Survey
* Your assessment is very important for improving the workof artificial intelligence, which forms the content of this project
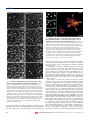
articles Antagonistic microtubule-sliding motors position mitotic centrosomes in Drosophila early embryos David J. Sharp*, Kristina R. Yu†, John C. Sisson†, William Sullivan† and Jonathan M. Scholey*‡ *University of California at Davis, Section of Molecular and Cellular Biology, 1 Shields Avenue, Davis, California 95616, USA †University of California at Santa Cruz, Department of Biology, 317 Sinsheimer Labs, Santa Cruz, California 95064, USA ‡e-mail: [email protected] The positioning of centrosomes, or microtubule-organizing centres, within cells plays a critical part in animal development. Here we show that, in Drosophila embryos undergoing mitosis, the positioning of centrosomes within bipolar spindles and between daughter nuclei is determined by a balance of opposing forces generated by a bipolar kinesin motor, KLP61F, that is directed to microtubule plus ends, and a carboxy-terminal kinesin motor, Ncd, that is directed towards microtubule minus ends. This activity maintains the spacing between separated centrosomes during prometaphase and metaphase, and repositions centrosomes and daughter nuclei during late anaphase and telophase. Surprisingly, we do not observe a function for KLP61F in the initial separation of centrosomes during prophase. Our data indicate that KLP61F and Ncd may function by crosslinking and sliding antiparallel spindle microtubules in relation to one another, allowing KLP61F to push centrosomes apart and Ncd to pull them together. entrosomes are the primary microtubule-organizing centres within animal cells and thus they play a fundamental part in numerous cellular and developmental events. For example, during mitosis, duplicated centrosomes form the bipolar mitotic spindle poles that determine the position and orientation of cleavage planes which, in turn, determine the sizes of daughter cells and spatial relationships that exist between them1. Moreover, in multinucleated cells, such as the Drosophila syncytial blastoderm, the relative positioning of centrosomes also contributes to the spacing of multiple adjacent nuclei2. Work in fungal and animal systems3 indicates that the microtubule-associated motors termed bipolar and carboxy-terminal kinesins4 may be important in determining the relative position of adjacent centrosomes, particularly during mitosis3–25. For example, severe loss-of-function mutations in the gene encoding the Drosophila bipolar kinesin, KLP61F, result in the formation of mono-astral mitotic spindles with duplicated but closely spaced centrosomes, leading to the proposal that this bipolar kinesin participates in the initial separation of centrosomes during mitotic spindle assembly13–17. This suggestion, however, is difficult to reconcile with our recent observation that KLP61F motors are sequestered in the nucleus during prophase when the centrosomes initially separate17. A mitotic function has also been proposed for the Drosophila C-terminal kinesin, Ncd, on the basis of the abnormal morphology of the mitotic spindles that form in its absence22, but the precise mitotic function of this motor remains unclear. Here we test the hypothesis that KLP61F and Ncd position mitotic centrosomes by generating opposing microtubule-sliding forces within interpolar microtubule bundles17 and telophase midbodies2. C Results Spindles assemble but collapse after loss of KLP61F function. To investigate the function of KLP61F in positioning centrosomes, we used the Drosophila early embryo system to study the effects of injecting antibodies against functionally essential epitopes found in the tail of KLP61F (ref. 17) into embryos containing fluorescently labelled tubulin and chromatin (Fig. 1). This allowed us to assess the influence of KLP61F on centrosome positioning in real-time by simultaneously visualizing nuclei (data not shown) and the microtubules emanating from centrosomes (Fig. 1). NATURE CELL BIOLOGY | VOL 1 | MAY 1999 | cellbio.nature.com During mitosis in control injected embryos, sets of duplicated centrosomes separated by about 120° to nearly opposite sides of the nucleus, at a rate of roughly 5 µm min-1. As the nuclear envelope broke down at the onset of