Survey
* Your assessment is very important for improving the work of artificial intelligence, which forms the content of this project
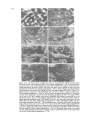
Clinical Science ( 1 989) 76, 269-276 269 Multi-organ damage resulting from experimental faecal peritonitis D. TIGHE, R. MOSS, S. BOGHOSSMN, M. F. HEATH, B. CHESSUM AND E. D. BENNETT Departments of Medicine 1, Electron Microscopy, Haematology and Microbiology,St Georges Hospital Medical School, London, and Department of Clinical Veterinary Medicine, University of Cambridge, Cambridge, U.K. (Received 25 January/29 April 1988; accepted 11 July 1988) SUMMARY 1. Using specific-pathogen-free New Zealand White rabbits, we have compared the effects of faecal peritonitis over a period of 5 h in eight test animals with eight controls in which a sham operation was performed. 2. There was morphological damage to lungs, liver and spleen of test animals. Lung capillaries and sinusoids of the liver showed occlusion by cell debris and leucocytes, with endothelial damage. The lungs also showed alveolar epithelial disruption, basement membrane exposure and type I1 pneumocytes lacking lamellar bodies. In the liver there was fibrin deposition and swollen Kupffer cells. The spleen showed degranulating neutrophils, fibrin deposits, p!atelet aggregates and activated macrophages, with no damage to the endothelium. 3. There was no morphological damage to the kidney or heart of test animals or to any organs of sham-operated animals. 4. There were mixed anaerobes and aerobes in faecal material used to induce peritonitis. Cultures of liver, spleen and kidney isolated four different types of micro-organisms. Blood cultures showed two types of micro-organisms. Cultures of lung and heart showed one type of micro-organism. 5. The presence of micro-organisms in an organ C O U ! ~ not be correlated with the degree of histological damage to that organ. 6. In test animals an early significant reduction in circulating leucocytes and platelets was sustained for the duration of the experiment with significant diffuse intravascular coagulation. 7. There was no change in test animal neutrophil adhesiveness until 120 min, when significant reduction was observed. 8. Serum phospholipase A, (EC 3.1.1.4) activity in the test group showed a threefold increase at 300 min. Correspondence: Dr D. Tighe, Department of Medicine 1, St Georges Hospital Medical School, Cranmer Terrace, London SW17 O R E . Key words: histological change, multi-organ damage, neutrophil adhesiveness, peritonitis, phospholipase A,. Abbreviations: ARDS, adult respiratory distress syndrome; LM, light microscopy; MABP, mean arterial blood pressure; PLA,, phospholipase A, (EC 3.1.1.4); SEM, scanning electron microscopy; TEM, transmission electron microscopy. INTRODUCTION Multi-organ failure associated with septicaemia is a common clinical problem with a mortality rate approaching 100%. Anatomical and physiological changes occurring in the lung after sepsis [l,21 and endotoxic shock [3] have demonstrated leucostasis, endothelial damage and pulmonary hypertension. Scant attention has been paid, however, to damage that occurs in other organs. There is little documentation of the relationship between the presence of micro-organisms and tissue damage affecting various organs in bacteraemia. In addition, it is unclear whether the degree of organ damage is proportional to the number of micro-organisms present. Leucocytopaenia has been shown in animal models of endotoxic shock and may be related to leucostasis occurring in various organs. Such leucostasis may be due to increased neutrophil adhesiveness to the endothelium, although little work has been done to establish whether there is any such increase in septic shock in humans. Phospholipase A, (PLA,; E C 3.1.1.4) activates the arachidonic acid cascade, the products of which may be responsible for some of the changes seen in septic shock. Serum PLA, has been demonstrated to be elevated in patients with septicaemia [4] and in a rabbit model of endotoxic shock [5].We have shown previously that administration of mepacrine, a