer and geographer associated with royal botanical gardens and
adopted a very comprehensive system of classification in their jointly published
book genera pantarum.
Feature of Classification:
i. This includes the names and descriptions of all genera of seed plant known so
far and classified accordingly.
ii. The plant kingdom comprises about 97205 species of seed plantsunder 202
orders in which orders treated now as families.
iii. This orders further grouped under several cohorts, now treated as orders.
iv. This orders further grouped under several cohorts, now treated as orders.
v. They placed first dicotyledons, then gymnosperms and lastly the
monocotyledons.
vi. They classify dicotyledonsinto 165 order gymnosperms into 3 orders and
mono cotyledons into 34 orders.
vii. They devide dicotyledons in to 3 divisions and 14 series which further
divided into cohorts and orders.
viii. The dicots started with family ranunculadceae, with free sepals and petals
and indefinite number of stamens and carpels are free. The dicots e4nds with
family labiatae with fused swpals and petals with definite number of carpels and
stamens.
ix. Monocots divided into seven series. Which are directed grouped into orders
without interpretation of cohorts.
Orders with epigenous flowers, i.e. orchidaceous and scitaminae were kept first,
follower by orders with petaloid hypogynous flowers, i.e. liliaceae. Then kept
palmaceae and araceae, finally ended with graminal and cyperaceae.
Phylogenetic Classification:
Phylogeny
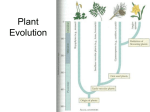
The evolutionary history of a group of genetically related organisms is called a
phylogeny. It includes ancestor species and descendant species. A phylogeny is
usually represented by a tree diagram called a phylogenetic tree. An early
example of a phylogenetic tree is Darwin’s “Tree of Life” (see Figure below). In
this diagram, Darwin was trying to show how he thought evolution had occurred.
The tree shows how species evolved through time, from the bottom of the tree to
the top. As species evolved, they formed new branches on the tree of life. Some of
these species eventually branched into additional descendant species. Others
died out, or went extinct, without leaving any descendants.
. This branching diagram represents the evolutionary histories of different
species. It is the only diagram that originally appeared in Darwin
Modern biologists still use phylogenetic trees to represent evolutionary
histories. A simple phylogenetic tree is shown in Figure below. The tips of the
branches represent genetically related species. The branching points represent
common ancestors. A common ancestor is the last ancestor species that two
descendant species shared before they took different evolutionary paths. In the
tree in Figure below, species 1 and 2 shared a more recent common ancestor
with each other than with species 3. Therefore, species 1 and 2 are more closely
related to one another than to species 3.
. This phylogenetic tree shows how hypothetical species 1, 2, and 3 are related to
one another through common ancestors.
Ancestor species are like your own ancestors. Your most recent common
ancestor with any siblings you may have is a shared parent. Your most recent
common ancestor with a first cousin is a shared grandparent. Your most recent
common ancestor with a second cousin is a shared great-grandparent. In general,
the more distant the relationship between you and relatives in your own
generation, the farther in the past you shared a common ancestor. The same
holds true for related species. The more distant the relationship between two
related species, the farther back in time they shared a common ancestor.
Numerical Taxonomy which is now easily carried out using computers is based
on all observable characteristics. Number and codes are assigned to all the
characters and the data are then processed. In this way each character is given
equal importance and at the same time hundreds of characters can be
considered. Cytotaxonomy that is based on cytological information like
chromosome number, structure, behaviour and chemotaxonomy that uses the
chemical constituents of the plant to resolve confusions, are also used by
taxonomists these days.
.Gross Morphology: done on the basis of functions.
.Vegetative Characterstics: Plant morphology treats both the vegetative
structures of plants, as well as the reproductive structures. The vegetative
(somatic) structures of vascular plants include two major organ systems: (1) a
shoot system, composed of stems and leaves, and (2) a root system.
.Androecium:
Flowers are made up of both reproductive and non-reproductive structures
arranged in four whorls. These include the calyx, the corolla, the gynoecium, and
the androecium.
1 Calyx: outermost whorl made up of usually green, leaf-like structures
called sepals
2 Corolla: whorl that contains often brightly colored petals
3 Androecium: third whorl that contains male reproductive structures
called stamens
4 Gynoecium: innermost whorl made up of female reproductive structures
called carpels
Both the calyx and corolla are the non-reproductive structures of a flower, while
the androecium and gynoecium are the reproductive structures. The gynoecium
produces egg cells, and the androecium produces sperm cells. In this lesson, we
will focus on the structure of the androecium.
.Cytological: study of cell
ALGAE
Algae:
.chlorophyll
.simple
.thalloid (An organism or structure resembling a thallus)
Thallus
.autotrophic
.largely aquatic (fresh water and marine)
They occur in variety of other habitats like moist stones, soils and wood. Some of
them also occur in association with fungi (lichen) and animals (sloth bear).
Sloth bear
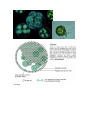
The size ranges from the microscopic unicellular forms like Chlamydomonas, to
colonial forms like Volvox and to the filamentous forms like Ulothrix and
Spirogyra. A few of the marine forms such as kelps, form massive plant bodies.
VOLVOX
Ulothrix
REPRODUCTION: ALGAE
The algae reproduce by vegetative, asexual and sexual methods. Vegetative
reproduction is by fragmentation. Each fragment develops into a thallus. Asexual
reproduction is by the production of different types of spores, the most common
being the zoospores. They are flagellated (motile) and on germination gives rise
to new plants. Sexual reproduction takes place through fusion of two gametes.