prometaphase, microtubules from opposite centrosomes began to fill the nuclear region and intercalate to form interpolar bundles. Over the next minute, the metaphase spindle was established and then maintained for 2–3 minutes before the onset of anaphase. During this time the relative position of centrosomes within mitotic spindles did not change substantially. In embryos injected with anti-KLP61F antibodies, the movement of centrosomes in relation to one another during early spindle assembly appeared unaffected and duplicated centrosomes separated at a rate nearly identical to that of controls (~5.5 µm min-1). The onset of prometaphase, as marked by the appearance of microtubules within nuclei and the formation of interpolar microtubule bundles, was also relatively normal. However, shortly after this the effects of KLP61F inhibition become apparent. Instead of forming stabilized bipolar metaphase spindles as in controls, separated centrosomes quickly slid back together, at a rate of 5–7 µm min-1, in the absence of KLP61F activity. Within 3 minutes after nuclear-envelope breakdown, nearly all the centrosomes that previously constituted the two spindle poles had collapsed together and appeared as small ‘mono-asters’ of microtubules. Analysis of multiple focal planes revealed that chromosomes remained attached at the distal ends of the centrosomal microtubules, which extend down into the interior of the embryo (data not shown). These data indicate that KLP61F may not function during the initial separation of centrosomes during prophase, as previously thought (although a redundant role in this process or the inaccessibility of specific antigenic sites on KLP61F molecules during prophase cannot be ruled out), and show, unequivocally, that KLP61F is essential for maintaining the spacing between centrosomes within mitotic spindles after they have first separated. The inhibition of KLP61F also affects the positioning of centrosomes between mitotic nuclei, because once the collapse of individual spindles was complete, adjacent collapsed spindles began to slide together ( Fig. 2). This sliding was preceded by the formation of interspindle microtubule bundles (Fig. 2b, small arrowheads), which were quite similar to the microtubule bundles that form © 1999 Macmillan Magazines Ltd 51 articles Figure 2 Mitotic centrosomes associated with adjacent spindles interact through microtubule bundles to slide together and form large arrays of collapsed mono-asters in the absence of KLP61F function. Time series of tubulin fluorescence, showing the interactions between centrosomes from adjacent spindles following the spindle collapse that results from injection of anti-KLP61F antibodies. a–c, Centrosomes from adjacent collapsed spindles (a) become bridged by bundles of microtubules (b, small arrows) and slide together (c). Sets of centrosomes that are not bridged by microtubule bundles remain separate (a–c, large arrows). d, Over time, large numbers of spindles coalesce to form large microtubule monoasters. Tubulin is shown in yellow and chromosomes in blue. Note how the chromosomes remain attached to the distal ends of the microtubules emanating from these asters. The times shown at the bottom right of each panel are measured as in Fig. 1. Similar results were obtained with at least 15 different embryos. Scale bar represents 7 µm in a–c and 15 µm in d. Figure 1 Centrosomes within bipolar spindles separate but then collapse together following the inhibition of the bipolar kinesin KLP61F. Time series of images showing tubulin fluorescence during mitotic spindle formation, in Drosophila embryos injected with anti-KLP61F antibodies (right panels) or with nonspecific rabbit IgG (left panels) at the same concentration as a control. Arrows follow the centrosomes in two individual spindles. Times shown at the bottom right of each panel are the times from the onset of prophase. Centrosome separation during prophase occurs similarly in both conditions, but as metaphase spindles are established in controls the centrosomes slide back together in embryos injected with anti-KLP61F antibodies. Similar results were obtained in more than 20 different embryos in 5 experiments. Drosophila melanogaster is an ideal system for real-time functional analyses of mitosis31. Most important, Drosophila early embryos are large, multinucleated