PLA, antagonist, markedly attenuates histological changes occurring in the rabbit lung after faecal peritonitis [l]. We undertook the present study using a rabbit peritonitis model described previously [l], to examine tissue 270 D. Tighe et al. changes in lungs, liver, kidney, heart and spleen by electron microscopy. Bacterial cultures of these tissue were performed to establish any correlation between tissue damage and the presence of micro-organisms. In view of the leucostasis that occurs in this animal model, we determined whether any increase in neutrophil adhesiveness took place. We also wished to confirm that serum PLA, activity would increase in this animal model. METHOD Faecal peritonitis was induced in eight specific-pathogenfree New Zealand White rabbits (2-2.5 kg) [l]. In brief, after anaesthesia a limited laparotomy was performed, the caecum opened, 5 ml of caecal contents removed and the intestinal incision closed. The caecal aspirate was then spread evenly around the peritoneal cavity and the laparotomy closed. A similar laparotomy was performed in the eight controls, but the caecum was not opened. Animals were killed after 300 min when samples of lung, kidney, liver, heart and spleen were taken for light microscopy (LM) and for transmission (TEM) and scanning electron microscopy (SEM). Tissue for TEM was fixed in 3% (v/v) glutaraldehyde in cacodylate buffer, post-fixed in 1% (w/v) osmium tetroxide in buffer, dehydrated in ascending concentrations of ethanol and embedded in Spurs' resin. Tissue for SEM was similarly fixed and dehydrated by alcohol and then taken through a freon alcohol series and critical point dried. Samples of lung, liver, heart, spleen, kidney, blood and caecal content were taken for bacteriological examination. The organ samples were dipped into boiling water for a few seconds to sterilize external surfaces and then the sample was cut in half. The size of the surface was noted and was then smeared over the surface of the plates of media. Cultures were incubated for 48 h in 10% CO, in air and anaerobically with 10% CO, for 48 h. Blood samples were taken and cultured using a poured plate technique with 0.1,0.5 and 1.0 ml of blood per plate. The plates were incubated both anaerobically and aerobically with 10% CO,. Three millilitres were also cultured in a Casteneda and a thioglycollate blood culture broth bottle. These two bottles were subcultured at 2 days and 5 days after inoculation on both aerobic and anaerobic media. Blood samples were taken at baseline, 30,60, 180 and 300 min for total and differential leucocyte counts, basic clotting parameters and neutrophil adhesion to nylon fibre in a Pasteur pipette by the method of MacGregor el aI. [6]. Fibrinogen was assayed by the method of Clauss [71. Serum PLAz levels were estimated on clotted plasma at baseline and 300 min. The method was a modification of those of Heath & Jacobson [8,9].The substrate used was 1-palmitoyl 2-[ 1-14C]oleoylphosphatidylcholine (Applied Science, Oud-Beijerland, Netherlands) diluted in egg phosphatidylcholine (Sigma, Poole, U.K.) to give 50 000 d.p.m. per assay tube, dispersed by brief ultrasonication at a concentration of 0.8 mmol/l in the assay buffer: 50 mmol/l 4-( 2-hydroxyethy1)-1-piperazine-ethanesulphonic acid/HCl (pH 7.5 at 37"C), 10 mmol/l CaCI,, 0.1% (w/v) bovine serum albumin. To 20 pl of rabbit serum were added 80 pl of sodium deoxycholate (5 mmol/l, in 150 mmol/l NaCI), and 100 pl of substrate. The assay tubes were incubated for 1 h at 3 7 T , then acidified with 25 pl of 0.5 mol/l H,SO,, and extracted with chloroform/methanol (containing 25 p g of oleic acid) by the procedure of Bligh & Dyer [lo]. Fifty microlitres of the chloroform layer was analysed by t.1.c. on LK6D silica gel plates (Whatman, Maidstone, U.K.),developed first with chloroform/methanol/water (65:25:4, v/v), then after drying, with hexaneldiethy1 ether/acetic acid (69:29:2, v/v). Iodine vapour was used to locate the oleic acid and phosphatidylcholine spots, which were scraped off for scintillation counting. PLA, activity was calculated from the proportion of radioactivity