These gametes can be flagellated and similar in size (as in Chlamydomonas) or
non-flagellated (non-motile) but similar in size (as in Spirogyra). Such
reproduction is called isogamous (form of sexual reproduction that involves
gametes of similar morphology). Fusion of two gametes dissimilar in size, as in
some species of Chlamydomonas is termed as anisogamous. Fusion between one
large, non-motile (static) female gamete and a smaller, motile male gamete is
termed oogamous, e.g., Volvox, Fucus.
FUCUS
USES: ALGAE
Algae are useful to man in a variety of ways. At least a half of the total carbon
dioxide fixation on earth is carried out by algae through photosynthesis. Being
photosynthetic they increase the level of dissolved oxygen in their immediate
environment. They are of importance as primary producers of energy-rich
compounds which form the basis of the food cycles of all aquatic animals. Many
species of Porphyra, Laminaria and Sargassum are among the 70 species of
marine algae used as food. Certain marine brown and red algae produce large
amounts of hydrocolloids (water holding substances), e.g., algin (brown algae)
and carrageen (red algae) are used commercially. Agar, one of the commercial
products obtained from Gelidium and Gracilaria are used to grow microbes and
in preparations of ice-creams and jellies. Chlorella and Spirullina are unicellular
algae, rich in proteins and are used as food supplements even by space travellers.
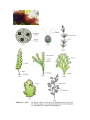
The algae are divided into three main classes: Chlorophyceae, Phaeophyceae
and Rhodophyceae
TYPES: ALGAE
Chlorophyceae
The members of chlorophyceae are commonly called green algae. The plant
body may be unicellular, colonial or filamentous. They are usually grass green
due to the dominance of pigments chlorophyll a and b. The pigments are
localised in definite chloroplasts. The chloroplasts may be discoid (disc like
structure), plate-like, reticulate ,cup-shaped, spiral or ribbon-shaped in different
species. Most of the members have one or more storage bodies called pyrenoids
located in the chloroplasts. Pyrenoids contain protein besides starch. Some algae
may store food in the form of oil droplets. Green algae usually have a rigid cell
wall made of an inner layer of cellulose and an outer layer of pectose (something
insoluble to water).
(reticulate)
Vegetative reproduction usually takes place by fragmentation or by formation of
different types of spores. Asexual reproduction is by flagellated zoospores
produced in zoosporangia. The sexual reproduction shows considerable
variation in the type and formation of sex cells and it may be isogamous,
anisogamous or oogamous. Some commonly found green algae are:
Chlamydomonas, Volvox, Ulothrix, Spirogyra and Chara
CHLOROPHYLL
Chlorophyll a
Chlorophyll a is a specific form of chlorophyll used in oxygenic photosynthesis.
It absorbs most energy from wavelengths of violet-blue and orange-red light.[3] It
also reflects green/yellow light, and as such contributes to the observed green
color of most plants. This photosynthetic pigment is essential for photosynthesis
in eukaryotes, cyanobacteria and prochlorophytes because of its role as primary
electron donor in the electron transport chain.
Chlorophyll b
Chlorophyll b is a form of chlorophyll. Chlorophyll b helps in photosynthesis by
absorbing light energy. It is more soluble than chlorophyll a in polar solvents
because of its carbonyl group. Its color is yellow, and it primarily absorbs blue
light.
Chlorophyll c
Chlorophyll c is a form of chlorophyll found in certain marine algae, including
the photosynthetic Chromista(separate kingdom to protista) (e.g. diatoms,
brown algae) and dinoflagellates.
Chlorophyll d
Chlorophyll d is a form of chlorophyll,
identified by Harold Strain and Winston
Manning in 1943.[1][2] It is present in marine red
algae and cyanobacteria which use energy
captured from sunlight for photosynthesis.[3]
Chlorophyll d absorbs far-red light, at 710 nm
wavelength, just outside the optical range.[4] An
organism that contains chlorophyll d is adapted
to an environment such as moderately deep
water, where it can use far red light for
photosynthesis,[5] although there is not a lot of
visible light
Chlorophyll c
Phaeophyceae
The members of phaeophyceae or brown algae are found primarily in marine
habitats. They show great variation in size and form. They range from simple
branched, filamentous forms (Ectocarpus) to branched forms as represented by
kelps (large seaweeds), which may reach a height of 100 metres. They possess
chlorophyll a, c, carotenoids (any of a class of mainly yellow, orange, or red fatsoluble pigments, including carotene, which give colour to plant parts such as
ripe tomatoes and autumn leaves) and xanthophylls (yellow pigments that occur
widely in nature and form one of two major divisions of the carotenoid group).
They vary in colour from olive green to various shades of brown depending upon
the amount of the xanthophyll pigment, fucoxanthin (accessory pigment in the
chloroplasts of brown algae)present in them.
Food is stored as complex carbohydrates, which may be in the form of laminarin
(The molecule laminarin (also known as laminaran) is a storage glucan (a
polysaccharide (polymeric carbohydrate molecules composed of long chains of
monosaccharide units) of glucose) found in brown algae)or mannitol. The
vegetative cells have a cellulosic wall usually covered on the outside by a
gelatinous coating of algin. The protoplast contains, in addition to plastids, a
centrally located vacuole and nucleus. The plant body is usually attached to the
substratum by a holdfast, and has a stalk, the stipe and leaf like photosynthetic
organ – the frond.