cells that undergo rapid and synchronous mitoses; thus the factors that position adjacent centrosomes both within and between mitotic spindles can be studied easily2,32. between the spindle poles during prometaphase. As collapsed spindle mono-asters that were not bridged by microtubule bundles did not slide together (Fig. 2, large arrowheads), the forces driving the 52 centrosomes together appear to require microtubule–microtubule interactions. The rate at which this occurs, ~6 µm min-1, is similar to the rate of spindle collapse, indicating that a similar mechanism may drive both. Over time, the coalescence of multiple adjacent centrosomes led to the formation of extremely large microtubule asters, with numerous centrosomes at their centres and condensed chromosomes attached to the distal ends of the microtubules (Fig. 2d). Although this phenotype indicates a probable metaphase arrest, chromosomes associated with these asters showed a pattern of phosphorylation of histone H3 that is characteristic of anaphase in wild-type cells (data not shown and ref. 26), a result that is being studied at present. The collapse of centrosomes within and between spindles resulted in the production of superficially similar mono-astral microtubule arrays and associated circular mitotic figures. Similar structures resulting from the inhibition of bipolar kinesins in mitotic extracts12, antibody-injected cells18 and mutants5–10,13 have been widely interpreted as resulting from defects in the initial separation of centrosomes, yet our data show that they could also arise from the collapse of preassembled bipolar spindles. Several other mitotic proteins have been assigned functions in the initial separation of centrosomes on the basis of similar phenotypes; our results indicate that the functions of these proteins should be re-evaluated. The effects of Ncd counterbalance those of KLP61F. The rate at which adjacent centrosomes slide together in the absence of KLP61F activity and the apparent requirement for interdigitating microtubules indicate that, in wild-type cells, a driving force may be generated by another microtubule-crosslinking and-sliding motor with the opposite transport properties to those of KLP61F. The C-terminal kinesin Ncd is a microtubule crosslinker that is directed to microtubule minus (more slowly growing) ends; it moves at a rate of about 8–15 µm min-1 in vitro19,20 and is therefore an attractive candidate for counterbalancing KLP61F activity. In © 1999 Macmillan Magazines Ltd NATURE CELL BIOLOGY | VOL 1 | MAY 1999 | cellbio.nature.com articles a Isometric tension b Collapse Centriole Ncd Microtubule KLP61F Figure 4 The relative positioning of mitotic centrosomes is determined by a balance of microtubule–microtubule sliding motors. The motors KLP61F and Ncd crosslink antiparallel microtubules in interpolar microtubule bundles. The force generated by KLP61F, a plus-end-directed motor, pushes centrosomes apart while Ncd, a minus-end-directed motor, pulls them together. a, Normally, during prometaphase and metaphase, these forces are counterbalanced, maintaining the spindle under isometric tension. b, If this balance is tipped, following the inhibition of KLP61F by antibody injections, the repulsive force between centrosomes is dissipated, the inward force generated by Ncd predominates, and centrosomes slide together. Small arrows show the direction of the force generated by individual motors on microtubules and large arrows show the direction of the resulting forces on centrosomes. Figure 3 Positioning of mitotic centrosomes and daughter nuclei involves an interaction between bipolar kinesins and C-terminal kinesins. Mitotic-spindle formation in embryos carrying a null allele (cand) for the C-terminal kinesin Ncd and injected with anti-KLP61F antibodies. a, Metaphase spindles assemble but are abnormally positioned in relation to one another. ‘Spurs’ or bundles of microtubules can also be seen running between adjacent spindles. b, The chromosomes segregate and early anaphase B occurs. Arrows indicate the spindle midbodies, consisting of interdigitating microtubules that span between the separating chromosomal masses. c, During later phases of anaphase B, these microtubule bundles shorten significantly and fray as centrosomes and daughter nuclei move back together. The inset shows