recorded in the fatty acid spot, and adjusted for the activity found with reagent blanks (20 p1 of NaCl instead of serum). Results are expressed as means fSEM. Statistical significance was evaluated by Student's r-test. RESULTS Histological analysis Samples of lung tissue (Fig. 1) showed reduction in capillary lumen area, with marked leucostasis and capillary loop occlusion by neutrophils, lymphocytes and cell debris. Although platelets were seen, aggregates never formed. Neutrophils were adherent to the capillary endothelium which showed hypertrophy with occasional disruption and exposure of basement membrane. Alveolar epithelial cells showed swelling and some bleb formation. Type I1 pneumocytes appeared deranged, having a reduced number of lamellar bodies. Neutrophils showed progressive degranulation of primary granules with the development of translucent vacuoles, many of which had lost their membranes. Areas of pooled glycogen and phagosomes containing bacteria were also seen. There was no interstitial oedema or fibrin deposition. Liver samples (Fig. 2) revealed dense occlusion of the sinusoids by degranulating neutrophils, lymphocytes, cell debris and fibrin deposits. Rod-like bacteria were present in the lumen of the sinusoids, some being phagocytosed by neutrophils. Kupffer and endothelial cells showed hypertrophy with consequent narrowing of the sinusoidal lumina and the space of Disse, with a reduction in sinusoidal luminal area. SEM revealed the sieve plates of the sinusoidal endothelium to be disrupted by areas of larger fenestrations, especially when adjacent to activated Kupffer cells. Most of the occlusion by leucocytes, Kupffer cells, endothelial cells and fibrin deposition occurred in sinusoids of zone 1 of the Rappaport acinus system. Sinusoids of zone 2 appeared constricted but with little damage, whilst the sinusoids of zone 3 appeared more patent where they entered the hepatic vein. Each acinus defines an area of liver tissue organized around the terminal branch of the portal vein in such a way that cells at the centre of the acinus, zone 1, are the first to receive blood, Multi-organ damage in rabbit peritonitis 27 1 Fig. 1. Lung. (a) Section of lung from a sham-operated animal showing erythrocytes (RBC)in patent thin-walled capillaries and thin interstitiurn. LM:original magnification X 550. ( b )Section from test animal showing capillary occlusion by polymorphonuclear leucocytes (PMN) and lymphocytes, with thickened interstitium. LM:original magmfication x 550. ( c )Septal wall from test animal showing degranulating PMN occluding capillaries, with endothelial and epithelial damage (arrows). T E M original magnification x 1700. ( d )Degranulating PMN with phagosome containing a bacterium, occluding a capillary within a thickened septa1 wall. TEM: original magnification X 6000. ( e )Fractured capillary loop from a sham-operated animal. SEM: original magnification X3400. (f)Fractured capillary loop from a test animal showing obstruction by an adherent PMN and RBC. S E M original magnification x 3400. followed by zone 2, and cells located toward the periphery and collecting vein, zone 3, being the last to receive blood. The spleen (Fig. 3) showed degranulating neutrophils in the red pulp, arterioles and venous sinuses. Many aggregates of platelets were seen in the red pulp, although there was no capillary occlusion. Macrophages showed evidence of activation and contained large amounts of phagocytosed cell debris. Fibrin was also found adjacent to the endothelium of the venous sinuses and red pulp. There was no damage to the endothelium. Heart and kidney histology showed no adherent leucocytes, endothelial disruption or fibrin deposition. There were no histological changes seen in any of the organs taken from the sham-operated animals. Microbiology Within the test group a number of different species of organisms were isolated. The caecal aspirate contained mixed aerobic and anaerobic bacteria to a count of at least 1.4 x loy per ml of 'emulsion'. The predominant species found were Bacteroides,Enterobacteriaceae and Streptococcusfaecalis. All