Vegetative reproduction takes place by fragmentation. Asexual reproduction in
most brown algae is by biflagellate (two flagella) zoospores that are pear-shaped
and have two unequal laterally attached flagella.
Sexual reproduction may be isogamous, anisogamous or oogamous. Union of
gametes may take place in water or within the oogonium (oogamous species).
The gametes are pyriform (pear-shaped) and bear two laterally attached
flagella. The common forms are Ectocarpus, Dictyota, Laminaria, Sargassum and
Fucus .
Polymer: large molecule, or macromolecule, composed of many repeated
subunits. Because of their broad range of properties, both synthetic and natural
polymers play an essential and ubiquitous role in everyday life
Ectocarpus
Mannitol: a colourless sweet-tasting crystalline alcohol which is found in many
plants and is used in various foods and medical products.
Rhodophyceae
Rhodophyta are commonly called red algae because of the red pigment, rphycoerythrin (red protein-pigment) in their body. Majority of the red algae are
marine with greater concentrations found in the warmer areas. They occur in
both well-lighted regions close to the surface of water and also at great depths in
oceans where relatively little light penetrates.
Red algae are multicellular. Some of them have complex body organisation. The
food is stored as floridean (type of storage glucan) starch which is very similar to
amylopectin (highly branched polymer found in plants)and glycogen (form of
storage in animals and fungi) in structure.
The red algae usually reproduce vegetatively by fragmentation. They
reproduce asexually by non-motile spores and sexually by non-motile
gametes. Sexual reproduction is oogamous and accompanied by complex
post fertilisation developments (After fertilization, a series of event occurs in
the zygote to develop into a seed). The common members are: Polysiphonia,
Porphyra , Gracilaria and Gelidium.
Gracilaria
Gelidium
BRYOPHYTES
Bryophytes include the various mosses and liverworts that are found commonly
growing in moist shaded areas in the hills . Bryophytes are also called
amphibians of the plant kingdom because these plants can live in soil but are
dependent on water for sexual reproduction. They usually occur in damp, humid
and shaded localities. They play an important role in plant succession on bare
rocks/soil.
PLANT SUCCESSION
Many plant communities are not self-sustaining. A field in the temperate
deciduous forest biome will remain a field only as long as it is grazed by animals
or mowed regularly. If these factors are eliminated, the balance tips in favor of
other species. The newcomers will, in turn, establish conditions that no longer
favor them but promote the growth of still other species. The revered naturalist
and writer, Henry David Thoreau, called this process succession.
Primary Plant Succession
The process of plant succession begins just as soon as a land area capable of
supporting plant life is formed. Some examples:
5 accumulation of sand dunes at the edge of the ocean or a lake
6 cooling of a lava flow
7 exposure of rock by a retreating glacier
Bare rock succession in a temperate deciduous forest biome
• The first colonizers are lichens and certain mosses. Acids secreted by the
lichens attack the rock and provide bits of soil. Additional soil particles
may be formed by weathering or be blown in from elsewhere. Damage
and decay to the lichens supplies some humus, and eventually enough soil
is formed to support other mosses.
• The growth, death, and decay of mosses produces more humus, and soon
there is enough to support the growth of
• grasses and
• shrubby growth such as low-bush blueberries and huckleberries. These,
in turn, provide the conditions for such sun-loving, fast-growing species
as
• gray birch trees and poplars (quaking aspens).
• In time, white pines replace these. In the dense shade of mature white
pines, only shade-tolerant maples and beech seedlings can gain a
foothold. If the shallow-rooted white pines are removed by a hurricane or
by lumbering, the maples and beeches can take over.
Bog succession
Another example of plant succession occurs as shallow ponds gradually fill in
with soil washed in from the surrounding terrain and organic matter produced
by underwater plants.
As we walk from the edge of a poorly-drained, boggy pond back into a temperate
deciduous forest, we pass through a series of zones that recreate in space the
plant succession that has been occurring in time.
• From the swamp loosestrife at the waters edge past
• sphagnum moss, leatherleaf, sheep laurel, and pitcher plants, then
• swamp azaleas, high-bush blueberries and poison sumac, followed by
• black spruce and American larch and, finally,
• swamp maples and alders
one passes concentric zones, each representing a later stage of plant succession
as the soil has become firmer and the shade denser.
Secondary Plant Succession
Lumbering, farming, fires, and hurricanes interrupt the process of succession by
removing the dominant plants in the community. Their elimination sets the stage
for a new succession to begin.
The many abandoned farms in New England (I live on one) illustrate this. People
often wonder why our pioneers built stone walls through the woods. The answer
is that they did not. The walls in the woods today once marked the boundaries of
fields and pastures, but when cultivation and grazing ceased, a secondary
succession began. Where I live,
•
•
•
•
•
the grass of abandoned fields soon became invaded by low-growing,
herbaceous species like plantains, and golden rods.
These were quickly followed by woody shrubs like the common juniper,
blueberries and gray-stemmed dogwood
Soon sun-loving gray birch, poplars, and Eastern red cedar ("old-field
cedar") became established.
White pines or, in sandy well-drained locations, oaks have followed.
These will persist until some disturbance such as fire, hurricanes, or
lumbering open up the land, and the process of secondary succession
begins again.