the midbodies in wild-type embryos during the same stage of mitosis. These are longer and the microtubules within them are more organized. Daughter nuclei were never observed to slide together in either wild-type or cand controls. Similar results were obtained from >10 embryos and spindle collapse was never observed as either a direct or an indirect consequence of injection of anti-KLP61F antibodies into cand embryos. Scale bar represents 12.5 µm in a–c and 13.5 µm in the inset. support of this proposal are the findings that Ncd becomes concentrated on interpolar microtubule bundles within spindles in vivo22, as does KLP61F (ref. 17), and that null mutations in the gene encoding Ncd result in abnormally long and unstable spindles21,22 that could arise from defects in spindle-pole positioning. Genetic interactions between bipolar and C-terminal kinesins have been reported in fungi23–25. To determine whether KLP61F and Ncd do, in fact, generate NATURE CELL BIOLOGY | VOL 1 | MAY 1999 | cellbio.nature.com opposing forces between mitotic centrosomes, we injected embryos carrying a null allele for Ncd (termed cand; ref. 27) with the antiKLP61F antibodies (Fig. 3). In striking contrast to results obtained in wild-type embryos, the inhibition of KLP61F in an Ncd-null background did not inhibit the formation of bipolar metaphase spindles (Fig. 3a). Although the spindles that formed under these conditions appeared to be randomly positioned, and abnormal bundles of microtubules extended between them, they did not collapse but instead successfully completed anaphase A and appeared to be relatively normal during early anaphase B (Fig. 3b). During the later stages of anaphase B and telophase, however, marked defects in the organization of microtubules within the spindle midbody became visible, and the spacing between the centrosomes associated with daughter nuclei decreased substantially (Fig. 3c). These observations lead us to draw two conclusions. First, because genetic deletions of Ncd suppress spindle collapse and the subsequent formation of microtubule mono-asters that otherwise occur in the absence of KLP61F function, we conclude that KLP61F and Ncd generate opposing forces to determine the relative positioning of centrosomes within spindles during prometaphase and metaphase. Second, as the inhibition of both KLP61F and Ncd results in spindle defects during late anaphase B and telophase that affect the relative positioning of daughter nuclei, we conclude that the activity of these motors also helps to determine internuclear spacing within the early embryo. Thus, the forces generated by KLP61F and Ncd are not important solely for mitosis; they may also influence other facets of development, such as nuclear positioning. © 1999 Macmillan Magazines Ltd 53 articles Discussion RECEIVED 10 MARCH 1999; REVISED 31 MARCH 1999; ACCEPTED 1 APRIL 1999; PUBLISHED MAY 1999. Our results provide, to our knowledge, the first evidence that bipolar and C-terminal microtubule-sliding motors interact to generate antagonistic forces that position centrosomes during animal development. However, it is likely that other motors are also important in this process. For example, studies of several systems implicate the minus-end-directed motor cytoplasmic dynein in the initial separation of centrosomes during prophase28 and in spindle elongation during anaphase B29. Our findings also clarify the mitotic function of Ncd and reveal that this motor functions together with the bipolar kinesin KLP61F to position centrosomes during stages of mitosis after prophase (that is, following the initial separation of centrosomes, which appears to occur by a KLP61F- and Ncd-independent mechanism). These observations, together with the previous findings that KLP61F and Ncd are both able to crosslink microtubules, are similarly localized to interpolar microtubule bundles, and generate motility in opposite directions along the surface lattice of the microtubule, lead us to propose that these motors position centrosomes by using a ‘sliding-filament mechanism’30 to generate force between antiparallel microtubules (Fig. 4). When these antagonistic forces balance one another, centrosome spacing is maintained under isometric tension. However, tipping this balance results in movements of centrosomes in the direction of the prevailing motor.h