blood cultures taken at baseline were sterile but those taken at 120 and 300 min grew Enterobacter agglomerans together with a Bacteroides species. Shamoperated animals did not have cultures taken. Cultures of heart and lung rarely produced more than a few colonies of E. agglomerans (Table 1). Cultures of liver, spleen and kidneys produced mixed growths of 272 Fig. 2. Liver. ( a ) Section from a portal area from a sham-operated animal showing patent sinusoids, several with erythrocytes (REX).LM: original magnification x 550. ( b )Section from a portal area from a test animal showing open sinusoids (*) followed by capillary obstruction from polymorphonuclear leucocytes (PMN), cell debris and fibrin (arrow) leading to more narrowed and obstructed sinusoid. LM: original magnification x 550. (c)Section from a test animal showing total obstruction of a sinusoid by a Kupffer cell (KC)and degranulating PMN (PMN).Fibrin ( F ) is present between obstructive cells. Endothelial cells and space of Disse cannot be clearly seen. TEM: original magnification X 1600. ( d )Section from a test animal showing fibrin ( F )deposition and cell debris occluding a sinusoid between enlarged endothelial cells (EC). The space of Disse can be seen at (D), but is mainly absent where endothelial cells flatten the microvilli on to the hepatocyte surfaces (arrow). TEM: original magnification x 1850. (e) Fractured surface from test animal showing sinusoidal lumen (SL), Kupffer cells (KC) and sinusoidal obstruction (Ob) by PMN, RI3C and cell debris. SEM: original magnification X 600. (f)Enlarged Kupffer cell (KC) obstructing sinusoidal lumina (SL). The endothelium close to Kupffer cells shows fenestrations larger than in other areas (arrows). Part of a hepatic chord (HC). SEM. original magnification X 2200. ( g ) Sinusoidal lumen from a sham-operated animal showing small endothelial fenestrations (FE). Hepatic microvillus surface (large arrow) of a hepatic chord (HC). Small arrow shows the bile canaliculi. SEM: original magnification x 3750. ( h )Sinusoidal lumen from a test animal showing large endothelial fenestrations through which the microvillus surface of the hepatocytes can be seen. SEM: original magnification X 3750. Multi-organ damage in rabbit peritonitis 273 Fig. 3. Spleen. ( a ) Section from a test animal showing a venous sinus (VS), red pulp area (Rp) with erythrocytes. Also present are many polymorphonuclear leucocytes (PMN) (P) and aggregates of platelets (PL).TEM: original magnification X 1100. ( 6 )Section from a test animal showing platelet aggregation, a macrophage (M) the remains of which surround a degranulating PMN (P).TEM: original magnification X 1700. ( c ) Fibrin deposit (F),reticular fibres (arrow) and macrophage (M) adjacent to venous sinus (VS). TEM: original magnification x 3150. ( d )Fibrin deposit showing characteristic banding. TEM: original magnification x 3500. Table 1. Microbiology and histology in test animals Micro-organisms: - ,no colonies; + ,colonies present. Tissue changes: + ,present, - ,absent. Lung Micro-organisms E. agglornerans S. faecalis Bacteroides Gram-positive anaerobic rods + Tissue changes Degranulating neutrophils Endothelial damage Fibrin deposits Disseminated intravascular coagulation Heart + Blood Liver Spleen Kidney + + + + + + + + + + + + + + - + + +- ++- - - - - - - + +- - - - + - - Table 2. Differential blood cell count Neutrophils, lymphocytes and platelets at time 0 and 300 min in test and sham-operated animals. Results are expressed as meant- SEM. Statistical significance: *P= 0.02, **P= 0.001. x Cell count (no. of cells/l) Time (min) Test animals Sham-operated animals 0 300 0 300 Neutrophils Lymphocytes 2.9 f 0.9 0.6+-0.1* 2.4 f 0.5 5.6 f 1.0* 2.4 f0.3 0.9 f 0.1 ** 3.4 f 1.2 3.3 f0.7 Platelets * 660 89 290 f 51** 700 f 82 710+55 D. Tighe et al. 274 Table 3. Blood clotting and PLA, Prothrombin time (PT), thrombin time (TT), kaolin partial thromboplastin time (KPTT), fibrinogen and PLA, at time 0 and 30 min in test and sham-operated animals. Results are expressed as mean k SEM. Statistical significance: *P= 0.05, **P= 