In general, plant succession is a reflection of the increasing efficiency of the
community at intercepting the energy of the sun and converting it into chemical
energy. As one stage of succession follows another,
• the biomass of the community increases. This is the outcome of the
increasing amount of
• net productivity — calories stored by the plant community.
• This, in turn, provides calories for a larger community of consumers.
• As succession continues, the diversity of species in the community
increases — at least for a time.
• When the system approaches its climax, the rate of increase in net
productivity of the plants is consumed by its own heterotrophs.
• The system comes into equilibrium and reaches peak efficiency at
channeling the energy of the sun into the food web of the community.
The graph (from Whittaker, R. H., Communities and Ecosystems, Macmillan, 1970)
shows the changes in number of species, biomass, and net productivity during
secondary succession in a temperate deciduous forest over a period of 160 years.
The plant body of bryophytes is more differentiated than that of algae. It is
thallus-like and prostrate (growing along the ground)or erect (straight), and
attached to the substratum by unicellular or multicellular rhizoids. They lack
true roots, stem or leaves. They may possess root-like, leaf-like or stem-like
structures.
Rhizoid: a filamentous outgrowth or root hair on the underside of the thallus in
some lower plants, especially mosses and liverworts, serving both to anchor the
plant and (in terrestrial forms) to conduct water.
The main plant body of the bryophyte is haploid (having a single set of unpaired
chromosomes). It produces gametes, hence is called a gametophyte. The sex
organs in bryophytes are multicellular. The male sex organ is called
antheridium. They produce biflagellate antherozoids. The female sex organ
called archegonium is flask-shaped and produces a single egg. The antherozoids
are released into water where they come in contact with archegonium. An
antherozoid fuses with the egg to produce the zygote (an ovum (female gamete)
and a sperm cell (male gamete)—combine to form a single diploid cell). Zygotes do not
undergo reduction division immediately. They produce a multicellular body
called a sporophyte. The sporophyte is not free-living but attached to the
photosynthetic gametophyte and derives nourishment from it. Some cells of the
sporophyte undergo reduction division (meiosis) to produce haploid spores.
These spores germinate to produce gametophyte.
Bryophytes in general are of little economic importance but some mosses
provide food for herbaceous mammals, birds and other animals. Species of
Sphagnum, a moss, provide peat that have long been used as fuel, and because of
their capacity to hold water as packing material for trans-shipment of living
material. Mosses along with lichens are the first organisms to colonise rocks and
hence, are of great ecological importance. They decompose rocks making it
suitable for the growth of higher plants. Since mosses form dense mats on the
soil, they reduce the impact of falling rain and prevent soil erosion. The
bryophytes are divided into liverworts and mosses.
LIVERWORTS
The liverworts grow usually in moist, shady habitats such as banks of streams,
marshy ground, damp soil, bark of trees and deep in the woods. The plant body
of a liverwort is thalloid, e.g., Marchantia. The thallus is dorsiventral (having
dissimilar dorsal and ventral surfaces)and closely appressed (push) to the
substrate (layer). The leafy members have tiny leaf-like appendages (a thing that
is added or attached to something larger or more important) in two rows on the
stem-like structures.
Asexual reproduction in liverworts takes place by fragmentation of thalli, or by
the formation of specialised structures called gemmae (sing. gemma). Gemmae
are green, multicellular, asexual buds, which develop in small receptacles (a
hollow object used to contain something) called gemma cups located on the
thalli. The gemmae become detached from the parent body and germinate to
form new individuals. During sexual reproduction, male and female sex organs
are produced either on the same or on different thalli.
The sporophyte is differentiated into a foot, seta and capsule. After meiosis,
spores are produced within the capsule. These spores germinate to form freeliving gametophytes.
A sporophyte is the diploid multicellular stage in the life cycle of a plant or alga.
It develops from the zygote produced when a haploid egg cell is fertilized by a
haploid sperm and each sporophyte cell therefore has a double set of
chromosomes, one set from each parent.
Mosses
The predominant stage of the life cycle of a moss is the gametophyte which
consists of two stages. The first stage is the protonema (A protonema (plural:
protonemata) is a thread-like chain of cells that forms the earliest stage (the
haploid phase) of a bryophyte or pteridophyte life cycle)stage, which develops
directly from a spore. It is a creeping, green, branched and frequently
filamentous stage. The second stage is the leafy stage, which develops from the
secondary protonema as a lateral bud. They consist of upright, slender axes
bearing spirally arranged leaves. They are attached to the soil through
multicellular and branched rhizoids. This stage bears the sex organs.
Vegetative reproduction in mosses is by fragmentation and budding in the
secondary protonema. In sexual reproduction, the sex organs antheridia (male
sex organ) and archegonia (female sex organ) are produced at the apex
(highest)of the leafy shoots. After fertilisation, the zygote develops into a
sporophyte, consisting of a foot, seta (stalk supporting the capsule of a moss or
liverwort, and supplying it with nutrients) and capsule.
The sporophyte in mosses is more elaborate than that in liverworts. The capsule
contains spores. Spores are formed after meiosis. The mosses have an elaborate
mechanism of spore dispersal. Common examples of mosses are Funaria,
Polytrichum and Sphagnum.