Methods Drosophila stocks. We used Drosophila mutant stocks. Flies carrying the histone–green fluorescent protein (GFP) construct were provided and maintained by the Sullivan laboratory. Claret non-disjunctional (Ncd) flies (denoted cand) that are a genetic null for Ncd were provided by R. Scott Hawley. Embryo collections. Adult flies were maintained at 25 °C in small plastic containers inverted over agar dishes. To obtain embryos at the correct stage of development (during the formation of the syncytial blastoderm), flies were allowed to lay on fresh agar trays for 1 h. These trays were then removed and allowed to age for another hour at room temperature. Embryos were mechanically dechorionated on double-sided sticky tape and immobilized on heptane glue. Embryos were then dessicated for 5–9 min and injected under halocarbon oil. Microinjections. Microinjections of rhodamine-conjugated tubulin (Molecular Probes) were performed as described31. Embryos were then allowed to sit for 5 min and were then re-injected at the same site with either anti-KLP61F antibodies or non-specific rabbit IgG. The anti-KLP61F antibody injected was an affinitypurified peptide antibody against specific epitopes in the tail domain of KLP61F (ref. 17). Nonspecific rabbit IgG was obtained from the flow-through off the KLP61F affinity column. Both were concentrated to an identical optical density (OD 280) by spin filtration in all side-by-side experiments. Optimal effects of injections of anti-KLP61F antibody were observed at antibody concentrations between 7 and 12 mg ml-1. Injections of antibody at concentrations of <7 mg ml-1 produced only a marginal effect. Successive freeze-thawing of the KLP61F antibody diminished its effects. Because of this, antibodies were aliquotted, flash-frozen in liquid nitrogen, and stored at −80 °C shortly after their purification and concentration. Embryo preparations for anti-phospho-histone-H3 immunofluorescence. Following injections with anti-KLP61F antibody, embryos were aged at 25 °C for 15–40 min, washed free of halocarbon oil with 100% heptane, fixed in 100% heptane:37.5% formalin (1:1) for 5 min, handdevitellinized on double-sided sticky tape, and then incubated in 10 mg ml-1 RNase at 37 °C for 3 h. Standard antibody incubations and washes were performed. The antibodies used were: rabbit antiphospho-histone-H3 (1:500 dilution; a gift from T.-T. Su) and anti-rabbit-Cy5 (1:250 dilution, Chemicon). DNA was detected with propidium iodide. Laser-scanning confocal microscopy. All images shown were obtained on a Leica TCS NT confocal microscopes. Time series were generated using pre-set programs in Leica TCS software (xyt mode). In these, images were acquired either from the same optical section or from serial Z sections every 30–60 s over variable periods of time. 54 1. Rappaport, R. in Cytokinesis in Animal Cells (eds Barlow, P. W., Bard, J. B. L, Green, P. B. & Kirk, D. L.) 1–386 (Cambridge Univ. Press, Cambridge, 1996). 2. Sullivan, W. & Theurkauf, W. E. The cytoskeleton and morphogenesis of the early Drosophila embryo. Curr. Opin. Cell Biol. 7, 18–22 (1995). 3. Hoyt, M. A. & Geiser, J. R. Genetic analysis of the mitotic spindle. Annu. Rev. Genet. 30, 7–33 (1996). 4. Vale, R. D. & Fletterick, R. J. The design plan of kinesin motors. Annu. Rev. Cell Dev. Biol. 13, 745–777 (1997). 5. Enos, A. P. & Morris, N. R. Mutation of a gene that encodes a kinesin-like protein blocks nuclear division in Aspergillus nidulans. Cell 60, 1019–1027 (1990). 6. Roof, D. M., Meluh, P. B. & Rose, M. D. Kinesin-related proteins required for assembly of the mitotic spindle. J. Cell. Biol. 118, 95–108 (1992). 7. Hoyt, M. A., He, L., Loo K. K. & Saunders, W. S. Two Saccharomyces cerevisiae kinesin-related geneproducts required for mitotic spindle assembly. J. Cell. Biol. 118, 109–120 (1992). 8. Saunders, W. S. & Hoyt, M. A. Kinesin-related proteins required for structural integrity of the mitotic spindle. Cell 70, 451–458 (1992). 9. Hagan, I. & Yanagida, M. Novel potential mitotic motor protein encoded by the fission yeast Cut7+ gene. Nature 347, 563–566 (1990). 10. Hagan, I. & Yanagida, M. Kinesin-related Cut7 protein associates with mitotic and meiotic spindles in fission yeast. Nature 356, 74–76 (1992). 11. Straight, A. F., Sedat, J. W. & Murray, A. W. Time-lapse microscopy reveals unique roles for kinesins during anaphase in budding yeast. J. Cell. Biol. 143, 687–694 (1998). 