0.01. Clotting time (s) Time (min) PT TT KPTT Test animals 0 300 5.2k0.3 6.5 k 0.5* 12.5k0.8 20.6 k 3.7 Sham-operated animals 0 300 5.5k0.3 5.3k0.4 12.1k1.6 10.9k1.8 E. agglomerans, S. faecalis, two types of Bacteroides species and a Gram-positive anaerobic rod. Haematology Peripheral neutrophil count in the test animals (Table 2 ) fell significantly to 19% of the baseline at 300 min. In the sham-operated rabbits there was a significant increase to 133% of baseline at 300 min. Peripheral lymphocyte count in the test animals fell significantly to 37.5% of baseline at 300 min. There was no significant change in the sham-operated group. Peripheral platelet count in the test animals fell significantly to 41% of baseline at 300 min. There was no significant change in the sham-operated group. Prothrombin time in the test group (Table 3) at 300 min increased significantly to 125% of baseline. The thrombin time increases to 165% and kaolin partial thromboplastin time to 145% were not significant. Plasma fibrinogen fell significantly to 69%. Plasma fibrinogen in the sham-operated animals fell significantly to 93%; the other parameters showed no significant change. Neutrophil adhesion in the test animals 30 min after peritonitis induction fell to 64% of baseline, 68% after 60 min and significantly to 74% after 120 min (Fig. 4). The sham-operated animals showed no significant changes. Serum PLA, activity in the test group showed a significant increase to 383% of baseline after 300 min (Table 3). In the sham-operated group there was a fall to 56% of baseline. DISCUSSION We have developed a rabbit model of faecal peritonitis simulating clinical peritonitis, a condition often associated with the development of multi-organ failure and the adult respiratory distress syndrome (ARDS). We have demonstrated an early and maintained reduction in peripheral blood leucocyte and platelet counts. There was evidence of progressive disseminated intravascular coagulation with significant reduction of plasma fibrinogen, although fibrin degradation products were not measured. These results are similar to those obtained by other groups using a variety of techniques [2,3]. 120 8 g .- Fibrinogen (dl) PLAz (i.u./l) 44.5k4.7 64.0 f 12 3.4k0.5 2.3 k 0.3** 0.9 k 0.5 3.6 rt 1 .O* 40.5k4.6 36.0k5.1 2.9k0.1 2.7kO.l" 1.8 k 0.7 1.Ok0.1 - 100- ao- M c 2 6040 20 - 1 I I I We have also demonstrated the expected histological abnormalities in the lungs showing capillary leucostasis, endothelial disruption and type I1 pneumocyte damage [2, 31. The changes shown in the liver and spleen have been reported less frequently but are similar to the findings of Balk et al. [2] and Coalson et al. [3]. In the liver there was fibrin deposition with evidence of severe damage to the sinusoids of zone 1 of the hepatic acinus but none to the hepatocytes or bile canaliculi. The spleen showed evidence of fibrin deposition, activation of tissue macrophages and aggregates of degranulating neutrophils but with no endothelial disruption. In marked contrast, the heart and kidney did not show any of these changes. Coalson et al. [3]were also unable to show leucostasis in these organs of their septic baboon model. There was heavy polymicrobial growth from the caecal aspirant and micro-organisms were cultured from blood. Polymicrobal growths of similar micro-organisms were found in the liver, spleen and kidneys, although the lungs and heart showed only a few colonies of one species. These findings strongly suggest that there is no direct relationship between the presence of micro-organisms in an Multi-organ damage in rabbit peritonitis organ and the degree of damage to that organ (Table 1). Although liver and spleen had polymicrobal growth with severe histological damage, this relationship was not true of the lungs where only one species was isolated but again with severe histological damage. The kidney with the presence of polymicrobial growth and heart where only one species was isolated showed no evidence of histological change. We should emphasize that there could have been functional changes in the organs not reflected by histological