PTERIDOPHYTES
The Pteridophytes include horsetails (Equisetum or horse tails is the only living
genus in Equisetaceae, a family of vascular plants that reproduce by spores
rather than seeds) and ferns. Pteridophytes are used for medicinal purposes and
as soil-binders (Soil binders consist of applying and maintaining a soil stabilizer
to exposed soil). They are also frequently grown as ornamentals. Evolutionarily,
they are the first terrestrial plants to possess vascular tissues – xylem and
phloem.
In pteridophytes, the main plant body is a sporophyte which is differentiated
into true root (A true root system consists of a primary root and secondary roots
or lateral roots), stem and leaves . These organs possess well-differentiated
vascular tissues. The leaves in pteridophyta are small (microphylls: very short
leaf) as in Selaginella (Selaginella is the sole genus of vascular plants in the
family Selaginellaceae, the spikemosses or lesser clubmosses) or large
(macrophylls) as in ferns. The sporophytes bear sporangia that are subtended by
leaf-like appendages called sporophylls. In some cases sporophylls may form
distinct compact structures called strobili or cones (Selaginella, Equisetum or
horse tail). The sporangia produce spores by meiosis in spore mother cells. The
spores germinate to give rise to inconspicuous (not clearly visible), small but
multicellular, free-living, mostly photosynthetic thalloid gametophytes called
prothallus. These gametophytes require cool, damp, shady places to grow.
Because of this specific restricted requirement and the need for water for
fertilisation, the spread of living pteridophytes is limited and restricted to
narrow geographical regions. The gametophytes bear male and female sex
organs called antheridia and archegonia, respectively. Water is required for
transfer of antherozoids – the male gametes released from the antheridia, to the
mouth of archegonium. Fusion of male gamete with the egg present in the
archegonium result in the formation of zygote. Zygote thereafter produces a
multicellular well-differentiated sporophyte which is the dominant phase of the
pteridophytes. In majority of the pteridophytes all the spores are of similar
kinds; such plants are called homosporous. Genera like Selaginella and Salvinia
(Salvinia, a genus in the family Salviniaceae, is a floating fern named in honor of
Anton Maria Salvini, a 17th-century Italian scientist. Watermoss is a common
name for Salvinia) which produce two kinds of spores, macro (large) and micro
(small) spores, are known as heterosporous. The megaspores and microspores
germinate and give rise to female and male gametophytes, respectively. The
female gametophytes in these plants are retained on the parent sporophytes for
variable periods. The development of the zygotes into young embryos take place
within the female gametophytes. This event is a precursor to the seed habit
considered an important step in evolution.
SEED HABIT
A seed consists of an embryo, stored food and a seed coat. The seed habit is
the most complex and evolutionary successful method of sexual reproduction. It
is found in vascular pIants. Today, seed plants, gymnosperms and angiosperms
flowering plants are the most diverse lineage within the vascular plants. Most of
this diversity in angiosperms occurred during Cretaceous time. The seed plants
have an adaptive advantage. They occur in a wide variety of habitats and
dominate today’s flora. This evolutionary success is due to the seed. It is one of
the most dramatic innovations during land plant evolution. The origin and
evolution of the seed habit was started in late Devonian times about 385 M.
The pteridophytes are further classified into four classes: Psilopsida (Psilotum);
Lycopsida (Selaginella, Lycopodium), Sphenopsida (Equisetum) and Pteropsida
(Dryopteris, Pteris, Adiantum).
Horse-tails
Clubmosses: Lycopodium is a genus of clubmosses, also known as ground pines
or creeping cedar, in the family Lycopodiaceae, a family of fern-allies
Selaginella
Lycopodium
Sphenopsida
Pteropsida
Dryopteris, Pteris, Adiantum
GYMNOSPERMS
The gymnosperms (gymnos : naked, sperma : seeds) are plants in which the
ovules are not enclosed by any ovary wall and remain exposed, both before and
after fertilisation. The seeds that develop post-fertilisation, are not covered, i.e.,
are naked. Gymnosperms include medium-sized trees or tall trees and shrubs
(Figure 3.4). One of the gymnosperms, the giant redwood tree Sequoia is one of
the tallest tree species. The roots are generally tap roots. Roots in some genera
have fungal association in the form of mycorrhiza (Pinus), while in some others
(Cycas) small specialised roots called coralloid roots are associated with N2fixing cyanobacteria. The stems are unbranched (Cycas) or branched (Pinus,
Cedrus). The leaves may be simple or compound. In Cycas the pinnate leaves
persist for a few years. The leaves in gymnosperms are well-adapted to
withstand extremes of temperature, humidity and wind. In conifers, the needlelike leaves reduce the surface area. Their thick cuticle (a protective and waxy or
hard layer covering the epidermis of a plant, invertebrate, or shell) and sunken
(having sunk or been submerged in water) stomata also help to reduce water
loss.
The gymnosperms are heterosporous (Heterospory is the production of spores
of two different sizes and sexes by the sporophytes of land plants); they produce
haploid microspores and megaspores. The two kinds of spores are produced
within sporangia that are borne on sporophylls which are arranged spirally
along an axis to form lax or compact strobili or cones. The strobili bearing
microsporophylls and microsporangia (a sporangium containing
microspores)are called microsporangiate or male strobili. The microspores
develop into a male gametophytic generation which is highly reduced and is
confined to only a limited number of cells. This reduced gametophyte is called a
pollen grain. The development of pollen grains take place within the
microsporangia. The cones bearing megasporophylls with ovules or
megasporangia are called macrosporangiate or female strobili. The male or
female cones or strobili may be borne on the same tree (Pinus) or on different
trees (Cycas). The megaspore mother cell is differentiated from one of the cells of
the nucellus. The nucellus is protected by envelopes and the composite structure
is called an ovule. The ovules are borne on megasporophylls which may be
clustered to form the female cones. The megaspore mother cell divides
meiotically to form four megaspores. One of the megaspores enclosed within the
megasporangium (nucellus) develops into a multicellular female gametophyte
that bears two or more archegonia or female sex organs. The multicellular
female gametophyte is also retained within megasporangium.