12. Sawin, K. E., Leguellec, K., Phillipe, M. & Mitchison, T. J. Mitotic spindle organization by a plus-enddirected microtubule motor. Nature 359, 540–543 (1992). 13. Heck, M. M. et al. The kinesin-like protein KLP61F is essential for mitosis in Drosophila. J. Cell Biol. 123, 665–679 (1993). 14. Cole, D. G., Saxton, W. M., Sheehan, K. B. & Scholey, J. M. A ‘slow’, homotetrameric, plus-end directed kinesin purified from Drosophila embryos. J. Biol. Chem. 269, 22913–22916 (1994). 15. Kashina, A.S. et al. A bipolar kinesin. Nature 379, 270–272 (1996). 16. Kashina, A. S., Scholey, J. M., Leszyk, J. D. & Saxton, W. M. An essential bipolar mitotic motor. Nature 384, 225 (1996). 17. Sharp, D.J. et al. The bipolar kinesin, KLP61F, crosslinks microtubules within interpolar microtubule bundles of Drosophila embryonic mitotic spindles. J. Cell. Biol. 144, 125–138 (1999). 18. Blangy, A. et al. Phosphorylation by p34(Cdc2) regulates spindle association of human Eg5, a kinesinrelated motor essential for bipolar spindle formation in vivo. Cell 83, 1159–1169 (1995). 19. McDonald, H. B., Stewart, R. J. & Goldstein, L.S.B. The kinesin-like Ncd protein of Drosophila is a minus-end directed microtubule motor. Cell 63, 1159–1165 (1990). 20. Walker, R. A., Salmon, E. D. & Endow, S. A. The Drosophila claret segregation protein is a minus-end directed motor molecule. Nature 347, 780–782 (1990). 21. Matthies, H. J., McDonald, H. B., Goldstein, L. S. B. & Theurkauf, W. E. Anastral meiotic spindle morphogenesis: role of the Non-claret Disjunctional kinesin-like protein. J.Cell Biol. 134, 455–464 (1996). 22. Endow, S. A. & Komma, D. J. Centrosome and spindle function of the Drosophila Ncd microtubule motor visualized in live embryos using Ncd-GFP fusion proteins. J. Cell Sci. 109, 2429–2442 (1996). 23. O’Connell, M. J., Meluh, P. B., Rose, M. D. & Morris, M. R. Suppression of the bimC mitotic spindle defect by deletion of klpA, a kar3 related kinesin like protein in Aspergillus nidulans. J. Cell Biol. 120, 153–162 (1993). 24. Pidoux, A. L., LeDizet, M. & Cande, W. Z. Fission yeast pkl1 is a kinesin-related protein involved in mitotic spindle function. Mol. Biol. Cell. 7, 1639–1655 (1996). 25. Saunders, W., Lengyel, V. & Hoyt, M. A. Mitotic spindle function in Saccharomyces cerevisiae requires a balance between different types of kinesin-related motors. Mol. Biol. Cell. 8, 1025–1033 (1997). 26. Su, T. T. et al. Exit from mitosis in Drosophila syncytial embryos requires proteolysis and cyclin degradation and is associated with localized dephosphorylation. Genes Dev. 12, 1495–1503 (1998). 27. Lewis, E. B. & Gancarella, W. Claret and non-disjunction. Genetics 37, 600–601 (1952). 28. Vaisberg, E. A., Koonce, M. P. & McIntosh, J. R. Cytoplasmic dynein plays a role in mammalian mitotic spindle formation. J. Cell Biol. 123, 849–858 (1993). 29. Saunders, W. S., Koshland, D., Eshel, D., Gibbons, I. R. & Hoyt, M. A. Saccharomyces cerevisiae kinesin- and dynein-related proteins required for anaphase chromosome segregation. J. Cell Biol. 128, 617–624 (1995). 30. McIntosh, J. R., Hepler, P. K. & Van Wie, D. G. Model for mitosis. Nature 224, 659–663 (1969). 31. Francis-Lang, H., Minden, J., Sullivan, W. & Oegema, K. Live confocal analysis with fluorescently labeled proteins. Methods Mol. Biol. 122, 223–237 (1995). 32. Kellogg, D. R. et al. Studies on the centrosome and cytoplasmic organization in the early Drosophila embryo. Cold Spring Harb. Symp. Quant. Biol. 56, 649–662 (1991). ACKNOWLEDGEMENTS This work was supported by grants from the NIH (to J.M.S. and W.S.) and NIH postdoctoral fellowships (to D.J.S. and J.C.S.). We thank S. Hawley and members of his laboratory for help with this study and for providing the Cand fly stocks; R. Saint for providing the histone–GFP line; G. Rogers for Fig. 4 and his intellectual contribution to this work; and the other members of the Scholey and Sullivan laboratories for assistance. The anti-KLP61F antibody used here was made in collaboration with T. Mitchison and we thank him for his continuing interest and for discussions about mitosis. Correspondence and requests for materials should be addressed to J.M.S. © 1999 Macmillan Magazines Ltd NATURE CELL BIOLOGY | VOL 1 | MAY 1999 | cellbio.nature.com