damage. These findings are compatible with those of Goris et al. [ll], who produced multi-organ damage in rats by inducing sterile peritonitis with zymosan. These data show that organ damage is likely to be produced by mediators released as a result of the peritoneal inflammation. This poses the question as to why leucocytes aggregate in the liver, lungs and spleen but not in other organs. We were unable to show any increase in neutrophil adhesiveness to account for leucostasis; indeed, we demonstrated a decrease that became significant at 120 min in those neutrophils still circulating in the peripheral blood. It may be postulated that those cells aggregating in the organs do, however, show an increase in adhesiveness which we were unable to measure. It is also possible that changes occur in endothelial adhesiveness modifying the relationship between the activated leucocytes and the endothelium. Fowler et al. [12] showed that there was an absence of any humoral factor in the blood of ARDS patients capable of causing chemotaxis or enzyme secretion in normal neutrophils. Zimmerman et al. [13] showed that plasma from ARDS patients was unable to promote a respiratory burst from healthy neutrophils. We have shown a fourfold increase in serum PLA, in response to experimental faecal peritonitis, this change in circulating activity being concurrent with the haematological and multi-organ histological damage. Infusion of endotoxin has been found to produce an 11-fold increase in PLA, [5]. Infusions of a rabbit plasma fraction containing PLA, caused a small effect on mean arterial blood pressure (MABP) which could be prevented by pretreatment of the material with an inhibitor of PLA, [5]. Vadas & Hay [5] suggested that circulating PLA, releases fatty acids and lysophosphatides from phospholipids and that these, and their metabolites, are responsible for both hypotension and margination of leucocytes. The data showing that MABP starts to fall before any significant change occurs in circulating PLA, [5] offers an alternative hypothesis for the role of elevated serum PLA,. It suggests that PLA, is secreted and has a local action while still at a high concentration, before some or all of the secreted enzyme escapes into the circulation. To attain high tissue concentrations secondarily, by diffusion out of the circulation, seems less likely. The serum level of PLA, would thus indicate escape of PLA, from the tissues, and would depend on (i)the rate of secretion within the tissues, and (ii)the rate of clearance into the circulation. Increased mediator production (e.g. prostaglandins) would result from an increase in PLA, secretion, or a decrease in clearance of PIA,. On the other hand, an 275 increased serum PLA, level would arise with an increase in secretion or an increase in clearance into the circulation. This may not therefore be a reliable index of mediator production, and so of potential damage and clinical prognosis. Elevation of serum PLA, has been found in patients at risk for multi-organ failure [l],the levels showing some correlation with prognosis. Absolute levels in humans are very much lower than those in rabbits [8],the peak in severe human cases after several days of illness being only 15% of the level measured in rabbits 5 h after endotoxin infusion. Preliminary results from another group [ 14, 151 show similar clinical correlations, but much greater absolute values. Assay methods used by this group of investigators were very different, thus comparisons may not be meaningful. Although the eicosanoids are important in the production of inflammation, cytokine polypeptides also play a role in this process. Thus micro-organisms, endotoxin, C5a and other agents induce inflammation by the release of the mediators interleukin-1 [16] and tumour necrosis factor [ 171. These cytokines, produced by mononuclear leucocytes, can stimulate neutrophil activation and adherence to capillary endothelium and can stimulate endothelium to produce more interleukin-1 and platelet-activating factor [ 181that can also stimulate neutrophils. This study has therefore shown that