Unlike bryophytes and pteridophytes, in gymnosperms the male and the female
gametophytes do not have an independent free-living existence. They remain
within the sporangia(a receptacle in which asexual spores are formed)retained on
the sporophytes. The pollen grain is released from the microsporangium. They
are carried in air currents and come in contact with the opening of the ovules
borne on megasporophylls. The pollen tube carrying the male gametes grows
towards archegonia in the ovules and discharge their contents near the mouth of
the archegonia. Following fertilisation, zygote develops into an embryo and the
ovules into seeds. These seeds are not covered.
.EMBRYO: an unborn or unhatched offspring in the process of development
Sequoia
Cedrus , Conifers
ANGIOSPERMS
Unlike the gymnosperms where the ovules are naked, in the angiosperms or
flowering plants, the pollen grains and ovules are developed in specialised
structures called flowers. In angiosperms, the seeds are enclosed by fruits. The
angiosperms are an exceptionally large group of plants occurring in wide range
of habitats. They range in size from tiny, almost microscopic Wolfia to tall trees
of Eucalyptus (over 100 metres). They provide us with food, fodder, fuel,
medicines and several other commercially important products. They are divided
into two classes : the dicotyledons and the monocotyledons. The dicotyledons
are characterised by having two cotyledons in their seeds while the
monocolyledons have only one. The male sex organs in a flower is the stamen.
Each stamen consists of a slender filament with an anther at the tip. The anthers,
following meiosis, produce pollen grains. The female sex organs in a flower is the
pistil or the carpel. Pistil consists of an ovary enclosing one to many ovules.
Within ovules are present highly reduced female gametophytes termed embryosacs. The embryo-sac formation is preceded by meiosis. Hence, each of the cells
of an embryo-sac is haploid. Each embryo-sac has a three-celled egg apparatus –
one egg cell and two synergids, three antipodal cells and two polar nuclei. The
polar nuclei eventually fuse to produce a diploid secondary nucleus. Pollen grain,
after dispersal from the anthers, are carried by wind or various other agencies to
the stigma of a pistil. This is termed as pollination. The pollen grains germinate
on the stigma and the resulting pollen tubes grow through the tissues of stigma
and style and reach the ovule. The pollen tubes enter the embryo-sac where two
male gametes are discharged. One of the male gametes fuses with the egg cell to
form a zygote (syngamy). The other male gamete fuses with the diploid
secondary nucleus to produce the triploid primary endosperm nucleus (PEN).
Because of the involvement of two fusions, this event is termed as double
fertilisation, an event unique to angiosperms. The zygote develops into an
embryo (with one or two cotyledons) and the PEN develops into endosperm
which provides nourishment to the developing embryo. The synergids and
antipodals degenerate after fertilisation. During these events the ovules develop
into seeds and the ovaries develop into fruit.
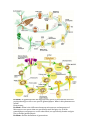
PLANT LIFE CYCLES AND ALTERNATION OF GENERATIONS
NCERT
In plants, both haploid and diploid cells can divide by mitosis. This ability leads
to the formation of different plant bodies - haploid and diploid. The haploid plant
body produces gametes by mitosis. This plant body represents a gametophyte.
Following fertilisation the zygote also divides by mitosis to produce a diploid
sporophytic plant body. Haploid spores are produced by this plant body by
meiosis. These in turn, divide by mitosis to form a haploid plant body once again.
Thus, during the life cycle of any sexually reproducing plant, there is an
alternation of generations between gamete producing haploid gametophyte and
spore producing diploid sporophyte.
However, different plant groups, as well as individuals representing them, differ
in the following patterns:
1. Sporophytic generation is represented only by the one-celled zygote.
There are no free-living sporophytes. Meiosis in the zygote results in the
formation of haploid spores. The haploid spores divide mitotically and
form the gametophyte. The dominant, photosynthetic phase in such
plants is the free-living gametophyte. This kind of life cycle is termed as
haplontic. Many algae such as Volvox, Spirogyra and some species of
Chlamydomomas represent this pattern.
2. On the other extreme, is the type wherein the diploid sporophyte is the
dominant, photosynthetic, independent phase of the plant. The
gametophytic phase is represented by the single to few-celled haploid
gametophyte. This kind of lifecycle is termed as diplontic. All seedbearing plants i.e. gymnosperms and angiosperms, follow this pattern .
3. Bryophytes and pteridophytes, interestingly, exhibit an intermediate
condition (Haplo-diplontic); both phases are multicellular and often
free-living. However, they differ in their dominant phases.
A dominant, independent, photosynthetic, thalloid or erect phase is
represented by a haploid gametophyte and it alternates with the short- lived
multicelluler sporophyte totally or partially dependent on the gametophyte
for its anchorage and nutrition. All bryophytes represent this pattern.
The diploid sporophyte is represented by a dominant, independent,
photosynthetic, vascular plant body. It alternates with multicellular,
saprophytic/autotrophic, independent but short-lived haploid gametophyte.