the production of faecal peritonitis in rabbits results in severe damage to lungs, liver and spleen with marked leucostasis within these organs. In contrast, the heart and kidney showed no evidence of damage. Furthermore, the presence of microorganisms in an organ is not a prerequisite for histological damage. There was the expected reduction in peripheral leucocytes and platelets with evidence of diffuse intravascular coagulation. There was a significant reduction in neutrophil adhesiveness. Although we have demonstrated a marked rise in serum PLA, activity, it is uncertain whether this is a cause or effect of the organ damage that we found. REFERENCES 1. Tighe, D.,Moss, R., Parker-Williams,J., Hynd, J. & Bennett, E.D. (1987) A phospholipase inhibitor modifies the pul- monary damage associated with Peritonitis in rabbits. Infensive C&e M e d h e , 13,284-296. 2. Balis, J.U., Rappaport, E.S.,Gerber, L. & Buddingh, F. (1974) Continuous endotoxemia in Rhesus monkeys as a clinically relevant model for shock lung. American Journal of Pathology, 74,90-95. 3. Coalson, J.J., Hinshaw, L.B., Guenter, C.A., Berrell, E.L. & Greenfield, L.J. (1975) Pathophysiologic responses of the subhuman primate in experimental septic shock. Laboratory Invesfigation,3 2 , 5 6 1-56 9. 4. Vadas, P. (1984) Elevated plasma phospholipase A, levels: correlation with the hemodynamic and pulmonary changes in Gram-negative septic shock. Journal of Laboratory and Clinical Medicine, 104, 873-881. 5. Vadas, P. & Hay, J.B. (1983) Involvement of circulating phospholipase A, in the pathogenesis of the hemodynamic changes in endotoxin shock. Canadian Journal of Physiology and Pharmacology, 61,561-566. 6. MacGregor, R., Spagnuolo, P. & Lentnek, A. (1974) In- 276 7. 8. 9. 10. 11. 12. D. Tighe et al. hibition of granulocyte adherence by ethanol, prednisolone and aspirin. Measured with an assay system. New England Journal of Medicine, 291,642-646. Clauss, A. (1957) Rapid physiological coagulation method in determination of fibrinogen. Acta Haematologica (Basel), 17,237-241. Heath, M. & Jacobson, W. (1984) Developmental changes in enzyme activities in foetal and neonatal rabbit lung. Cytidylyl transferase, cholinephosphotransferase, phospholipase A , and A?, B-galactosidase and B-glucuronidase. Pediatric Research, 18,395-401. Heath, M. & Jacobson, W. (1985) The inhibition of lysosoma1 phospholipase Az from rabbit lung by ambroxol and its consequences for pulmonary surfactant. Lung, 163, 337-344. Bligh, E. & Dyer, W. (1959) A rapid method of total lipid extraction and purification. Canadian Journal of Biochemical Physiology, 3 7 , 9 1 1 -9 1 7. Goris, J., Boekholz, W., van Bebber, I., Nuytinck, J. & Schilling, P. (1986) Multiple-organ failure and sepsis without bacteria. Archives ofSicqery, 121,897-901. Fowler, R.A., Fisher, B.J., Centor, R.M. & Carchman, R.A. (1984) Development of the adult respiratory distress syndrome: progressive alterations of neutrophil chemotactic 13. 14. 15. 16. 17. 18. and secretory processes. American Journal of I’athology, 116,427-435. Zimmerman, G., Renzetti, A. & Hill, H. (1983) Functional and metabolic activity of granulocytes from patients with adult respiratory distress syndrome. American Review of Respiratory Diseases, 127,290-300. Hoffmann, G.E., Schmidt, D. & Bastian, B. (1985) Bestimmung der phospholipase A im serum bei akuter pankreatitis. Journal of Clinical Chemistry and Clinical Biochemistry, 23,582-583. Koeniger, R., Staffa, F., Hoffmann, G.E., Schmid, T.-0. & Guder, W.G. ( 1 986) Automated analysis of phospholipase A activity in serum of intensive care patients. Fresenius’ Zeitschrifficer Analytische Chemie, 324,308. Dinarello, C.A. (1984) Interleukin-1. Review of Infectious Diseases, 6,51-95. Mathews, N. ( 1 978) Tumour-necrosis factor from the rabbit. 11. Production by monocytes. British Journal of Cancer, 38,310-315. Sauder, D.L., Maunessa, N.L., Katz, S.I., Dinarello, C.A. & Gallin, J.I. ( 1 984) Chemotactic cytokines: the role of leukocytic pyrogen and epidermal cell thrombocyte activating factor in neutrophil chemotaxis. Journal of lrnrniinology, 132,828-832.