Such a pattern is known as haplo-diplontic life cycle. All pteridophytes
exhibit this pattern (Figure 3.7 c).
Interestingly, while most algal genera are haplontic, some of them such as
Ectocarpus, Polysiphonia, kelps are haplo-diplontic. Fucus, an alga is
diplontic.
REFERENCE
THE LIFE CYCLE OF PLANTS
Alternation of Generations
All plants undergo a life cycle that takes them through both haploid and diploid
generations. The multicellular diploid plant structure is called the sporophyte,
which produces spores through meiotic (asexual) division. The multicellular
haploid plant structure is called the gametophyte, which is formed from the
spore and give rise to the haploid gametes. The fluctuation between these diploid
and haploid stages that occurs in plants is called the alternation of generations.
The way in which the alternation of generations occurs in plants depends on the
type of plant. In bryophytes (mosses and liverworts), the dominant generation is
haploid, so that the gametophyte comprises what we think of as the main plant.
The opposite is true for tracheophytes (vascular plants), in which the diploid
generation is dominant and the sporophyte comprises the main plant.
Bryophyte Generations
Bryophytes are nonvascularized plants that are still dependent on a moist
environment for survival (see Plant Classification, Bryophytes . Like all plants,
the bryophyte life cycle goes through both haploid (gametophyte) and diploid
(sporophyte) stages. The gametophyte comprises the main plant (the green moss
or liverwort), while the diploid sporophyte is much smaller and is attached to
the gametophyte. The haploid stage, in which a multicellular haploid
gametophyte develops from a spore and produces haploid gametes, is the
dominant stage in the bryophyte life cycle. The mature gametophyte produces
both male and female gametes, which join to form a diploid zygote. The zygote
develops into the diploid sporophyte, which extends from the gametophyte and
produces haploid spores through meiosis. Once the spores germinate, they
produce new gametophyte plants and the cycle continues.
Tracheophyte Generations
Tracheophytes are plants that contain vascular tissue; two of the major classes of
tracheophytes are gymnosperms (conifers) and angiosperms (flowering plants).
Tracheophytes, unlike bryophytes, have developed seeds that encase and protect
their embryos. The dominant phase in the tracheophyte life cycle is the diploid
(sporophyte) stage. The gametophytes are very small and cannot exist
independent of the parent plant. The reproductive structures of the sporophyte
(cones in gymnosperms and flowers in angiosperms), produce two different
kinds of haploid spores: microspores (male) and megaspores (female). This
phenomenon of sexually differentiated spores is called heterospory. These
spores give rise to similarly sexually differentiated gametophytes, which in turn
produce gametes. Fertilization occurs when a male and female gamete join to
form a zygote. The resulting embryo, encased in a seed coating, will eventually
become a new sporophyte.
Problem : In gymnosperms and angiosperms, spores of two distinct sexes are
produced and give rise to sex-specific gametophytes. What is this phenomenon
called?
Heterospory.
Problem : What is the difference between microspores and megaspores?
Microspores are spores that are specifically male and give rise to male
gametophytes; megaspores, on the other hand, are specifically female and give
rise to female gametophytes.
Problem : Define alternation of generations.
The fluctuation between the diploid (sporophyte) and haploid (gametophyte) life
stages that occurs in plants.
Problem : Through what process do sporophytes generate spores?
Meiosis.
Fertilization in plants occurs when haploid gametes meet to create a diploid
zygote, which develops into an embryo. In gymnosperms (conifers) and
angiosperms (flowering plants), the meeting of the gametes occurs in the
following way: male gametes are enclosed in pollen grains and are carried by
wind or insects to the female reproductive organs. The final product of
fertilization--the embryo--is encased in a seed. For this reason, these two types
of tracheophytes are termed seed plants.
Gymnosperm Fertilization
The female gametophyte contains several archegonia, where the egg cells
originate and develop. The gametophyte itself is surrounded by layers of
sporangia and integument; all of these elements comprise an ovule, which is
found on the surface of a female cone. Fertilization occurs when pollen grains
(male gametophytes) are carried by the wind to the open end of an ovule, which
contains the eggs, or female gametophyte. There, the pollen grain develops an
outgrowth called a pollen tube, which eventually penetrates to the egg cell
within one of the archegonia. The sperm cells within the pollen tube then vie to
fertilize the egg. Once fertilization has occurred, the embryo develops within the
female gametophyte, and the ovule becomes the seed, complete with a food
source (the gametophyte tissue) and a seed coat (the integument). This embryo,
which will eventually become a new sporophyte, consists of two embryonic
leaves, the epicotyl and hypocotyl.
Angiosperm Fertilization
The female reproductive organ of angiosperms is the pistil, located in the middle
of the flower. As in gymnosperms, the male gametophyte is the pollen grain. In
order for fertilization to occur in most flowering plants, insects or other animals
must transport the pollen to the pistil. A major distinguishing feature of
angiosperms is the practice of double fertilization.
Figure %: Double Fertilization
An angiosperm ovule contains an egg cell and a diploid fusion nucleus, which is
created through the joining of two polar nuclei within the ovule. When a pollen
grain comes into contact with the stigma, or top of the pistil, it sends a pollen
tube down into the ovary at the pistil's base. As the pollen tube penetrates the
ovule, it releases two sperm cells. One fuses with the egg to create a diploid
zygote, while the other joins with the fusion nucleus to form a triploid nucleus.
This triploid nucleus turns into an endosperm, which nourishes the developing
embryo (filling the role of gametophyte tissue in the gymnosperm seed). As in
gymnosperms, the ovule becomes a seed, encasing the embryo and endosperm in
a seed coat. But unlike gymnosperms, in angiosperms the ovary containing the
ovules develops into a fruit after fertilization. The fruit gives the embryos the
double benefit of added protection against desiccation and increased dispersal,
since it is eaten by far-ranging animals who then excrete the seeds.
In order for fertilization to occur, angiosperms either self-pollinate, in which a
particular plant fertilizes itself, or cross-pollinate, in which one plant is fertilized
by another of the same species. Cross-pollination generally produces far more
vigorous plants, and is encouraged through differential development of the male
and female gametophytes on a flower, or through the positioning of these
gametophytes so that self- pollination is difficult.
Problem : How does a pollen grain function in fertilization?
Pollen grains, or the male gametophytes of flowering plants, carry the sperm
cells to the female reproductive organs. They are transported either by the wind
(as in conifers) or by insects (as in most flowering plants).
Problem : What are the three main components of a seed, and from what are
they derived?
The three components of the seed are the embryo, the food source (derived from
gametophyte tissue in gymnosperms and from endosperm in angiosperms), and
the seed coat (derived from the integument of the ovule).
Problem : What is the female reproductive organ of a flowering plant, and where
must pollen land in order to fertilize the eggs it contains?
Pistils are the female reproductive organs of angiosperms, and pollen grains
must land on their stigmas, or tips, in order to reach the egg cells.
Problem : Are ovaries characteristic of angiosperms or gymnosperms? What are
their functions?
Ovaries, found in angiosperms, enclose the ovules and develop into fruits after
fertilization. Ovaries protect the embryos from drying out and help in their
dispersal.
Problem : How is cross-pollination encouraged in most angiosperms?
Cross-pollination is encouraged through differential development of the male
and female gametophytes on a flower, or through the positioning of these
gametophytes so that self-pollination is difficult.
Through asexual reproduction, many plants can produce genetically identical
offshoots (clones) of themselves, which then develop into independent plants.
This process is called vegetative propagation, or vegetative reproduction. One
way in which vegetative propagation occurs is through fragmentation, a process
in which a severed plant part can grow into a whole new plant. Other modes of
vegetative propagation include the production of specialized structures such as
tubers, runners, and bulbs. The advantages to this kind of asexual reproduction,
which can occur either naturally or artificially, stem from the fact that it can
occur more rapidly than seed propagation and can allow a genetically superior
plant to produce unlimited copies of itself without variation.
Tubers
Tubers, such as potatoes, are fleshy underground storage structures composed
of enlarged parts of the stem. A tuber functions in asexual propagation as a result
of the tiny scale leaves equipped with buds that grow on its surface. Each of
these buds can form a new plant, genetically identical to the parent.
Runners
Runners, such as those found on strawberry plants, are slender horizontal stems
that spread outward from the main plant . Entirely new plants can develop from
nodes located at intervals on the runners; each node can give rise to new roots
and shoots.
Bulbs
Bulbs, such as onions and tulips, are roughly spherical underground buds with
fleshy leaves extending from their short stems. Each bulb contains several other
buds which can give rise to new plants.
Problem : Explain the difference between seed propagation and vegetative
propagation.
With seed propagation, reproduction occurs through embryos (contained in
seeds) that are produced sexually. With vegetative propagation, by contrast,
plants reproduce asexually through genetically identical offshoots (clones),
which then develop into independent plants.
Problem : What are the advantages to vegetative propagation in comparison
with seed propagation?
Vegetative propagation can occur more rapidly than seed propagation and can
allow a genetically superior plant to produce unlimited copies of itself without
variation.
Problem : What is the name for the process by which a severed plant part grows
into a whole new plant?
Fragmentation.
Problem : List three kinds of specialized structures developed by some plants to
allow vegetative propagation.
Tubers, runners, and bulbs.
Problem : What occurs at the nodes of runners?
Each node can give rise to new roots and shoots, so that an entirely new plant
can develop.
Grafting is an artificial form of vegetative propagation in which parts of two
young plants are joined together, first by artificial means and then by tissue
regeneration. Typically, a twig or bud is cut from one plant and joined to a rooted
plant of a related species or variety. The twig or bud is called the scion, and the
plant onto which is it grafted (and that provides the roots) is called the stock.
The scion eventually develops into an entire shoot system. Grafting often allows
horticulturalists to combine the best features of two different plants into one
plant. Sometimes the stock and scion retain independent characteristics, and
sometimes the stock alters the characteristics of the scion in some desirable way.
Problem : What is grafting?
Grafting is an artificial form of vegetative propagation in which parts of two
young plants are joined together, first by artificial means and then by tissue
regeneration.
Problem : From a horticultural standpoint, what is the purpose of grafting?
Grafting often makes it possible to combine the best features of two different
plants into a single plant.
Problem : When parts of two plants are grafted together, what is the rootproviding plant called?
The stock.
Problem : When parts of two plants are grafted together, what is the rootless
twig or bud called?
The scion.
Problem : Do the characteristics of the stock always affect those of the scion?
No; although this is sometimes the case, it is possible for the scion to retain
independent characteristics after being grafted onto a stock with different
characteristics.